
Pramod Kumar Singh
Babu Banarasi Das Northern India Institute of Technology, Lucknow-226028, India
Praveen Kumar
Department of Botany, University of Lucknow, Lucknow-226007, India
Pramod Kumar Tandon
Department of Botany, University of Lucknow, Lucknow-226007, India
Research Journal of Soil Biology
Year: 2014 | Volume: 6 | Issue: 1 | Page No.: 1-16







ABSTRACT
Soil sodicity is worldwide problem in arid and semi-arid region. In India large areas of sodic soils exist. Screening of commercial crops for their tolerance towards sodicity can encourage farmers to grow them on these lands thereby rehabilitating them. Fennel (Foeniculum vulgare) is a cash crop and its aromatic fruit is used as condiment and also in traditional and modern medicine. A soil pot culture study was conducted to assess salt tolerance status of fennel in sodic soils. The plants were raised on soil having different levels [10, 20, 25, 35 and 40 ESP (exchangeable sodium percentage)] of soil sodicity in complete randomized design. Various parameters such as plant growth, biomass, activities of anti-oxidative enzymes, water content, proline accumulation, concentration of photosynthetic pigments, cation concentration and essential oil quality were studied. Results indicated that increasing soil sodicity resulted in significant decreases in plant growth, biomass and leaf water potential (Ψ became more negative). However, cell sap pH, electrical conductivity, sodium concentration and proline accumulation were increased. Increasing soil sodicity increased activities of antioxidant enzymes like catalase and superoxide dismutase while peroxidase and nitrate reductase declined. The chlorophylls and carotenoid concentration decreased at higher sodicity while carotenoids/chlorophyll ratio increased. Seed quality improved at higher ESP. The gas chromatogram for essential oil revealed that improved seed quality resulted from increased trans-Anethole (sweetness of seed) and decreased d-fenchone (bitterness of seed) content. Thus, fennel crops can be recommended for growth in sodic soils (ESP up to 25) with economic gains.
PDF Abstract XML References Citation
Received: January 10, 2013;
Accepted: March 12, 2013;
Published: April 15, 2014
How to cite this article
Pramod Kumar Singh, Praveen Kumar and Pramod Kumar Tandon, 2014. Soil Sodicity Alters Antioxidative Enzymes, Photosynthetic Pigments, Water
Content and Essential Oil Quality of Fennel (Foeniculum vulgare Mill.). Research Journal of Soil Biology, 6: 1-16.
DOI: 10.3923/rjsb.2014.1.16
URL: https://scialert.net/abstract/?doi=rjsb.2014.1.16
DOI: 10.3923/rjsb.2014.1.16
URL: https://scialert.net/abstract/?doi=rjsb.2014.1.16
INTRODUCTION
Fennel is biennial herb commonly found in Mediterranean region. This is horticultural crop grown for production of seeds used as spice or the essential oil for industrial application in perfumery, cosmetic, pharmaceutics and vegetative parts used in salad. The constituents of essential oil, including trans-anethole, d-fenchone, camphene and methylchavicol are paramount importance in the pharmaceutical industries and in confectionary (Wealth of India, 1978; Abdallah et al., 1998). In India, fennel is cultivated in arid and semiarid region, mainly in the state of Rajasthan, Gujarat and Uttar Pradesh where about one third of the salt affected soils of India occur (Yadav, 1993). The utilization of salt affected wastelands can be promoted in view of the limited land resources by growing spice crops. In India, however, there are large tracts of salt affected soils (7.4 million ha) of which about 50.0% is adversely affected by sodicity (Tyagi and Minhas, 1998). Recently, Garg (2011) reported that fennel has potential of growing in sodic soil without affecting its oil content. In sodic soil, soil reaction and salt (particularly Na) content are the two factors which are of prime importance in evaluating the production potential of most of the crops (Bernstein, 1975). Excess Na may compete with K in membrane transport and when accumulated in the cytoplasm, it inhibits many enzymes (Epstein, 1998). More than 50 enzymes are activated by K+ and Na+ cannot substitute in this role (Bhandal and Malik, 1988). Increased intracellular Na concentration is also believed to predispose plants to oxidative stress. The high level of antioxidant enzyme activities is involved in salt tolerance and repair of oxidative damage resulting from salt stress (Fahmey et al., 1998). The cells are protected against Reactive Oxygen Species (ROS) such as •O2 (super oxide radicals), •OH (hydroxyl radical) and H2O2 (hydrogen peroxide) by operation of intricate anti-oxidative mechanism (Foyer et al., 1997). The enzymatic processes basically involve dismutation of •O2 by superoxide dismutase that generates another partially reduced oxygen species, H2O2. Normally the enzymes– catalase and peroxidase, catalyze the removal of cellular H2O2 (Alscher et al., 1997).
In past years, considerable work has been done overseas with cereals, oil seed and vegetable crops to assess the tolerance of these crops in sodic soil (Mass and Hoffmann, 1977; Abrol and Bhumbla, 1979; Singh et al., 1981; Singh and Abrol, 1985; Garg and Srivastava, 1986). There is inadequate published information available about fennel tolerance and antioxidative response against sodic soils (Singh et al., 2002; Garg et al., 2004, 2005; Tandon et al., 2009; Garg, 2012). An alternate approach is to utilize sodic soils for growing compatible crop species and aromatic crops and producing economic yield. Hence the present study was undertaken with the objective to assess the tolerance status, metabolic activities and essential oil quality of fennel influenced by soil sodicity.
MATERIALS AND METHODS
Experimental site: The pot experiment was conducted for two consecutive years at Banthra Research Station of National Botanical Research Institute, Lucknow. The area lies between 26°40'-26°45' N latitude and 80°45'-80°53' E on the Lucknow-Kanpur highway at an elevation of 129 m above the Mean Sea Level (MSL). The meteorological parameters indicate that the climate of the area is semi-arid, subtropical and monsoonic with an average annual rainfall of 872 mm. The mean maximum and minimum temperatures were 39.1°C and 7.6°C, respectively (Garg et al., 2000).
