
Alka Upadhyay
University of Lucknow, Lucknow, Uttar Pradesh, India
Satyendra Tripathi
Department of Civil Engineering, Birla Institute of Technology and Science, Pilani, Hyderabad Campus, 500078, India
S.N. Pandey
University of Lucknow, Lucknow, Uttar Pradesh, India
Research Journal of Soil Biology
Year: 2012 | Volume: 4 | Issue: 3 | Page No.: 69-80







ABSTRACT
Bishops weed-Ammi majus L. (Apiaceae) is one of the richest natural sources of linear coumarins and furanocoumarins; umbelliferone, psoralen, xanthotoxin, bergapten and imperatorin and having great medicinal value. The fruit of A. majus has been used in the Mediterranean and bordering regions in the treatment of leucoderma, psoriasis, vitiligo and now a days in cancer and AIDS treatments also. Clay pot culture was conducted to know the response of plant to salt stress, growth and biochemical responses and nutrients uptake in different soil sodicity. The results showed inhibited growth responses with increasing sodicity of soil. Also, decrease in chlorophyll contents a, b and carotenoid contents were also observed. Further changes in the activities of catalase were obtained. On contrary a reverse trend for peroxidase was observed in leaves with increased ESP levels. Sodium content in plants was increased, while potassium and zinc was decreased in different parts of plant with increase in the soil sodicity. Copper varied differently in different plant parts of plant. These observations showed the inhibition in the growth responses due to sodicity stress.
PDF Abstract XML References Citation
Received: April 11, 2012;
Accepted: July 06, 2012;
Published: February 13, 2013
How to cite this article
Alka Upadhyay, Satyendra Tripathi and S.N. Pandey, 2012. Effects of soil sodicity on growth, nutrients uptake and Bio-chemical responses of Ammi majus L. Research Journal of Soil Biology, 4: 69-80.
DOI: 10.3923/rjsb.2012.69.80
URL: https://scialert.net/abstract/?doi=rjsb.2012.69.80
DOI: 10.3923/rjsb.2012.69.80
URL: https://scialert.net/abstract/?doi=rjsb.2012.69.80
INTRODUCTION
Ammi majus L. commonly grown in temperatures range of 68-72 F (20-22°C) and belongs to the family Umbelliferae and locally known as atrilal. Geographic origin of Ammi majus L. is Southern Europe, Turkey and North Africa. Its average height is 12-36 inches (30-90 cm). Flowering attributes is performed in compound umbels bearing about 10 small white flowers per umbel, resembling lace work. Two to three pinnate, light green, lance-shaped leaves are attributed for culture works. It needs full sun to partial shade for proper growth. Germination normally occurs within 10-14 days.
Sodic soils occurring in the Indo-gangetic plains in northern India (Abrol and Bhumbla, 1971) and many other parts of the world (Elgabaly, 1971; Szabolcs, 1977), contain a considerable quantity of sodium carbonate in addition to high amounts of exchangeable sodium. The problem of sodicity is associated with elevated soil pH and under these conditions the availability of certain plant nutrients is reduced resulting in to severe loss in crop production (Sharma, 2006). Elevated Na+ concentration in soils is also highly toxic to many plants, although tolerance levels vary between different plant species. The excess sodium and high pH impart adverse soil physicalproperties leading to poor air-water- plant relationships (Acharya and Abrol, 1975) and crop failure on alkali soils, results largely from toxicity of sodium carbonate and bicarbonate and osmotic effect of other salts present. Excess sodium imparts poor physical properties to soils (Mehrotra and Agrawala, 1979). These soils have, therefore, extremely low permeability, are puddle easily and upon drying form compact blocks that may present a physical barrier to seed germination and root penetration. Soluble carbonates result in high soil pH (Mashhady and Rowell, 1978). This in turn brings about nutritional imbalances in the soil which prevent or limit the growth of most crops (Qadir et al., 2007; Garg and Malhotra, 2008). Salt problems, which commonly occur in arid and semi-arid irrigated areas (Barbiero et al., 2001), lower the productivity of extensive areas of agricultural land throughout the world (Dahiya and Dahiya, 1977).
