
Santi Mohan Mandal
Central Research Facility, Indian Institute of Technology Kharagpur, Kharagpur-721302, WB, India
Dipjyoti Chakraborty
Department of Botany, University of Burdwan, Burdwan-713104, WB, India
Kajal Gupta
Central Research Facility, Indian Institute of Technology Kharagpur, Kharagpur-721302, WB, India
Research Journal of Seed Science
Year: 2008 | Volume: 1 | Issue: 1 | Page No.: 26-33







ABSTRACT
Variation in the seed size and seed mass in Hyptis suaveolens (L.) Poit. was studied and correlated with seedling establishment in a natural population. A two-fold increase in seed size and mass was observed in the large seeds over the small seeds. Large seeds showed a higher germination potential (78%) than smaller seeds (41%). α-amylase activity in large seeds was higher than small seeds. Endo-ß-mannanase expression in large seeds was detectable by Gel diffusion plate assay. Seedlings from large seeds were capable of emerging from greater burial depths. Total biomass and leaf area was greater in large seeded population in one-month seedling but subsequently after two months, the large and small seeded population showed no significant difference. Variation in seed size might be considered to be an adaptive response of H. suaveolens to its particular niche.
PDF Abstract XML References Citation
How to cite this article
Santi Mohan Mandal, Dipjyoti Chakraborty and Kajal Gupta, 2008. Seed Size Variation: Influence on Germination and Subsequent Seedling Performance in Hyptis suaveolens (Lamiaceae). Research Journal of Seed Science, 1: 26-33.
URL: https://scialert.net/abstract/?doi=rjss.2008.26.33
URL: https://scialert.net/abstract/?doi=rjss.2008.26.33
INTRODUCTION
There is an enormous variation in seed size throughout the range of vascular plants. Variation in seed size has been documented both among and within species (Michaeles et al., 1988; Westoby et al., 1992). Intraspecific seed size variability is regarded as an important aspect in the evolution of angiosperms (Venable, 1992). Variation in seed size can have a huge implication on germination (Wulff, 1986; Eriksson, 1999), seedling vigor (Turnbull et al., 1999), defoliation (Agren, 1989), drought (Caddic and Linder, 2002) and the competitive ability of the resulting plants (George and Bazzaz, 1999).
Seed size is a key determinant of the mobility, survival and success of plant propagules (Harper, 1997). Large seeds are generally suggested to be fit to survive environmental stress at the seedling establishment stage owing to larger food reserve (Leishman et al., 2000; Westoby et al., 2002). But, a ten-fold increase in seed mass is associated with a ten-fold decrease in the number of seeds that a plant can produce per unit canopy per year (Henery and Westoby, 2001), which gives the small seeds a better chance to disperse. This advantage of small seeded species due to seed size/number trade-off might be balanced by a combination of: (a) advantage of large seeded species during seedling establishment and (b) greater canopy area and longevity of large seeded species (Moles and Westoby, 2004).
Variations in seed size also lead to substantial changes in seed mass among different populations, within a population or even on the same plant. Seed mass is known have a positive correlation with both plant size and plant longevity (Rees, 1997; Ehrlen and Lehtila, 2002). The potential adaptive consequences of producing seeds of different mass have a variety of implications on germination
(Wulff, 1973), the ability to emerge from different burial depths (Ruiz de Clavizo, 2001), seedling vigor (Eriksson, 1999) and dispersal (Morse and Schmitt, 1985). Thus, both seed size and mass have important implications on the survivable and establishment of a species at its particular niche. The concept of seed mass as a straight trade-off between producing a large number of seeds, each with low establishment ability and producing fewer, larger seeds, each with a higher chance of successful establishment needs to be tested in species showing seed size/mass variation within the same plant.
Hyptis suaveolens (L.) Poit, an ethnobotanically important annual weed of Lamiaceae, is widely distributed in tropics and subtropics (Harley and Reynolds, 1992). It normally grows in places where soils have been profoundly disturbed and may be considered as rudral species (Wulff, 1973). The plant parts are used by tribals as stimulant, carminative, suderfic and galactogoguie (Kirtikar and Basu, 1991) and contain a high amount of volatile oils (Asekum and Ekundayo, 2002). The seeds of H. suaveolens show variation in size within the same population and even within the same plant (Wulff, 1973). Such seed size variation might have important consequences on seedling survivability of the species. The plant is aggressive and competitively dominant over other neighboring species, which might be attributed to its obnoxious nature or better survivability of seedlings.