Plant material and experimentation: Fennel (Foeniculum vulgare Mill.) were grown at five different soil sodicity (ESP) levels i.e., 10, 20, 25, 35 and 40 in Complete Randomized Design (CRD) and each with four replications. The soils were collected from the different five sites of the farm upto 15 cm depth and kept it for drying. The physicochemical properties of soils were analysed adopting the methods of (Richards, 1954) and are indicated in Table 1. After drying, the soils were thoroughly crushed, properly mixed with a basal dose of 200 mg N kg-1 soil as Ca (NO3)2, 100 mg P kg-1 soils as KH2PO4 and 100 mg S kg-1 soil as MgSO4 and filled in earthen clay pots. Prior to filling of soils, these pots were lined on their inner side by alkathene sheet to check the leaching of ions and also contamination from the clay of the pots. Fifteen seeds of fennel were sown in the last week of October and thinning was done after 30 days to allow 4 plants to grow in each pot.
| Table 1: | Physicochemical properties of the soils collected from different sites for soil pot culture |
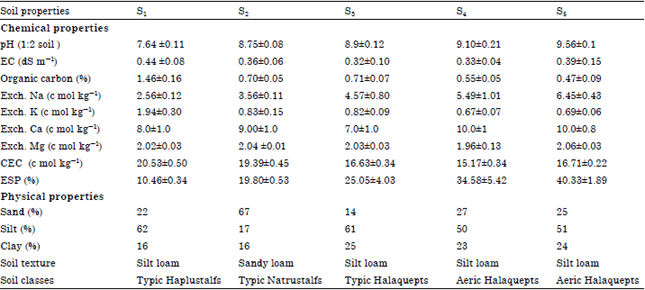 | |
| *S indicates levels of soil sodicity S1 = 10 ESP, S2 = 20 ESP, S3 = 25 ESP, S4 = 35 ESP and S5 = 40 ESP: Exchange able sodium percentage, EC: Electrical conductivity | |
The ESP level was maintained throughout experiment by supplying carbonates and by carbonate mixture solution and irrigation was done when required by tap water. The crop was harvested in second week of April.
Growth, Cell sap pH, EC, water potential and water status: Data were recorded as plant height and dry matter yield (oven dried at 70°C for 24 h) and seed yield. Plant height was measured from soil level to the top of umbel. The water status of leaf tissue was ascertained when plants were about 100 days old. At the same time cell sap pH and Electrical Conductivity (EC) of leaf samples were determined using EC and pH meter after homogenization one gram of fresh leaves in 20 mL of 0.025 M EDTA solution (Dwivedi et al., 1980). Leaf water potential (Ψ) was measured using Wescor’s microvoltmeter model HR33T and C-52 leaf chambers when the plant growing at field capacity. The same leaves were sampled for measuring water status of the leaf. Determination of Water Saturation Deficit (WSD) was made by measuring fresh and hydrated (incubated in glass distilled water) for three hours at 10°C in the dark) and oven dried weights. Leaf area was determined by Leaf Area meter Delta-T devices. The WSD and Specific Water Content (SWC) were calculated following standard procedure (Barr and Weatherley, 1962).
Yield and cation analysis: Yield was recorded at harvest. Harvested plants were washed and thoroughly separated into root, stem and leaves and dried in an oven at 70°C for 24 h. Tissue concentration of cations were measured in the solution after wet digestion (HNO3: HClO4, 10:1v/v mixture) of the oven dried plant material (Piper, 1942). The cations (Na, K and Ca) were measured on flame photometer while Mg was measured on atomic absorption spectrophotometer. The cation accumulation was calculated by multiplying of dry weight with cation concentration.
Chloroplastic pigments, proline and soluble protein: Concentration of chlorophylls and carotenoids were determined in 80% acetone extract of the young fully expanded fourth leaf by the method of Lichtenthaler (1987). The homogenate was centrifuged at 4000x g for 10 min to remove the residue. The colour intensity of clear supernatants was measured at 663.2, 646.8 and 470 nm for chlorophyll a, chlorophyll b and carotenoids, respectively. Results have been expressed as mg chlorophyll or carotenoids g-1 fresh weight. The concentration of free proline was determined in fresh leaf tissue with acid ninhydrin complex in toluene (Bates et al., 1973). The protein concentration in the homogenate was determined in Tri Chloro Acetic acid (TCA) precipitate according to Lowry et al. (1951) using Bovine Serum Albumin (BSA) as standard.
Enzyme extraction: Fresh leaf tissue (2.5 g) was homogenized in 10.0 mL of chilled 50 mM potassium phosphate buffer (pH 7.0) containing 1.0% insoluble Polyvinyl Pyrrolidone (PVP) using chilled pestle and mortar kept in ice bath. The homogenate was filtered with two-fold muslin cloth and centrifuged at 20,000x g for 10 min in refrigerated centrifuge at 2°C. The supernatant was stored at 2°C and used for enzyme assays within 4 h.
Enzyme assays: The activity of catalase (EC. 1.11.1.6) was assayed using the method of Euler and Josephson (1927) as modified by Bisht et al. (1989). Reaction mixture (10 mL standardized against 0.1N KMnO4) containing 0.5 mM H2O2 and 1.0 mM of potassium phosphate buffer (pH 7.0) was taken in a test tube and stabilized at 25°C. The reaction was initiated by adding 1 mL of suitably diluted enzyme extract and the contents were mixed thoroughly. The reaction was allowed to proceed for five minutes and then stopped by addition of 2.0 mL of 4N H2SO4. Corresponding blanks, in which H2SO4 was added prior to the addition of enzyme extracts, were run simultaneously. The final reaction mixture was then titrated against 0.1N KMnO4. The H2O2 decomposed was then calculated as the difference in titer value of respective blank and sample. Enzyme activity is expressed as μ mole H2O2 reduced per unit fresh matter or protein weight.