MATERIALS AND METHODS
Experiment was conducted to study the effects of soil sodicity (ESP 4.6, 17.9, 33.5 and 57.4) on growth, nutrients uptake and biochemical responses of plants Ammi majus L during Nov 2006 to May 2007. Growth (height, branching and flowering) observations were taken at 45, 75 and 105 DAS (Days After Sowing). Pigment contents (chlorophyll and carotenoid) and enzymes activity (catalase and peroxidase) in leaves were determined at 70 DAS. Uptake and tissue concentration of some nutrient elements (Na, K, Zn and Cu) were also analyzed in different plant parts (leaf, stem and root) of A. majus.
Clay pot culture: Earthen clay pots (15 kg soil size) were filled with sodic soils, used for growing the test plant atrilal. Clay pots were lined their inner side by alkathene sheet to check the leaching and contamination from the clay of the pots.
Experiments were carried out in triplicate for each treatment.
Collection of soil: The sodic soils were collected from the different location of Lucknow district with surface soil (0-15 cm depth) samples (Piper, 1967).
Preparation of soil: The collected soils from the different site up to 15 cm depth (surface soils) only kept it for drying. After drying, the soils were thoroughly crushed and properly mixed. The earthen clay pots which were lined on their inner side by alkathene sheet, 15 kg soils were filled in each pot. 20 Seeds of test plants Atrilal were sown in sodic soils of four level (S1, S2, S3, S4) of Exchangeable Sodium Percent (ESP) in triplicate. Experiment was set up in Complete Randomized Design (CRD). The climatic conditions was observed regularly light, temperature, rainfall etc. Thinning was done after 30 days to allow 10 plants to grow in each pot. The properties of soil have been shown in Table 1.
Growth observations: The observation was recorded at successive stages of growth and yield to make critical analysis of crop as affected by different treatments. The heights of the five representative plants from each pot were measured in centimeters from Ground level up to the highest level reached by the leaves. Number of branches, flowers and fruits were counted from five randomly selected plants from each pot and averaged.
Plant analysis: Harvested plant sample of Ammi majus L. at 150 days of sowing were analyzed for mineral concentration. Plant sample were dried at 700°C for 48 h in oven.
| Table 1: | Properties of soils before sowing the test plants |
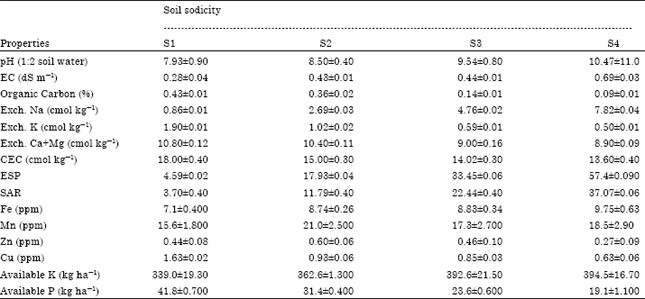 | |
| EC: Electrical conductivity, ESP: Exchangeable sodium percentage, SAR: Sodium absorption ratio, CEC: Cation exchange capacity, Exch.: Exchangeable | |
For homogenization, these were ground in stainless steel grinder and subsequently used for chemical analysis. Processed plant samples were digested in diacid mixture (HNO3+HClO3) (2:1) ratio on hot plate under controlled temperature for estimation of Na, K, Zn and Cu. Na, K by flame photometrically as described by Piper (1967) and Zn, Cu were determined by Atomic absorption spectrophotometer Analist-300 Perkin Elmer. The uptakes of K, Na, Cu and Zn in the Plant sample (Ammi majus L.) were calculated by multiplying nutrient concentration with dry matter yield (g plant-1).
Enzyme assay: Catalase was assayed by the modified method of Bisht (1972). Enzyme activity is expressed as mole H2O2 reduced per unit fresh matter or protein weight. Modification of the method given by Luck (1963) was used for peroxidase assay.