The present study was undertaken to determine and consequences of seed size variation in H. suaveolens with regard to germination and seedling performance. We also studied the activity of α-amylase and endo-β-mannanase during germination. Both α-amylase and endo-β-mannanase is constitutively expressed in germinating seeds (Black, 1996; Choudhury and Gupta, 1998).
MATERIALS AND METHODS
Observations on growth, phenology, seed germination and dispersal were made in a natural population of H. suaveolens growing in the IIT- Kharagpur campus, WB, India. The main stem develops by late July. Stems erect, woody, much branched, growing to 1.5-2 m in height. The broad leaves are in opposite pairs on the stem with small mauve flowers in clusters in the upper leaf axils. Flowering starts from the month of September and the plants complete their life cycle by November-December. The seeds are enclosed within a persistent spiny calyx. Seeds were collected in the month of December and sun dried. They were stored in small glass vials in desiccators. The seeds were grouped into two classes according to their size as: Large seeds (3.76±0.12) and small seeds (2.89±0.04). Average seed weight (ten seeds, ten replicates) of the two size classes was taken using electron balance (Afcoset) and was 4.6±0.03 mg for the large seeds and 2.7±0.05 mg for the small seeds.
The following experiments were conducted with freshly harvested sun dried seeds. All treatments contained five replicates of 20 seeds in each set.
Germination Analysis
Effect of Light and Seed Size: Four sets each of large and small seeds were imbibed in water for 12 h. Imbibed seeds were taken in Petridishes on a double layer of Whatman No.1 filter paper moistened with 5 mL distilled water and allowed to germinate under continuous dark, 8, 12, 16 h photoperiod and continuous light at 30°C and 85-90% relative humidity inside the growth chamber (NK Biotron, Nippon Medical and Chemical Instruments Co. Tokyo, Japan).
Effect of temperature and seed size: Six sets each of large and small seeds were kept to germinate at 15, 20, 25, 30, 35 and 40°C under 16 h photoperiod and 85-90% relative humidity as above.
Daily germination counts were made of seeds with 2 mm of radicle protruding through the testa. Total germination percentage, time in h required for initiation of visible germination (T1) was considered for analysis. Mean Germination Time (MGT) and Coefficient of Velocity (CVG) of Germination was calculated using the method of Nicholas and Heydecker (1968).
MGT = Σ (nt)/ Σ (n), CVG = Σ (n)/ Σ (nt) x 100, where n = number of seeds emerged on a day, t = time or days from soaking.
To quantify the amount of mucilage secreted, seeds were immersed in water for 10 min and amount of mucilage formed calculated as a function of percentage increase in surface area.
α-Amylase and Endo-β-Mannanase Activity: Amylase activity was estimated according to the method of Choudhury and Gupta (1998). Briefly, imbibed seeds (one set each of large and small seeds) were allowed to germinate at 30°C, 16 h photoperiod and taken for analysis prior to initiation of germination. Seeds were homogenized in chilled 0.1 M sodium phosphate buffer (pH 6.8) at 0°C and amylase activity measured in terms of subsequent analysis of the residual substrate. For endo-β-mannanase activity, seeds were allowed to imbibe in distilled water and germinate described previously. Endosperm of each group of seed was homogenized with 200 μL of 0.1 M citric acid, 0.2 M sodium phosphate buffer (pH 5.0) for gel diffusion assay. Gel plates for the assay were prepared by dissolving 0.08% (w/v) galactomannan substrate (locust bean gum, sigma) in the same buffer. To dissolve the gum, the solution was heated at 95°C for 2 min and stirred vigorously for 30 min. It was then centrifuged at 11000 g for 15 min at 4°C. High melting agarose was added to the solution to a concentration of 0.8% (w/v) and heated to boiling. The solution was cooled to 50-60°C before 30 mL was dispensed into sterile petridishes. On solidification, wells were scoped out using cork borer. One hundred microliter of extract from each group was transferred to the wells of the gel-diffusion plates and allowed to diffuse 24 h at 25°C. Gels were stained with 10 mL of Congo red dye (Still et al., 1997).