Peroxidase (EC 1.11.1.7) was assayed after Bisht et al. (1989), a modified method of Luck (1963). The reaction mixture (5.0 mL) contained 2.0 mL of 0.1 M potassium phosphate buffer (pH 7.0), 1.0 mL of 0.01% H2O2 and 1.0 mL of 0.05% p-phenylenediamine. The reaction was started by adding 1.0 mL of diluted enzyme extract to the reaction mixture and allowed to proceed for 5.0 min. Reaction was stopped by adding 2.0 mL of 4 N H2SO4. Corresponding blanks were maintained in which H2SO4 was added to the substrate mixture prior to the addition of enzyme extract. These tubes were kept in refrigerator for 30 min at 4°C and then contents were centrifuged at 4000x g for 15 min at room temperature. The color intensity of the supernatant was read at 485 nm. The enzyme activity was expressed in units 100 per mg Fresh Weight (FW) or per mg protein, an enzyme unit being defined as the difference of 0.01 in the optical density between blank and sample per minute of reaction time.
The activity of Superoxide dismutase (EC 1.15.1.1) was assayed by measuring its ability to inhibit the photochemical reduction of Nitro-Blue Tetrazolium (NBT) adopting the method of (Beauchamp and Fridovich, 1971). The 3.0 mL reaction mixture contained 50.0 mM potassium phosphate buffer (pH 7.8), 10 mM methionine, 1.17 mM riboflavin and 56 mM NBT and enzyme extract. Riboflavin was added in last and switching on the light started reaction. The reaction was allowed to take place for 30 min and was stopped by switching of the light. The absorbance of the solution was measured at 560 nm and from which the absorbance of the unirradiated reaction mixture that served as respective blank was deducted. A 560 was plotted as a function of fresh matter equivalent of the reaction mixture. From the resultant graph, fresh matter equivalents of enzyme extract corresponding to 50% inhibition of the reaction was read and considered as one enzyme unit.
Nitrate reductase (1.6.6.1) was assayed by the method of Hageman and Reed (1980) in vivo. Chopped leaf tissue (500 mg) was placed into the test tube. Incubation medium comprising of 0.1 M (molar) KH2PO4 pH 7.5, 1% v/v 1-propanol and 0.03 to 0.05 M KNO3 incubated for 30 min at 30°C on water bath and stopped the reaction by placing tubes in boiling water for 2 min. Aliquots were taken and NO2 was estimated. The difference between NO2 formed in 30 min. and NO2 produced in 5 min was used to calculate the rate at NO2 production (equated with NO3 reduction). Standard curve was prepared by KNO2, 1.0 mL of sulphanilamide (1% in 1.5 N HCl), 1.0 mL of 0.02% NED [N-(1-Napthyl) ethylene diamine hydrochloride] in Dry Weight (DW). These were mixed well and allowed to stand 20 min at room temperature and Optical Density (OD) was read at 540 nm.
Essential oil contents and chemical constituents: The essential oil content was determined by hydro-distillation (Langenau, 1948) of the powdered seeds in Clevenger type apparatus (Clevenger, 1928) for 4 h. The major chemical constituents of the oil were determined by gas liquid chromatography which was done with multiple temperature programming and FID detector having capillary column 30x0.25 mm. Oven temperature was programmed from 80-230°C with a rise of 5°C min-1. Injector and detector temperature was maintained at 200 and 230°C, respectively.
![]()
Statistical analysis: Statistical method and test of significance appropriate to the design were applied to the data for discriminating the treatment effects from chance effects. To elucidate the nature and magnitude of the effect, the level of significance was calculated by one way ANOVA and 'F' test in the analysis of variance (Panse and Sukhatme, 1967).
RESULTS
Plant growth and yield: Increase in soil ESP markedly suppressed plant growth, the suppression in growth was more pronounced at 35 and 40 ESP that was 80 and 74% of control, respectively (Table 2). The decrease in the total biomass yield was very steep till 35 ESP and 40 ESP which were decreased 57.38 and 62.2%, respectively. The biomass yield was obtained 55.4% at 25 ESP.
Cell sap pH, EC, water relation and proline content: The cell sap pH in leaf tissue was non-significantly increased from 4.1 in control to 4.4 at high ESP while EC was significantly increased on increasing the soil ESP. The increasing ESP levels resulted in significant decreased in water potential (from -9.44 Ψ to -10.52 Ψ, became more negative), SWC and degree of succulence were decreased on increasing ESP while WSD increase. The accumulation of proline was significantly increased on increasing the soil ESP. In control proline content was 16.8 mg g-1 FW and at 40 ESP level it was 31.5 mg g-1 FW which was increases 78.4% than the control (Table 3).
Chloroplastic pigments: Chlorophyll and carotenoids content decreased with increasing ESP. The decline was steep for first 15 units increase in ESP i.e., from 10-25 ESP (52% for total chlorophyll and 42% for carotenoids) than noticed from 25-40 ESP (31% for chlorophyll and 21% for carotenoids) (Table 4).