Chlorophyll: Method given by Arnon (1949) was used to determine chlorophyll contents. The pigment concentrations were calculated using following numerical expressions with specific absorption coefficient of McKinney (1940) for chlorophyll in green leaves:
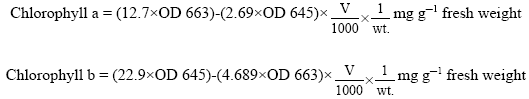 |
| (1) |
Where:
| (2) |
Statistical analysis: The one-way ANOVA and F-test has been used to determine statistical significance. where:
| (3) |
MSB = The mean square between groups (explained variance)
MSE = The mean square within groups (error variance)
RESULTS
Growth attributes: Effect of soil sodicity on vegetative growth of plant Ammi majus L. has been shown in Table 2. The increase in ESP (exchangeable sodium percentage) markedly suppressed the plant growth observed at 45, 75 and 105 DAS. The increase in ESP from 4.6 to 57.4 produced significant effects on plant height. Decreases in plant height at different ESP levels were observed. The maximum reduction in plant height was found at high ESP level (57.4). With increase the ESP levels, non-significant difference was observed in number of branch. Flowering was not started up to 45.
DAS and firstly appeared in control plants. The decrease in plant height and flowering were significant statistically, but branching was non-significant at 75 DAS. At the mature stage (105 DAS), plant showed decrease in height and flowering by 61 and 85%, respectively, grown at high ESP level (57.4). Growths of test plants were increased with increase in the duration of exposure at all ESP levels, whereas growth rate was decreased with increase in sodicity and time exposure. Maximum growth responses of plants were observed grown in non-sodic soil (ESP.5). Biomass of the plant decreased significantly also showed. The average fresh weight of the A. majus showed 20, 23.33 and 57.50% decrease over the control at ESP 17.9, 33.5 and 57.4, respectively. Dry weight also decreases by 27.71, 31.33 and 60.24% with increasing the soil sodicity.
| Table 2: | Effect of soil sodicity on vegetative growth of plant Ammi majus L |
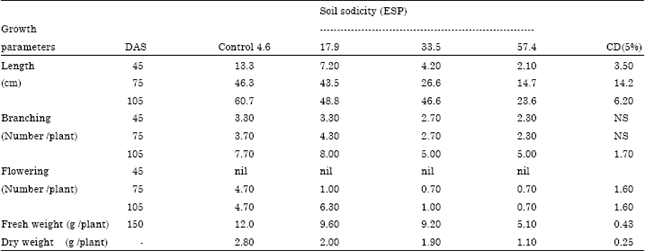 | |
| Table 3: | Effect of soil sodicity on photosynthetic pigments in Ammi majus L. leaves |
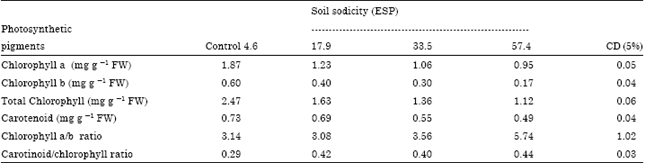 | |
| Table 4: | Effect of soil sodicity on enzymes activity (catalase and peroxidase) in Ammi majus L. leaves |
 | |
Biochemical responses
Photosynthetic pigments: A significant decrease in the chlorophyll (a, b and total) and carotenoids contents was observed in A. majus leaves grown in sodic soils (ESP 17.93, 33.5 and 57.4) as shown in Table 3. Decrease in chlorophyll a, b, total and carotenoid contents were observed by 49, 71, 54 and 34%, respectively at high sodic soil (ESP 57.4) with respect to control. Significant increase was found in the ratio of chlorophyll a and b at ESP 33.5 and 57.4, respectively, whereas, carotinoid/chlorophyll ratio increased significantly with increasing ESP level in soil.
Enzyme activities: A significant increase in the activities of peroxidase was observed in leaves with increased ESP levels. On contrary a reverse trend was obtained for catalase. Increase in peroxidase activity by 26, 33 and 54% was analyzed at 17.9, 33.5, 57.4 ESP levels, respectively, as compared to control (ESP 4.6). On the other hand, decrease in catalase activity by 20.8, 27.4 and 32.3% was found at 17.9, 33.5 and 57.4 ESP levels, respectively as shown in Table 4.
Nutrients uptake: Some nutritional uptake like Sodium, Potassium, Zinc and Copper by plant part (leaf, root and stem) are shown in Table 5-8.