Emergence: The ability of seedlings from large and small seeds to emerge from different burial depths was determined by burying germinating seeds (10 each of large and small) at 1, 2, 3, 4, 5 and 6 cm depth in soil trays. Three replicate trays per seed type and individual burial depth were used. Trays were examined over one month and the number of seedling emerged recorded.
Seedling Growth: To determine the growth rate of seedlings obtained from large and small seeds, 10 one-week-old seedlings of each type were transferred to pots. After 10 days, the length and width of cotyledons was measured and area calculated. After one and two month`s duration, the above ground vegetative portion of the plants was harvested and the number, length and width of all leaves recorded. Total dry weight of the plants was taken after oven drying 60°C for 48 h.
Data on germination and seedling parameters were collected and analyzed statistically. Analysis of variance (ANOVA) F-test was done and the critical difference were computed using Fishers LSD procedure at p = 0.05.
RESULTS
Germination
Effect of Light: The large seeds showed higher germination percentage irrespective of the photoperiod (Table 1). Optimum light requirement for germination was 16 h for both seed types. The larger seeds showed lower mean germination time and higher coefficient of velocity of germination. Continuous darkness was inhibitory in comparison to continuous light for both the large and small seeds.
| Table 1: | Effect of light on seed germination of H. suaveolens. (values represent mean±SD); T1 represent time requirement for the initiation of visible seed germination MGT = mean germination time, CVG = Coefficient of velocity of germination |
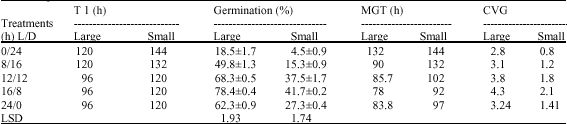 | |
| Table 2: | Effect of temperature on seed germination of H. suaveolens. (values represent mean±SD). T1 represents time requirement for the initiation of visible seed germination MGT = mean germination time, CVG = coefficient of velocity of germination |
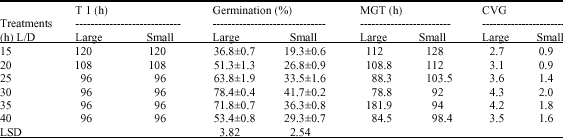 | |
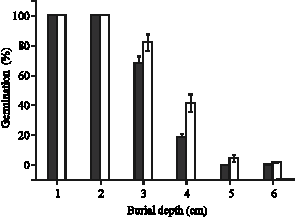 |
| Fig. 1: | Seedling emergence of large and small seeds of H. suaveolens buried at various depths |
Effect of Temperature: Optimum temperature for germination was found to be 30°C for both seed types (Table 2). The MGT and CVG decreased below and above this temperature in both seed types. Although the germination percentage did not decrease sharply in the large seeds as observed in the small seeds on increasing the temperature from 30 to 35°C, the mean germination time was higher for the large seeds.
Amylase activity was observed in germinating seeds of both sizes. The activity was higher in large seeds (0.229±0.003 mg substrate hydrolyzed g-1 d.wt min-1) than the small seeds (0.137±0.002 mg substrate hydrolyzed g-1 d.wt min-1). Endo-β-mannanase activity could not be detected in germinating small seeds by gel diffusion assay whereas large seeds showed high endo-β-mannanase expression as observed by the clear hydrolysis zone in galactomannan agar plate.
Emergence: Seedlings from large seeds emerged from greater burial depths than that of small seeds (Fig. 1). Seedlings of both large and small seeds at depths of 1-2 cm emerged successfully but at
| Table 3: | Characteristics of the above ground seedling of one and two-month-old H. suaveolens plants from large and small seeds (values represent mean±SD) |
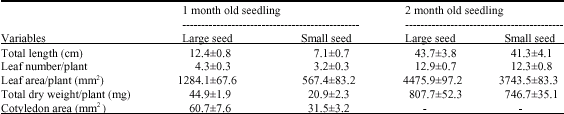 | |
greater depths, the percentage emergence was significantly higher for large seed seedlings. Emergence was impaired at depths more than 4 cm.