| Table 2: | Plant growth, biomass and essential oil contents of fennel plant grown at different soil sodicity levels |
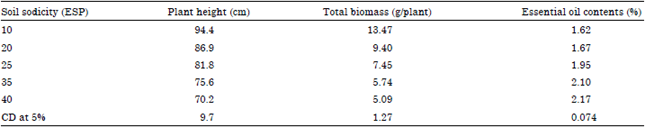 | |
| Table 3: | Soil sodicity induced changes in cell sap pH, cell sap EC, water relation and proline content in fennel plant |
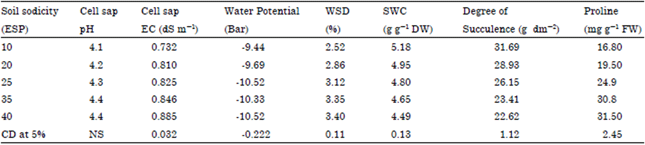 | |
| WSD: water saturation deficit, SWC: Specific water content, FW: Fresh weight | |
| Table 4: | Soil sodicity induced changes in the chlorophyll contents (a, b and total chlorophyll) and carotenoids in the leaf of fennel |
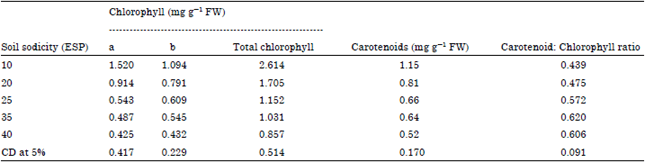 | |
Similar trend was obtained for chlorophyll a, but decline in chlorophyll b was gradual. Carotenoids/chlorophyll ratio was significantly increased with increasing soil ESP (from 0.44 in control to 0.61 at 40 ESP).
Activities of oxidative stress-responsive enzymes: Variable effect of increasing ESP on activities of catalase, peroxidase and SOD were observed. Significant increases in the activities of catalase and SOD were noticed with increasing ESP, however, the magnitude of increase was more pronounced beyond 20 ESP (activity of catalase was nearly 183% of control at 40 ESP and activity of SOD was 200% of control at 40 ESP). On the contrary a reverse trend was obtained for peroxidase (activity was about 86% of control at 40 ESP), activity decreased gradually in response to increase in soil ESP (Table 5).
Nitrate reductase activity and soluble protein: The activity of nitrate reductase decreased on increasing soil ESP. However, the decreases were significant above 25 ESP (activity was about 78% of control at 40 ESP). The concentration of soluble protein decreased progressively, in the leaf tissue with increasing soil ESP and was nearly 81% of control at 40 ESP (Table 5).
| Table 5: | Soil sodicity alters in antioxidative enzyme (CAT, POD and SOD), nitrate reductase and soluble protein in the leaf of fennel |
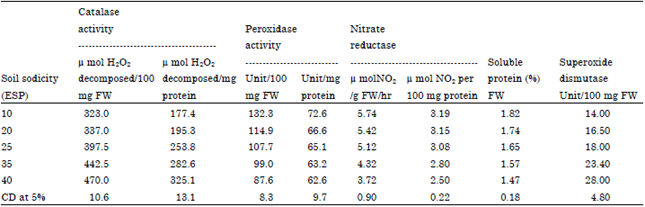 | |
| CAT: Catalase, POD: Peroxidase, SOD: Superoxide dismutase, Hr: Hour | |
| Table 6: | Effect of soil sodicity on cation concentration in different plant parts of fennel |
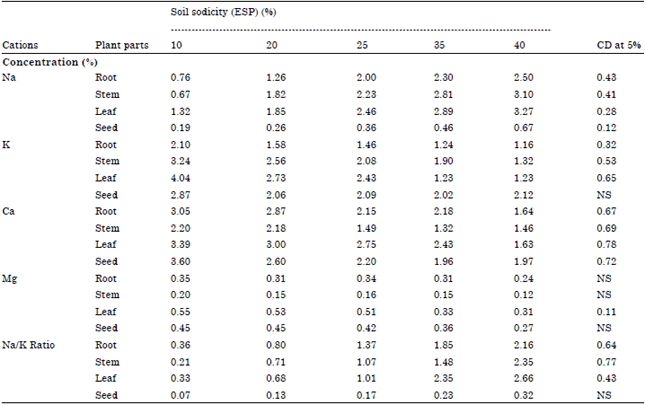 | |
| Na: sodium, K: Potassium, Ca: Calcium: Mg: Magnesium | |
Cation concentration: The concentration of Na was significantly increased in all plant parts i.e. root, stem, leaf and seed on increasing soil ESP while K, Ca and Mg were decreased (Table 6). Maximum concentration of Na was noticed in leaves and least in seed. The Na/K ratio was also increased on increasing soil ESP (Table 6). However, significant increase in Na/K ratio was noticed above 25 ESP.
Essential oil contents and major chemical constituents: The essential oil content increased on increasing the soil ESP. The essential oil content was 1.62% in control and maximum 2.17% at 40 ESP (Table 2).