Sodium (Na): Sodium content in plants was significantly increased with increase in the soil sodicity. Plants showed maximum uptake and accumulation of sodium at high ESP (57.4) in soil. Tissue concentration of Na was found to be more in root in compared to other plant parts (stem and leaves). Study revealed that uptake and accumulation of Na in A. majus were increased with increased exchangeable Na concentration in soil. Increased Na accumulation in leaf by 26, 40 and 44.6% were determined with gradual increase in the ESP in soil. 1.70, 8.48 and 33% increase in root and 12, 51 and 137% increase in stem was obtained (Table 5).
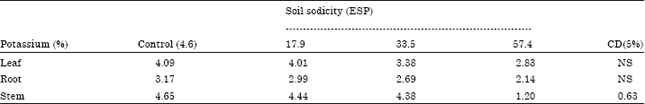
Potassium (K): Tissue concentration of potassium was significantly decreased in different parts of plant A. majus with increase in soil ESP levels. Maximum concentration of potassium was quantified in control (ESP 4.6) plants and gradually decreased with increased ESP levels.
| Table 5: | Effect of soil sodicity on accumulation of sodium |
 | |
| Table 6: | Effect of soil sodicity on potassium concentration |
 | |
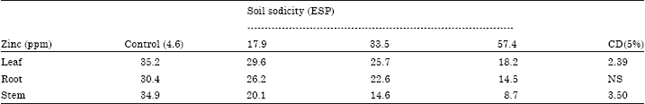
| Table 7: | Effect of soil sodicity on zinc concentration |
 | |
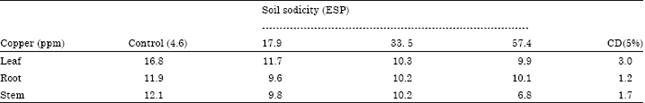
| Table 8: | Effect of soil sodicity on copper concentration |
 | |
Gradual decrease in potassium accumulation in leaf and root was not significant up to 33.5 ESP but at ESP 57.4, decrease was statistically significant in the root and leaf (Table 6).
Zinc (Zn): Tissue concentration of zinc was found to be decreased with increasing soil ESP level in different plant parts of A. majus. The maximum accumulation was determined in control plants. Uptake and accumulation of Zn was more in root as compared to shoot. A significant decrease in Zn accumulation in leaf and stem was obtained, while decrease in root was non- significant statistically. The respective decrease at ESP 17.9, 33.5 and 57.4 was observed by 15.8, 27.0, 48.3 in leaf; 13.8, 25.6, 52.4% in root and 42.5, 58.4, 75.0% in stem respectively (Table 7).
Copper (Cu): Tissue concentration of copper varied differently in different plant parts of A. majus. Maximum tissue concentration of Cu (16.77 ppm) was found in leaf of plants. Significant decrease in Cu accumulation in root was obtained at ESP 17.9 and 57.4 levels as compared to control, but difference were non-significant between 17.9 to 33.5 and 57.4 ESP levels. Maximum decline in tissue concentration of Cu was found at high ESP (57.4) level (Table 8).