Seedling Growth: The area of cotyledons of 10-day-old seedlings from large seeds was significantly greater than that of seedling from small seeds (Table 3). One-month-old seedling exhibited significant difference in leaf area, plant dry weight and height of the plant. This difference was subsequently decreased over the next one month.
DISCUSSION
Generally, seed size differences within a species is induced by environmental factors such as restrictions in resource availability as the growing season develops (Vaughton and Ramsey, 1998) and by changes in other ecological factors that affect plant growth (Pitelka et al., 1983). The first question to ask is whether there is any reason to seek a general adaptive explanation for seed size variation with this magnitude. According to Winn (1991), plants may not have the capability of producing a completely uniform seed size, simply as a result of variations in resource availability during fruit development. Present results indicate that seed size-mass variation in H. suaveolens may be a part of its survival strategy in a rudral habitat.
When a seed has reached its microclimate it must germinate and establish or lie dormant until favorable conditions are available. Light quality, temperature and moisture are the most important eco-physiological factors regulating the germination of seeds. H. suaveolens seeds on wetting produce large amount of mucilage effectively increasing seed size by about 138% in both large and small seeds. Myxocarpy-the condition of having mucilaginous cells in the endocarp has been reported in a large number of Labiates, which might help in water retention and survivability of the germinating seeds in wild. A light requirement for germination is one of the main determinants of the ability of species to accumulate a persistent soil seed bank (Pons, 1991). It can determine the position in or on the soil where germination occurs. Germination in a number of species is initiated when the hydrated seed is illuminated with white light (Toole, 1973). The seeds of H. suaveolens were found to be positively photoblastic and germination percentage increased with increasing duration of light period with a slight inhibition at continuous light (Table 1). Mean germination time was lowest and coefficient of velocity of germination highest for large seeds at 16 h photoperiod and it might be optimum for germination though such a light period is generally unavailable to plants in the wild. Under natural conditions, the photoperiod remains mostly around 12 h where about 68% germination was observed in the large seeds. Earlier reports show a higher germination percentage of large seeds of H. suaveolens under light saturation period (Wulff, 1973). The abundance of H. suaveolens in disturbed soils may be explained by the present observations with photoperiod, that as the seeds are suddenly exposed to more light in upturned soil they will show a flush of germination. A light requirement hinders germination of seeds that are buried too deep in the soil, which if germinated would have produced non-viable seedlings. Milberg et al. (2002), have shown that large seeds have a lower light requirement than a smaller seed for germination and it was evident in H. suaveolens, which shows a variation in seed size.
The effect of light on seed germination is frequently dependent upon temperature since phytochrome undergoes some thermal reactions such as thermal reversion and possible destruction (Bewley and Black, 1994). The optimum temperature requirement was 30°C for both large and small seeds. A higher temperature regime, though, decreased germination was not inhibitory suggesting that under natural conditions, H. suaveolens seeds are more likely to germinate in the hot and humid monsoons rather than the dry, cold and winter season thus ensuring better plantlet establishment.
Reserve effect is one of the major strategies adopted by plants for ensuring seedling survival and competitive edge to the emerged seedling (Leishman et al., 2000). The larger a seed is the greater will be the metabolic reserve available to make up for lack of resources in the environment. A larger seed size also confers enhanced ability to penetrate ground cover (Rees, 1997). During germination, hydrolytic enzymes are activated that metabolize the storage protein for utilization by the growing embryo. The amino acids obtained from the hydrolysis of storage protein are required for the translation of amylase that catalyzes the hydrolysis of starch (Teiz and Zeiger, 2002). Amylase activity as a response to changes in phytochrome during development and germination is reported in cereal grains (Sinjorogo et al., 1993; Bewly and Black, 1994). In the present experiments, the higher amylase expression during germination of large seeds might be due to the high amount of storage protein and carbohydrate. Large seeds produce more sugar monomer due to high-level amylase, which catalyzes the hydrolysis of starch into sugar. This sugar then fuels respiration in the embryo facilitating early germination.