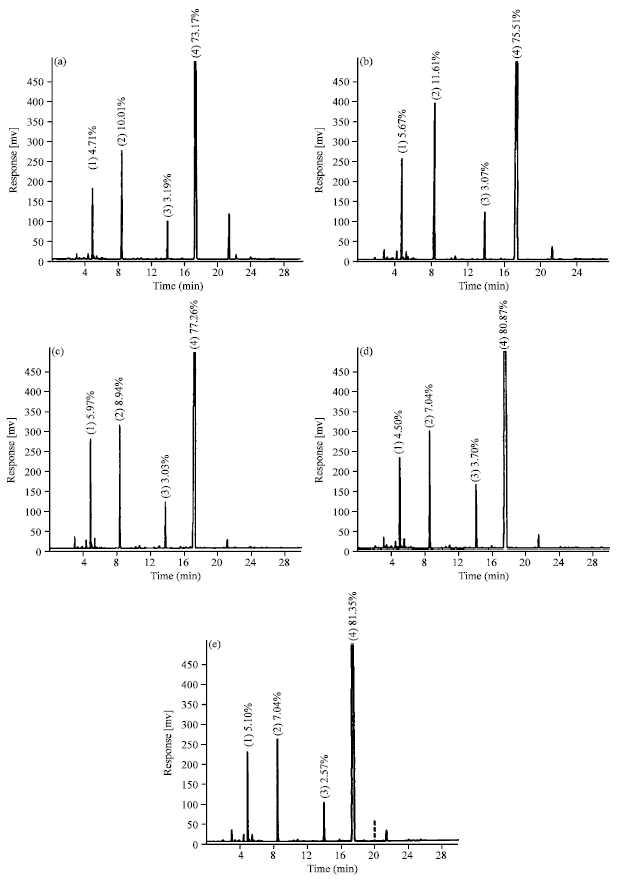 | |
| Fig. 1: | Gas chromatograph indicates major chemical constituent’s content in essential oil of fennel grown at different soil sodicity (exchangeable sodium percentage). a: Control (10 ESP), b: 20 ESP, c: 25 ESP, d: 35 ESP, e: 40 ESP with 1st peak: Limonene, 2nd: d-fenchone, 3rd: Methylchavicol and 4th: trans-anethole |
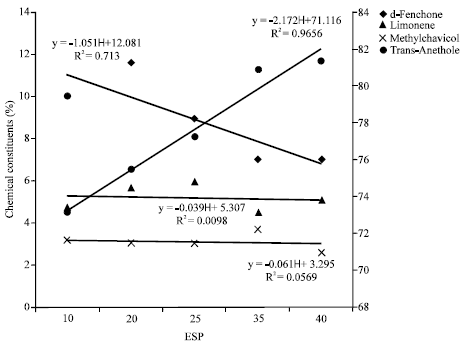 | |
| Fig. 2: | Correlation between different chemical constituents of fennel oil and soil sodicity (exchangeable sodium percentage) |
The increase in essential oil content was steep for last 15 units increase in ESP i.e., from 25-40 ESP(20, 30 and 34% increase than the control at 25 ESP, 35 ESP and 40 ESP, respectively). The Gas Liquid Chromatography analysis of essential oil indicated that the trans-anethole was the major constituent of essential oil of fennel and was found to increase with increasing the soil ESP (Fig. 1). It was 73% at 10 ESP and 81.35% at 40 ESP. While other major constituents like d-fenchone decreased on increasing the soil ESP. The d-fenchone varied from 10.01% at 10 ESP to 7.04% at 40 ESP. An inconsistent variation was observed in the case of limonene and methylchavicol contents. Correlation analysis revealed a positive relationship among trans-anethole and d-fenchone contents (R2 = 0.965 and R2 = 0.713, respectively) with soil ESP and displayed a reciprocal relationship between each other (Fig. 2).
DISCUSSION
Increasing ESP profoundly affected the survival, growth and metabolic process of fennel. Plant height was greatly suppressed and stunting, was apparent at 25 ESP. Similar retardation of growth in fennel due to salinity was observed by Mangal et al. (1986) and Graifenberg et al. (1996). The reduced growth was likely caused by the high Na concentration that lead to increasing osmotic potential of the circulating soils solution as well as due to ionic imbalance resulting from excessive uptake of Na ions. The plant require more energy making osmotic adjustments by accumulating organic and inorganic solutes to lower the osmotic potential inside their cells to counteract the low osmotic potential of the soil solution out side. The lost energy results in reduced growth (Brady and Weil, 2002). Increase in of cell sap pH and EC can be considered as a protective strategy to withstand sodicity stress. This may be due to the synthesis and accumulation of organic acids, which regulate the pH and EC of cell sap (Dwivedi et al., 1975) and may, helped to maintain osmoticum to withstand sodicity stress. The decreased water potential and SWC and increase in WSD reveals that plant experienced water stress that could be attributed to impedance in water uptake. Similar observations were made with Salicornia rubra and Atriplex griffithii var stocksii plants where water potential decreased with increase in salinity (Khan et al., 2000, 2001). Increased accumulation of proline in the leaf tissue might be caused by increase in WSD in plants exposed to higher sodicity stress. Proline is accumulated in water stress (Carceller and Fraschina, 1980). Decreased yield in terms of biomass is related to reduction in height (Graifenberg et al., 1996). The decrease in dry matter production may also due to decreased photosynthetic capacity of plant. Photosynthetic efficiency is related to the photosynthetic pigments of plants grown at high ESP, which showed decrease in the present study. Both forms of chlorophyll a and b along with carotenoids declined, such decreases in chlorophyll a and b contents as a result of increasing of soil sodicity has been observed by several workers (El-Sharkawi et al., 1986; Tewari and Singh, 1991). This reduction in chlorophyll content may be related to the enhanced activity of chlorophyllase (Reddy and Vora, 1986). According to Lapina and Popov (1970) salt stress conditions have been found to disrupt fine structure of the chlorophyll and unstability of the pigment protein complex resulting in to reduced chlorophyll contents. Carotenoids scavenge free radicals that are generated owing to excess excitation energy from chlorophyll during photosynthesis (Arora et al., 2002).
Oxidative stress is due to high built up of active oxygen species and is also detriment for plants grown at high ESP faced oxidative stress is indicated by increase carotenoids/chlorophyll ratio and increased activity of catalase, an antioxidative enzyme. The increased carotenoids/chlorophyll ratio is suggested to protect the pigment protein complex of reaction center (Perez-Gelvez and Minguez-Mosquera, 2002). According to Fahmey et al. (1998), the activities of anti-oxidant enzymes (catalase, peroxidase and glutathione reductase) were increased in cells as a result of increasing salt concentration. They stated that the highest proportional increase in enzyme activity compared with control was exhibited by catalase, followed by peroxidase and glutathione reductase. The high levels of enzyme activities are involved in salt tolerance and in mitigating the impaired oxidative metabolism resulting from salt stress. However, activity of peroxidase was decreased. Similar observation was made by Mittal and Dubey (1992). They stated that increased levels of salinity caused significant increase in peroxidase activity in sensitive cultivars of rice whereas in tolerant the activity decreased under salinization. Salt sensitive cultivars always maintained higher levels of peroxidase specific activity in embryo axis compared to tolerant under both controls as well as salt treatments. Peroxidase is known to be actively associated with growth and developmental process and the changes in their activity are regarded as characteristics in relation to chemically and physically controlled growth and developmental process in plants (Gaspar et al., 1985). In present study, tolerance of fennel crop to soil sodicity appears to be due to increased activity of catalase that decomposed H2O2 that might have been formed due to altered ionic status of the cell.