DISCUSSION
Increasing ESP (exchangeable sodium %age) profoundly affected the survival, growth and biochemical responses of Ammi majus L. (atrilal). Detrimental effects of increased ESP levels in sodic soil were found on the plants height. Reduction in growth of test plants as measured by differences in plant height and branching/leaves, growing on sodic soils as compared to similar plants growing on normal soils (ESP 4.6) are consistent with observation on vetiver by Anwar et al. (1996), observed similar height reduction of vetiver plants growing in sodic soil under greenhouse conditions. Dinesh et al. (2004) also observed decrease in height of plants, branching/leaves and flowers with increase in soil ESP. Relative growth rate of many agricultural crops has been observed to be retarded under sodic conditions in soil (Hocking, 1993; Gill and Qadar, 1998). A. majus appeared more tolerant to soils sodicity with less reduction in growth of plants growing on sodic soil (ESP 33.5). The reduction in plant growth could be attributed might be due to the presence of high Na content in soil in available form (Garg and Malhotra, 2008). High Na concentration that lead to increase osmotic potential of the circulating soils solution as well as due to ionic imbalance stress resulting from excessive uptake of Na ions (Greenway and Munns, 1980; Garg and Malhotra, 2008). Toxic effects of Na have been also determined by Trapp et al. (2008), reported decrease in biomass is related to reduction in plant height, branches/leaves and flowers/ fruits with exposure of high Na in soil. Increasing salt levels of the growth medium caused a marked inhibitory effect on fresh and dry weights and maximum reduction in biomass was observed at highest salt level (Ashraf and Orooj, 2006). Decrease in plant height, branching/leaves production, relative growth rate and biomass due to high sodic condition have been reported for many crops (Soliman and Doss, 1992; Yadav et al., 1996; Singh and Singh, 1997; Dua, 1998; Prasad et al., 2006). A. majus appeared to be tolerant species of soil sodicity (ESP 33.5) with less reduction in growth of plants than high sodic soil (ESP 57.4). The medicinal plants grown on non-sodic (control) soil showed better growth than the soils contained high exchangeable Na. With increase in exchangeable sodium percentage levels, there was a continuous decrease in chlorophyll a and b content along with carotenoids in leaves of A. majus observed. Similar findings were also reported by Tewari and Singh (1991) in lentil. Lapina and Popov (1970) reported that, salt stress conditions have been found to disrupt fine structure of the chlorophyll and instability of pigment protein complex, resulting in to reduction of chlorophyll contents. Also, reduction in pigment contents could be attributed due to Na-induced oxidative stress in plants (Kennedy and de Filippis, 1999) Increased intracellular Na concentration is believed to accelerate oxidative stress. Oxidative stress is due to the high build up of active oxygen species and cause detrimental effects on plants grown at high ESP (>10) faced oxidative stress is indicated by increased carotenoids/chlorophyll ratio and increased activity of catalase, an antioxidative enzyme. The increased carotenoids/ chlorophyll ratio protects the pigment protein complex of reaction center (Perez-Vich et al., 2002). According to Fahmey et al. (1998), the activities of antioxidant enzyme (catalase and peroxidase) were increased in cells as a result of increased salt concentration. Increased peroxidase and decreased catalase activity of A. majus, were observed. Mittal and Dubey (1991), reported that peroxidase activity decreased with increase levels of salt in salt tolerant plant species. Also they stated that increased levels of salinity caused significant increase in peroxidase activity in sensitive cultivars of rice (CSR-1 and CSR-3) whereas in tolerant, the activity decreased under salinization. Salt sensitive cultivars always maintained higher levels of peroxidase compared to tolerant. Thus, enhanced activities of catalase and reduced activity of peroxidase in sodic soil confirm tolerance against sodic stress conditions. These findings confirmed the salt sensitive to A. majus. The results are also in accordance with Tewari and Singh (1991). Peroxides are known to be actively associated with growth and development process (Gaspar et al., 1985). Activities of antioxidant enzymes (CAT and POX) are very effective in discriminating the canola cultivars for salt tolerance (Ashraf and Ali, 2008) have been reported. Parihar and Kumar (2002) also found the decrease in the catalase activity with increase in ESP level above 10. Tissue concentration of elements related with the availability of concerned element in the soil, where plant was grown. The sodium concentration increased in different plant parts of A. majus on increasing the soil sodicity. This increase may be correlated with soil where it may be seen that increasing ESP increased exchangeable Na (Marschner, 1995). A consistent increase in Na+ and decrease in K+, in both roots shoots and leaf of A. majus were found with