Tissues surrounding the embryo form a barrier that needs to be penetrated for radicle emergence after embryo germination. The mechanical resistance to penetration of this endosperm cap weakness during imbibitions as the time of radicle emergence approaches (Dahal and Bradford, 1990) and there is a linear relationship between endosperm puncture force and germination rate (Karssen et al., 1989). The cell walls of the micropylar endosperm cap are composed predominantly of some galactomannan polymer such as mannose, galactose that is cleaved by endo-β-mannanase (Groof et al., 1988). Our observations show high endo-β-mannanase activity in large seeds prior to initiation of germination. Thus, cell wall weakening of the micropylar endosperm cap due to the hydrolysis of galactomannan substrate favors early radicle emergence in large seeds.
With regards to emergence from soil, germinating seedlings of H. suaveolens from large seeds were more adapted to emerge from greater burial depths than small seeds. A large seed size often confers enhanced ability to penetrate ground cover and survive burial by litter (Metcalfe and Grubb, 1997). Seedling from large seeds initially showed vigorous growth and possess larger cotyledons than those of small seeds. The differences in growth pattern of seedlings derived from large seeds and those from small seeds were prominent during the first one-month after which the variation was not significant. Thus, it might be suggested that after reaching a certain phase, the growth pattern becomes constant and to attain that stage in seedling development, plantlets from small seeds have to develop more rapidly. Survival was quantified over a time likely to have encompassed most of the period over which large seediness is expected to benefit seedlings. Our results correlate with studies where seedling establishment was monitored in later stage seedlings, the early advantage if any of the large seeded species disappeared with time (Saverimuttu and Black, 1996). Furthermore, a study quantifying the relationship between seed mass and seed production per unit canopy has shown that relationships with slopes are not significantly different (Henery and Westoby, 2001).
CONCLUSIONS
Seed traits are embedded in the life cycle of plants and its strategy must match the strategy of the established plant. H. suaveolens plants dominate in a rudral habitat (high nutrition, disturbed habitat). Seed dispersal, germination, seedling emergence and subsequent survival of the species dictate its competitiveness and success in such a habitat. Variation in seed size in H. suaveolens might be a strategy for increasing the probability of success in its particular niche. A plant that has evolved to live in a particular niche must have seeds that can germinate, disperse and establish in there (Rees, 1997). The presence in H. suaveolens of seeds of two size classes seems to be a major contributor to the success of the plant in a rudral habitat. Large seeds are advantageous during rigorous and unfavorable environmental conditions seed germination and seedling emergence is ensured. At favorable environmental conditions the numerical advantage of small seeds gives it a competitive edge over other species. Instead of producing only large seeds thus compromising on number or only small seeds which while ensuring greater dispersal thereby depending heavily on ambient environmental conditions for seedling emergence, seeds of both size classes are produced in H. suaveolens plants which benefits the species under variable environmental conditions. It also ensures optimum utilization of the available resources which might otherwise have gone waste in increasing seed size or seed number.
ACKNOWLEDGMENT
The authors are indebted to the SAP, UGC for financial assistance and The University of Burdwan for providing necessary facilities.
REFERENCES
- Agren, J., 1989. Seed size and number in Rubus chamaemorus: Between habitat variation and effects of defoliation and supplemental pollination. J. Ecol., 77: 1080-1092.
Direct Link - Asekum, O.T. and Ekundayo, 2002. Essential oil constituents of Hyptis suaveolens (L.) Poit, (bush tea) leaves from Nigeria. J. Essen. Oil Res., 12: 227-230.
Direct Link - Black, M., 1996. Liberating the radicle: A case for softening-up. Seed Sci. Res., 6: 39-42.
CrossRefDirect Link - Caddic, L.R. and H.P. Linder, 2002. Evolutionary strategies for reproduction and dispersal in African Restionaceae. Aust. J. Bot., 50: 339-355.
Direct Link - Choudhury, S. and K. Gupta, 1998. Studies on the germination mechanism of Catharenthus roseus (L.) G. Don cv alba seeds: Effect of promoters and pH. Seed Sci. Technol., 26: 719-732.