Superoxide dismutase (SOD) is an important antioxidant enzyme and is the first line of defense against oxidative stress in plants. SOD causes dismutation of superoxide radicals at almost diffusion- limited rates to produce H2O2 (Salin, 1987). It plays an important part in determining the concentration of O2 and H2O2 in plants and hence performs a key role in the defense mechanism against free-radical toxicity (Bowler et al., 1992). In the present study, SOD was significantly increased than the control on increasing the sodicity. Maximum increment was observed at high ESP. Similar trend were also observed by several workers. According to him salt stress in cultivars differing in salt tolerance has revealed increased SOD activity in salt-tolerant genotypes of pea, cotton and tomato and induction of SOD enzyme activity was suggested as a reason for improved tolerance to salinity in these cases (Gosset et al., 1994; Hernandez et al., 1999; Mittova et al., 2003). Steep increase in total SOD activity levels has been recorded in Bruguiera gymnorrhiza and B. parviflora during salt stress (Takemura et al., 2000; Parida et al., 2004).
Decreased activity of nitrate reductase with increasing ESP can be a result of decreased biosynthesis or enhanced degradation of enzyme. It has been reported that salt stress causes a shift of ribosomes from the polymeric to the monomeric form in maize seedlings affecting the biosynthesis process of enzyme (Hsiao, 1973). However, decrease due to enhanced degradation of nitrate reductase by an inactivating system appears to be most probable and has been suggested by plant. The presence of such inactivating system for the control of RNA has already been reported by Trogisch et al. (1989). Although it is not yet confirmed whether nitrate reductase is an adoptive enzymes with synthesis being induced by NO3¯ via gene activation (Travis et al., 1969; Breteles et al., 1978), there is general agreement that nitrate enhances nitrate reductase activity (Benzioni et al. 1971). Another possibility for decreased enzymatic activities under salt stress may be because of limited substrate availability in the leaves, resulting from inhibition of NO3¯ uptake (Lacuesta et al., 1990). Although, Martinez and Cerda (1989) could not find any relationship between NO3¯ content of the leaves and enzymes activity, several investigators have emphasized the importance of efflux of vacuolar NO3¯ ions and movement of NO3¯ ions from the roots to the shoots under the influence of salt stress (Aslam and Huffaker, 1989). Although Na+ and K+ ions are essential for the synthesis and activity of nitrate reductase, their salts are strong inhibitors, which is one of the reasons for lower activity of nitrate reductase under salt stress. Decreased activity of nitrate reductase may have hampered the process of nitrogen assimilation. This proposition derives support from the data obtained for soluble protein, soluble protein content decreased with increase in soil ESP.
The analysis regarding elements indicates that the outcome was primarily a function of availability of concerned element in the soil or soil solution. The sodium concentration increased in different plant parts of fennel on increasing the soil ESP. The increase may be correlated with the data obtained for soil where it may be seen that increasing ESP increased exchangeable Na. Graifenberg et al. (1996) also observed that in fennel the Na concentration increased on increasing the salinity. The accumulation of Na was greater in leaves than in shoots or roots (Cherian and Reddy, 2000). Increase of Na in the plants is inevitable when exposed to salt stress. Salt tolerant varieties of crops like barley, linseed, mustard, rice, safflower and wheat have shown less accumulation of Na compared with sensitive ones. The reduction in K uptake in tolerant varieties were minimum resulting in low Na:K ratio (Janardhan et al. 1986; Singh and Singh, 1990; Chhipa and Lal 1995). But Kantian (1975) found higher Na accumulation in tolerant rice variety compared with sensitive one. In the present study, the Na concentration and accumulation was higher and can be taken as an index for its tolerance. Plants growing under sodic/saline condition often show an increase in Na contents in shoots and are accompanied by a decrease in other essential elements (Afridi et al., 1983; Qadar, 1995). Similarly, Chabra et al. (1979) found that increase in the Na concentration was accompanied by decrease in Ca concentration as the ESP increased while there was no effect on the Mg contents of the plant parts.
The content of essential oil was significantly increased on increasing the soil ESP in present study. An increase in trans-anethole content in fennel essential oil might be attributed to decline in the primary metabolites due to the effect of sodicity, causing intermediary products to become available for secondary metabolite synthesis. This had already been observed in some of the salt stressed plants (Morales et al., 1993). In other plants similar results were obtained particularly in mentha by Prasad et al. (1996) while in chamomilla, though oil content was decreased due to sodicity but few of its constituents e.g., chameazulene and bisabolol oxide-B in oil increased (Ram et al., 1999). The main constituents of essential oil of fennel seed are trans-anethole, d-fenchone, methylchavicol and limonene in which trans-anethole and methylchavicol are phenylpropanoids while d-fenchone is bicyclic monoterpines. The percent contents of phenylpropanoids (trans-anethole and methylchavicol) showed reciprocal trends against the percent contents of the bicyclic monoterpines (d-fenchone) at different ESP. The effects of soil sodicity on production of phenylpropanoids and monoterpines can be explained on the basis of different primary metabolic pathways of carbon. Phenylpropanoids (trans-Anethole and methylchavicol) biosynthesized from phosphoenolpyruvate (PEP) and d-erythrose-4-phosphate via the shikimic acid pathway. While monoterpene (d-fenchone) are biosynthesized from acetyl-CoA via the mevalonate pathway. The common biosynthetic point of the two pathways is that PEP is a precursor of acetyl CoA via pyruvate. It is suggested that PEP is being diverted into the shikimate pathway for production of the phenylpropanoids in sodic stress condition. Similar suggestions have been made by Dewick (1997) and Robbers et al. (1996).
CONCLUSION
Considering the growing demands of spices, medicinal and aromatic crops in the global market and efficient utilization of the productive lands for food grain production to meet the demands of increasing population, it has become imperative to utilize unproductive barren sodic lands for growing fennel crops. Due to higher yield of essential oil contents and improved quality (increased in trans-anethole content and decrease in d-fenchone content) may compensate with reduced seed yield.