an increase in salt concentration into the growth medium in consonance with Ashraf and Orooj (2006). Accumulation of toxic ions has earlier been reported in a number of plant species (Singh and Singh, 1990; Abha et al., 1991; Rajpar and Wright, 2000) referred to as salt accumulators, barley is also one of them (Munns et al., 2000). It has been generally observed that plants exposed to saline environment (NaCl), take up high amounts of Na+, whereas the uptake of K+, is significantly reduced. However, reasonable amounts of K+ are required by plants to maintain the integrity and functioning of cell membranes (Marschner, 1995; Davenport et al., 1997; Wei et al., 2003). Active transport of Na+, through Na+, /H+, antiports (Shi et al., 2003) and other ion channels in plant cell membranes that are likely to have key roles in both toxicity to high salt concentrations (NaCl) and salt tolerance mechanisms (Tyerman and Skerett, 1998; Schachtman and Liu, 1999). The salt tolerance of thousands of species was determined in the hope to find salt-tolerant cash crops (Lieth and Mochtchenko, 2003). Accumulation of Na was greater in roots than in stem and leaves of A. majus. also followed the same trend except at high ESP (57.4) level. The result obtained in the experiment is in agreement with Prasad et al. (2003, 2006) who stated that increase in the soil ESP enhanced the concentration of sodium and decreased that of potassium content in shoot and root tissues of geranium and mint as compared to control. Reduction in growth and biochemical responses could be attributed due to specific accumulation of Na+, in plant tissue which is toxic and may be one of the main reasons for growth inhibitor (Greenway and Munns, 1980). Thus, maintaining a low cytoplasmic Na+, concentration is important for many plants growing in Na affected environments. Plant cells maintain a low cytosolic Na+, concentration presumably through Na+, exclusion, extrusion or compartmentation (Niu et al., 1995). Gorham et al. (1990) reported that the capacity of Na+, exclusion from the shoot is well correlated to the salt tolerance degree. The lemongrass avoided a great Na+, accumulation in shoots as compared to the roots (Patra et al., 2002). Selectivity of K+, over Na+, ion is one of the important traits for salt tolerance (Perez-Alfocea et al., 1996). Salt tolerant varieties of crops like barley, linseed, mustard, rice, sunflower and wheat have shown much less accumulation of Na compared with sensitive ones and with minimum reduction in K uptake, resulting in low Na: K or high K:Na ratio (Chhipa and Lal, 1995; Qadar, 1995). In this present study minimum uptake by A. majus to Na+, showed tolerance behavior to Na. It is also evident that salt tolerance in most plants is associated with low uptake and accumulation of Na+, which is mediated through the regulation of influx and or by active efflux from the cytoplasm to the vacuoles and also back to the growth medium (Blumwald, 2000; Chinnusamy et al., 2005). Abha et al. (1991) also reported that Na content increased significantly in okra with increasing soil ESP, while K content decreased. Differences in nutrients accumulation between plants growing on sodic and normal soils may be due to differences in the availability of nutrients in soil (Pandey et al., 2008). K uptake decreased significantly with increased ESP levels in test plants. Anwar et al. (1996) also observe the same finding in vetiver. Potassium in soil has been reported to be adversely affected by sodicity (Singh et al., 1993; Patra et al., 1998). Soil sodicity adversely effects the growth and nutrients accumulation in aromatic plants has been reported by (Patra et al., 1998). Accumulation of Zn and Cu in the root, shoot and leaves of test plants was decreased on increasing the soil ESP levels. A decrease in uptake of Cu and Zn has been shown in plants grown in salt stress (Singh and Singh, 1994). Prasad et al. (2006) also found the decrease of Zn concentration in plant part with increase in soil ESP. The problem of Zn and Cu accumulation is very acute under sodic conditions where CO-3 and HCO-3 are supposed to be major anions interfering with their availability and may cause precipitation of these nutrients as insoluble salts due to high soil pH. In the changing climate conditions, there is urgent need to study of bio-chemical properties of plants on soil sodicity.
CONCLUSION
Looking in to the increasing demand of aromatic medicinal product, it has been imperative to boost the commercial production of these materials. Since there is already much pressure on production of food, fiber and fodder crops in the good quality fertile lands. The cultivation on these salt affected soils requires special economic agronomic management and amelioration before starting the cultivation. A. majus were found to be moderately tolerant to sodicity and Na excluder. Further study is required to know the best reclamation technology to improve the soil condition to achieve the increasing food demand.
ACKNOWLEDGMENT
The authors are thankful to Department of Botany, University of Lucknow, Lucknow, India to providing infrastructural facilities.