Direct Link - Dahal, P. and K.J. Bradford, 1990. Effects of priming and endosperm integrity on seed germination rates of tomato genotypes: II. Germination at reduced water potential. J. Exp. Bot., 41: 1441-1453.
CrossRef - Eriksson, O., 1999. Seed size variation and its effect on germination and seedling performance in the clonal herb Convallaria majalis. Acta Oecol., 20: 61-66.
CrossRefDirect Link - George, L.O. and F.A. Bazzaz, 1999. The fern understory as an ecological filter: Emergence and establishment of canopy tree seedling. Ecology, 80: 833-845.
CrossRef - Groof, S.P.C., B. Kielszewska-Rokicka, E. Vermeer and C.M. Karssen, 1998. Gibberellin induced hydrolysis of endosperm cell wall in gibberellin-deficient tomato seeds prior to radicle protrusion. Planta, 174: 500-504.
CrossRef - Henery, M.L. and M. Westoby, 2001. Seed mass and seed nutrient content as predictors of seed output variation between species. Oikos, 92: 479-490.
Direct Link - Michaels, H.J., B. Benner, A.P. Hartgerink, T.D. Lee, S. Rice, M.F. Willson and R.I. Bertin, 1988. Seed size variation: Magnitude, distribution and ecological correlates. Evol. Ecol., 2: 157-166.
CrossRefDirect Link - Milberg, P., L. Andersson and K. Thompson, 2000. Large-seeded spices are less dependent on light for germination than small-seeded ones. Seed Sci. Res., 10: 99-104.
Direct Link - Moles, A.T. and M. Westoby, 2004. Seedling survival and seed size: A synthesis of the literature. J. Ecol., 92: 372-383.
Direct Link - Nichols, M.S. and W. Heydecker, 1968. Two approaches to the study of germination data. Proc. Int. Test Assoc., 33: 531-540.
Direct Link - Pitelka, L.F., M.E. Thayer and S.B. Hansen, 1983. Variation in achene weight in Aster acuminatus. Can. J. Bot., 61: 1415-1420.
CrossRefDirect Link - Ruiz-de-Clavizo, E., 2001. The role of dimorphic achenes in biology of the annual weed Leonodon longirrostris. Weed Res., 41: 275-286.
Direct Link - Sinjorgo, K.M., M.A. de Vries, J.C. Heistek, M.J. van Zeijl, S.W. van der Veen and A.C. Douma, 1993. The effect of external pH on the gibberellic acid response to barley aleurone. J. Plant Physiol., 142: 506-509.
CrossRefDirect Link - Still, D.W., P. Dahal and K.J. Bradford, 1997. A single seed assay for endo-mannanase activity from tomato endosperm and radicle tissues. Plant Physiol., 13: 13-20.
Direct Link - Teiz, L. and E. Zeiger, 2002. Plant Physiology. 3rd Edn., Sinauer Associates, Sunderland, Massachusetts, ISBN: 9780878938230, Pages: 690.
Direct Link - Turnbull, L.A., M. Rees and M.J. Crawley, 1999. Seed mass and the competition/colonization trade-off: A sowing experiment. J. Ecol., 87: 899-912.
CrossRefDirect Link - Vaughton, G. and M. Ramsey, 1998. Sources and Consequences of seed mass variation in Banksia marginata (Proteaceae). J. Ecol., 86: 563-573.
CrossRefDirect Link - Westoby, M., E. Jurado and M. Leishman, 1992. Comparative evolutionary ecology of seed size. Tree Ecol. Evol., 7: 368-372.
CrossRefDirect Link - Westoby, M., D.S. Falster, A.T. Moles, P.A. Vesk and I.J. Wright, 2002. Plant ecological stratigies: Some leading dimensions of variation between species. Annl. Rev. Ecol. Syst., 33: 125-129.
Direct Link - Wulff, R., 1973. Intrapopulational variation in the germination of seeds in Hyptis suaveolens. Ecology, 54: 646-649.
CrossRefDirect Link - Wulff, R., 1986. Seed size variation in Desmodium paniculatum: II. Effects on seedling growth and physiological performance. J. Ecol., 74: 99-114.
Direct Link