ACKNOWLEDGMENT
The author is thankful to Dr. V.K. Garg, Retired Scientist, National Botanical Research Institute, Lucknow for proper guidance and support. I am also thankful to Department of Science and Technology, New Delhi for financial assistance through SERC Fast Track Scheme (SR/FT/L-172/2004).
REFERENCES
- Abdallah, N., EI-Gengaihi and E. Sedrak, 1998. The effect of fertilizer treatment on yield of seed and volatile oil of fennel (Foeniculum vulgare Mill). Pharmazie, 33: 607-608.
PubMed - Alscher, R.G., J.L. Donahue and C.L. Cramer, 1997. Reactive oxygen species and antioxidants: Relationship in green cells. Physiol. Plant., 100: 224-233.
CrossRefDirect Link - Arora, A., R.K. Sairam and G.C. Srivastava, 2002. Oxidative stress and antioxidative systems in plants. Cur. Sci., 82: 1227-1238.
Direct Link - Aslam, M. and R.C. Huffaker, 1989. Role of nitrate and nitrite in the induction of nitrate reductase in leaves of barley seedling. Plant Physiol., 91: 1152-1156.
Direct Link - Barrs, H.D. and P.E. Weatherley, 1962. A re-examination of the relative turgidity technique for estimating water deficits in leaves. Aust. J. Biol. Sci., 15: 413-428.
Direct Link - Benzioni, A., Y. Vaadia and S.H. Lips, 1971. Nitrate uptake by roots as regulated by nitrate reduction products in the shoot. Physiol. Plant., 28: 288-290.
CrossRef - Bernstein, L., 1975. Effects of salinity and sodicity on plant growth. Ann. Rev. Phytopathol., 13: 295-312.
CrossRefDirect Link - Bhandal, I.S. and C.P. Malik, 1988. Potassium estimation, uptake and its role in the physiology and metabolism of flowering plants. Int. Rev. Cytol., 110: 205-254.
CrossRefDirect Link - Bowler, C., M.V. Montagu and D. Inze, 1992. Superoxide dismutase and stress tolerance. Annu. Rev. Plant Physiol. Mol. Biol., 43: 83-116.
CrossRefDirect Link - Breteles, H., C.H. Hamish and T. Cate, 1978. Ionic balance of root shoot nitrate transfer in dwarf bean. Physiol. Plant., 42: 53-56.
CrossRef - Cherian, S. and M.P. Reddy, 2000. Salt tolerance in the halophyte Suaeda nudiflora MOQ: Effect of NaCl on growth ion accumulation and oxidative enzymes. Indian J. Plant Physiol., 5: 32-37.
Direct Link - Chhipa, B.R. and P. Lal, 1995. Na/K ratio as the basis of salt tolerance in wheat. Aust. J. Agric. Res., 46: 533-539.
CrossRefDirect Link - Clevenger, J.F., 1928. Apparatus for the determination of volatile oil. J. Am. Pharm. Assoc., 17: 345-349.
CrossRefDirect Link - Dwivedi, R.S., N.S. Randhawa and R.L. Bansal, 1975. Phosphorus-zinc interaction. Plant Soil, 43: 639-648.
Direct Link - El-Sharkawi, H.M., F.M. Salame and A.A. Mazen, 1986. Chlorophyll response to salinity, sodicity and heat stresses in cotton, rama and millet. Photosynthetica, 20: 204-211.
Direct Link - Epstein, E., 1998. How calcium enhances plant salt tolerance. Science, 280: 1906-1907.
CrossRefDirect Link - Euler, H.V. and K. Josephson, 1927. Uber katalase. I. Justus Liebigs Ann. Chem., 452: 158-181.
CrossRefDirect Link - Foyer, C.H., H. Lopez-Delgado, J.F. Dat and I.M. Scot, 1997. Hydrogen peroxide- and glutathione-associated mechanisms of acclimatory stress tolerance and signalling. Physiol. Plant., 100: 241-254.
CrossRefDirect Link - Perez-Gelvez, A. and M.I. Minguez-Mosquera, 2002. Degradation of non-esterified and esterified xanthophylls by free radicals. Biochim. Biophys. Acta (BBA)- Gen. subj., 1569: 31-34.
CrossRef - Garg, V.K., P.K. Singh and A. Mathur, 2000. Characterization and classification of sodic soils of Gangetic alluvial plains at banthra Lucknow. Agropedology, 10: 163-172.
Direct Link - Garg, V.K., 2012. Sodicity affects growth yield and cation composition of fennel (Foeniculum vulgare Mill.). J. Spices Arom. Crops, 21: 130-135.
Direct Link - Garg, V.K., P.K. Singh and P. Pushpangadan, 2005. Exchangeable sodium induced changes in yield, water relation and cation composition of fennel (Foeniculum vulgare Mill). J. Environ. Biol., 26: 335-340.
PubMed - Gaspar, T., C. Penel, F.J. Castillo and H. Greppin, 1985. A two-step control of basic and acidic peroxidases and its significance for growth and development. Physiol. Plant., 64: 418-423.
CrossRef - Gosset, D.R., E.P. Millhollon and M.C. Lucas, 1994. Antioxidant response to NaCl stress in salt-tolerant and salt-sensitive cultivars of cotton. Crop Sci., 34: 706-714.
CrossRefDirect Link - Graifenberg, A., L. Botrini, L. Giustiniani, M. Lipucci and D. Paola, 1996. Salinity effects growth, yield and elemental concentration of fennel. HortScience, 31: 1131-1134.
Direct Link - Hsiao, T.C., 1973. Rapid changes in levels of polyribosomes in Zea mays in response to water stress. Plant Physiol., 46: 281-288.
CrossRef - Hernandez, J.A., A. Campillo, A. Jimenez, J.J. Alarcon and F. Sevilla, 1999. Response of antioxidant systems and leaf water relations to NaCl stress in pea plants. New Phytol., 141: 241-251.