ABBREVIATION
| CD | = | Critical difference |
| CEC | = | Cation exchange capacity |
| cm | = | Centimeter |
| cmol | = | Centmole |
| DAS | = | Days after sowing |
| DW | = | Dry weight |
| EC | = | Electrical conductivity |
| ESP | = | Exchangeable sodium percentage |
| FW | = | Fresh weight |
| g | = | Gram |
| ha | = | Hectare |
| kg | = | Kilogram |
| Meq/L | = | Milliequivalent per liter |
| Mg | = | Milligram |
| ml | = | Milliliter |
| mm | = | Millimeter |
| NS | = | Non-significant |
| ppm | = | Parts per million |
| SAR | = | Sodium adsorption ratio |
| VC | = | Vermicompost |
REFERENCES
- Arnon, D.I., 1949. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol., 24: 1-15.
CrossRefPubMedDirect Link - Ashraf, M. and Q. Ali, 2008. Relative membrane permeability and activities of some antioxidant enzymes as the key determinants of salt tolerance in canola (Brassica napus L.). Environ. Exp. Bot. 63: 266-273.
CrossRef - Ashraf, M. and A. Orooj, 2006. Salt stress effects on growth, ion accumulation and seed oil concentration in an arid zone traditional medicinal plant ajwain (Trachyspermum ammi [L.] Sprague). J. Arid Environ., 64: 209-220.
CrossRefDirect Link - Barbiero, L., V. Valles, A. Regeard and C. Cheverry, 2001. Residual alkalinity as tracer to estimate the changes induced by a forage cultivation in a non-saline irrigated sodic soil. Agric. Water Manage., 50: 229-241.
CrossRef - Blumwald, E., 2000. Sodium transport and salt tolerance in plants. Curr. Opin. Cell Biol., 12: 431-434.
CrossRefDirect Link - Chinnusamy, V., A. Jagendorf and J.K. Zhu, 2005. Understanding and improving salt tolerance in plants. Crop Sci., 45: 437-448.
Direct Link - Chhipa, B.R. and P. Lal, 1995. Na/K ratio as the basis of salt tolerance in wheat. Aust. J. Agric. Res., 46: 533-539.
CrossRefDirect Link - Davenport, R.J., R.J. Reid, F.A. Smith and C.H. Bornman, I997. Sodium-Calcium interactions in two wheat species differing in salinity tolerance. Physiol. Plant., 99: 323-327.
Direct Link - Dinesh, K., K, Singh, H.S. Chauhan, A. Prasad, S.U. Beg and D.V. Singh, 2004. Ameliorative potential of palmarosa for reclamation of sodic soils. Commun. Soil Sci. Plant Anal., 35: 1197-1206.
Direct Link - Gaspar, T., C. Penel, F.J. Castillo and H. Greppin, 1985. A two-step control of basic and acidic peroxidases and its significance for growth and development. Physiol. Plant., 64: 418-423.
CrossRef - Gorham, J., A. Bristol, E.M. Young, R.G. Wyn Jones and G. Kashmir, 1990. Salt tolerance in the Triticeace: K/Na Discrimination in barley. J. Exp. Bot., 41: 1095-1101.
CrossRef - Hocking, P.J., 1993. Distribution and redistribution of mineral nutrients and dry matter in grain sorghum as affected by soil salinity. J. Plant Nutr., 16: 1753-1774.
CrossRef - Kennedy, B.F. and L.F. de Filippis, 1999. Physiological and oxidative response to NaCl of salt tolerant Grevillea ilicifolia and the salt sensitive Grevillea arenaria. J. Plant Physiol., 155: 746-754.
Direct Link - Mashhady, A.S. and D.L. Rowell, 1978. Soil alkalinity: I. Equilibria and alkalinity development. J. Soil Sci., 29: 65-75.
CrossRef - Mittal, R. and R.S. Dubey, 1991. Behaviour of peroxidases in rice: Changes in enzyme activity and isoforms in relation to salt tolerance. Plant Physiol. Biochem., 29: 31-40.
Direct Link - Munns, R., R.A. Hare, R.A. James and G.J. Rebetzke, 2000. Genetic variation for salt tolerance of durum wheat. Aust. J. Agric. Res., 51: 69-74.
Direct Link - Niu, X., R.A. Bressan, P.M. Hasegawa and J.M. Pardo, 1995. Ion homeostasis in NaCl stress environments. Plant Physiol., 109: 735-742.
PubMedDirect Link - Pandey, S.N., B.D. Nautiyal and C.P. Sharma, 2008. Pollution level in distillery and its phytotoxic e_ect on seed germination and early growth of maize and rice. J. Environ. Biol., 29: 267-270.
PubMed - Patra, D.D., M. Anwar, Tajuddin and D.V. Singh, 1998. Growth of aromatic grasses in sodic and normal soil under sub tropical conditions. J. Herbs Spices Med. Plants, 5: 11-20.
CrossRef - Patra, D.D., A. Prasad, M. Anwar, D. Singh and S. Chand et al., 2002. Performance of lemongrass cultivars intercropped with chamomile under sodic soils with different levels of gypsum application. Commun. Soil Sci. Plant Anal., 33: 1707-1721.
Direct Link - Prasad, A., A. Chattopadhyay and D.V. Singh, 2003. Growth and cation accumulation of mint genotypes in response to soil sodicity. Commun. Soil Sci. Plant Anal., 34: 2683-2697.
CrossRef - Prasad, A., A. Chattopadhyay, S. Chand, A.A. Naqvi and A. Yadav, 2006. Effect of soil sodicity on growth, yield. Essential oil composition and cation accumulation in rose scented geranium. Commun. Soil Sci. Plant Anal., 37: 1805-1817.
CrossRef - Qadar, A., 1995. Potassium and sodium contents of shoot and laminac of rice cultivars and their sodicity tolerance. J. Plant Nutr., 18: 2281-2290.
Direct Link - Rajpar, I. and D. Wright, 2000. Effect of sowing method on survival,ion uptake and yield of wheat (Triticum aestivum L.) in sodic soils. J. Agric. Sci., 134: 369-378.
Direct Link - Schachtman, D. and W. Liu, 1999. Molecular pieces to the puzzle of the interaction between potassium and sodium uptake in plants. Trends. Plant Sci., 4: 281-287.
CrossRef - Singh, S.B. and M.V. Singh, 1990. Effectof exchangeable sodium on growth, yield and mineral composition of Barley varieties. J. Indian Soc. Soil. Sci., 38: 135-138.
Direct Link - Singh, D. and B. Singh, 1997. Influence of residual sodium carbonate of sodic water on the lemon grass (Cymbopogon cytratus). J. Indian Soc. Soil Sci., 45: 354-357.
Direct Link - Singh, M., D.S. Ruhal and N. Singh, 1993. Zinc availability in sodic soils reclaimed partially to different ESP levels. J. Indian Soc. Soil Sci., 41: 251-255.
Direct Link - Soliman, S.M. and M. Doss, 1992. Salinity and mineral nutrition effects on growth and accumulation of organic and inorganic ions in two cultivated tomato varieties. J. Plant Nutr., 15: 2789-2799.
Direct Link - Tewari, T.N. and B.B. Singh, 1991. Stress studies in lentil (Lens esculenta Moench). II. Sodicity-induced changes in chlorophyll, nitrate and nitrite reductase, nucleic acids, proline, yield and yield components in lentil. Plant Soil, 136: 225-230.
CrossRefDirect Link - Trapp, S., D. Feificova, N.F. Rasmussen and P. Bauer-Gottwein, 2008. Plant uptake of NaCl in relation to enzyme kinetics and toxic effects. Environ. Exp. Bot., 64: 1-7.
CrossRef - Tyerman, S.D. and I.M. Skerett, 1998. Root ion channels and salinity. Sci. Hortic., 78: 175-235.
CrossRef - Wei, W., P.E. Bilsborrow, P. Hooley, D.A. Fincham, E. Lombi and B.P. Forster, 2003. Salinity induced differences in growth, ion distribution and partitioning in barley between the cultivar Maythorpe and its derived mutant Golden Promise. Plant Soil, 250: 183-191.
CrossRef - Perez-Alfocea, F., M.E. Balibrea, A. Santa Cruz and M.T. Estan, 1996. Agronomical and physiological characterization of salinity tolerant in a commercial tomato hybrid. Plant Soil, 180: 251-257.
CrossRef - Greenway, H. and R. Munns, 1980. Mechanisms of salt tolerance in nonhalophytes. Annu. Rev. Plant Physiol., 31: 149-190.
CrossRefDirect Link