CrossRefDirect Link - Hageman, R.H. and A.J. Reed, 1980. Nitrate reductase from higher plants. Methods Enzymol., 69: 270-280.
CrossRef - Janardhan, K.V., B.N. Patil and D.S. Raiker, 1986. Relative tolerance of safflower (Carthamus tinctorius, L) varieties to saline water irrigation. Indian J. Plant Physiol., 29: 118-124.
Direct Link - Kantian, S., 1975. Significance of salt transport patterns in rice varieties differing in salt tolerance. Communi. Soil Sci. Plant Anal., 6: 63-69.
CrossRef - Khan, M.A., B. Gul and D.J. Weber, 2001. Effect of salinity on the growth and ion content of Salicornia rubra. Commun. Soil Sci. Plant Anal., 32: 2965-2977.
CrossRefDirect Link - Khan, M.A., A.U. Irwin and A.M. Showalter, 2000. Effects of salinity on growth, water relations and ion accumulation of the subtropical perennial halophyte, Atriplex griffithii var. stocksii. Ann. Bot., 85: 225-232.
CrossRefDirect Link - Lacuesta, M., B Gonzalez-Moro, C. Gonzale-Murua and A. Munoz-Rueda, 1990. Temporal study of the effect of phosphinothricin on the activity of glutamine synthetase glutamate dehydrogenase and nitrate reductase in Medicago sativa L. J. Plant. Physiol., 136: 410-414.
Direct Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Maas, E.V. and G.J. Hoffman, 1977. Crop salt tolerance-current assessment. J. Irrig. Drainage Div., 103: 115-134.
Direct Link - Mangal, J.L., A. Yadava and G.P. Singh, 1986. Effect of different levels of soil salinity on germination, growth, yield and quality of coriander and fennel. South Indian Hort., 34: 26-31.
Direct Link - Martinez, V. and A. Cerda, 1989. Nitrate reductase activity in tomato and cucumber leaves as influenced by NaCl and N source. J. Plant. Nutr., 12: 1335-1350.
Direct Link - Mittal, R. and R.S. Dubey, 1992. Mitochondrial acid phosphatase and adenosine triphosphatase activities in germinating rice seeds following NaCl salinity stress. Indian J. Plant Physiol., 35: 174-181.
Direct Link - Mittova, V., M. Tal, M. Volokita and M. Guy, 2003. Up-regulation of the leaf mitochondrial and peroxisomal antioxidative systems in response to salt-induced oxidative stress in the wild salt-tolerant tomato species Lycopersicon pennellii. Plant Cell Environ., 26: 845-856.
CrossRefDirect Link - Morales, C., R.M. Cusido, J. Palazon and M. Bonfill, 1993. Response of Digitalis purpurea plants to temporary salinity. J. Plant Nutr., 16: 327-335.
CrossRef - Parida, A.K., A.B. Das and P. Mohanty, 2004. Defense potentials to NaCl in a mangrove, Bruguiera parviflora: Differential changes of isoforms of some antioxidative enzymes. J. Plant Physiol., 161: 531-542.
CrossRefPubMedDirect Link - Qadar, A., 1995. Potassium and sodium contents of shoot and laminac of rice cultivars and their sodicity tolerance. J. Plant Nutr., 18: 2281-2290.
Direct Link - Reddy, M.P. and A.B. Vora, 1986. Changes in pigment composition, Hill reaction activity and saccharides metabolism in Bajra (Pennisetum typhoides S & H) leaves under NaCl salinity. Photosynthetica, 20: 50-55.
Direct Link - Richards, L.A., 1954. Diagnosis and Improvement of Saline and Alkali Soils. United State Government Printing Office, Washington, DC., USA., Pages: 160.
Direct Link - Salin, M.L., 1988. Toxic oxygen species and protective systems of the chloroplast. Physiol. Plant., 72: 681-689.
CrossRefDirect Link - Singh, P.K., A.R. Chowdhury and V.K. Garg, 2002. Yield and analysis of essential oil of some spice crops grown in sodic soils. Indian Perf., 46: 35-40.
Direct Link - Singh, S.B. and M.V. Singh, 1990. Effect of exchangeable sodium on the growth yield and mineral composition of barley varieties. J. Indian Soc. Soil Sci., 38: 135-138.
Direct Link - Singh, S.B. and I.P. Abrol, 1985. Effect of exchangeable sodium percentage on growth yield and chemical composition of onion and garlic. J. Indian Soc. Soil Sci., 33: 358-361.
Direct Link - Takemura, T., N. Hanagata, K. Sugihara, S. Baba, I. Karube and Z. Dubinsky, 2000. Physiological and biochemical responses to salt stress in the mangrove, Bruguiera gymnorrhiza. Aquat. Bot., 68: 15-28.
CrossRefDirect Link - Travis, R.L., W.R. Jordan and R.C. Huffaker, 1969. Evidence for an inactivating system of nitrare reductase in Hordeum vulgare L. during darkness that requires protein synthesis. Plant Physiol., 44: 1150-1156.
PubMed - Yadav, I.S.P., 1993. Salt affected soils and their management with special reference to Uttar Pradesh. J. Indian Soc. Soil Sci., 41: 623-629.
Direct Link - Bates, L.S., R.P. Waldren and I.D. Teare, 1973. Rapid determination of free proline for water-stress studies. Plant Soil, 39: 205-207.
CrossRefDirect Link - Tewari, T.N. and B.B. Singh, 1991. Stress studies in lentil (Lens esculenta Moench). II. Sodicity-induced changes in chlorophyll, nitrate and nitrite reductase, nucleic acids, proline, yield and yield components in lentil. Plant Soil, 136: 225-230.
CrossRefDirect Link - Beauchamp, C. and I. Fridovich, 1971. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem., 44: 276-287.
CrossRefPubMedDirect Link