
Michaela Schropfer
Research Institute of Leather and Plastic Sheeting, Freiberg, Germany, Meissner Ring 1-5, 09599 Freiberg, Germany
Michael Meyer
Research Institute of Leather and Plastic Sheeting, Freiberg, Germany, Meissner Ring 1-5, 09599 Freiberg, Germany
Research Journal of Phytochemistry
Year: 2016 | Volume: 10 | Issue: 2 | Page No.: 58-66







ABSTRACT
Background: The mechanism of interaction of vegetable tanning agents with skin collagen is hardly predictable due to the variability of chemical interactions of the vast number of different substance classes. Objective: In the present study it was aimed to apply less extensive methods to classify the mechanism of the cross linking action of vegetable agents with non-soluble collagen. Methodology: Hide powder was treated with solutions of different vegetable tanning agents at different pH values. The cross linking effect was analysed with DSC measurements of denaturation temperature, determination of bound amine groups before and after acidic hydrolysis and resistance against enzymatic proteolytic attack. Results: In this study a high number of common vegetable tanning extracts was analysed and classified according to the mode of interaction in three different groups: Ionic (reversible interaction), covalent non-acid-resistant and covalent acid-resistant interaction. Moreover, the tanning action of new plant extracts of common wide spread plants like privet leaf could be identified. Conclusion: The right combination of a limited number of analytical methods is sufficient to characterise the reaction mechanism of tanning agents with collagen material.
PDF Abstract XML References Citation
Received: June 02, 2016;
Accepted: June 10, 2016;
Published: June 15, 2016
How to cite this article
Michaela Schropfer and Michael Meyer, 2016. Investigations Towards the Binding Mechanisms of Vegetable Tanning Agents to Collagen. Research Journal of Phytochemistry, 10: 58-66.
URL: https://scialert.net/abstract/?doi=rjphyto.2016.58.66
URL: https://scialert.net/abstract/?doi=rjphyto.2016.58.66
INTRODUCTION
The interaction between plant polyphenols and proteins are widely investigated over the last decades and important for different fields such as leather production, beverage industry, human and animal health and in medicine. In particular, the interaction of soluble globular proteins or model peptides with tannins is of great interest for the food and nutrition industry. The role of the phenolic tanning agents is rather a disturbing one in this context. However, the methods that have been used to analyse the interaction are not transferable to insoluble proteins like collagen in hide1.
The mechanism of the protein-tannin binding is mainly proposed as hydrogen bonding or/and hydrophobic bonding. Little attention has been paid to the study of covalent bonds between proteins and tannins2-4.
The intention of tanners or manufacturers of collagen or other biopolymer-based medical products is the application of polyphenolic tannins as cross linking agents to stabilise the biopolymer structure. So far, the knowledge of the existence of covalent bonding between hide collagen and oxidised condensed vegetable tannins such as wattle or quebracho found its way in the latest text book for tanners5. But there are examples of recent publications which ignore the existence of different reaction mechanisms for different classes of vegetable tannins6.
Tannins of vegetable origin can be divided into three classes according to their chemical structure: Hydrolysable tannins, condensed tannins and tannins with iridoid or secoiridoid structure. Hydrolysable tannins are esters of gallic acid and glucose moieties (Fig. 1a). Condensed tannins are polymerised flavonoids, mostly based on catechine monomers (Fig. 1b)7,8. Iridoids belong to the substance class of terpenes and consist of a cyclopentanpyrane molecule with two oxygen atoms (Fig. 1c). The oxidised form is called secoiridoid which usually appears in a stabilised form as glycoside (Fig. 1d)9. A typical representative of hydrolysable tannins is tara, condensed tannins are the major components in wattle and secoiridoids can be found e.g., in olive leaves.
It has been suggested that hydrolysable and condensed tannins form hydrogen bonds between the polyphenolic-OH groups and the peptide oxygen of the collagen molecule10,11 or between the protonated amino-groups and the oxygen atom of the phenolic-OH groups12. The flavonoid macromolecules of condensed tannins can additionally be oxidised in the presence of the plants-own polyphenol oxidases. The resulting quinones react covalently via a nucleophilic Michael addition with amino-groups of the collagen3,13. Iridoids and secoiridoids undergo a ring opening after enzymatic deglycosilation, creating two aldehyde groups that can interact with alkaline aminoacids of collagen14. The use of iridoids and secoiridoids as cross-linking material for collagen is relatively new.
The interactions between soluble proteins and polyphenolic substances from plants can be characterised by precipitation of soluble proteins, by calorimetric determination of binding constants15, light scattering16, SDS polyacrylamide electrophoresis14, Differential Scanning Calorimetry (DSC) of denaturation6,17 or turbidity18 as examples. An overview can be found elsewhere1. A comparison of the interaction of all the three groups of vegetable tannins investigated with identical methods and under identical experimental conditions, as it is described here, could not be found. Therefore, we characterised the effects of vegetable tannins to insoluble collagen by determination of free amino-groups after derivatisation with trinitrobenzensulfonic acid (TNBS)19, measurement of denaturation temperature with DSC17 and resistance against enzymatic susceptibility17 have been applied.
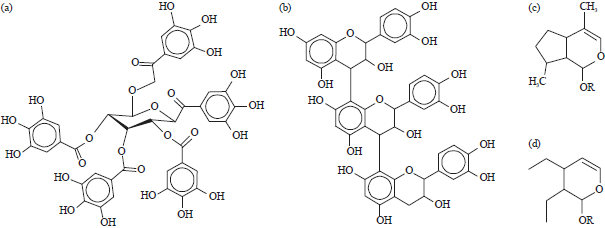 | |
| Fig. 1(a-d): | Structure of (a) Hydrolysable tannins, (b) Condensed tannins and (c-d) Secoiridoids before and after ring opening due to deglycosilation |
It is aimed to obtain new detailed insights into the interactions between vegetable tannins and collagen by combining different methods and conditions. Thereby, we were able to clearly distinguish between these different interaction mechanisms. The methodology provides simple and fast a tool box to characterise unknown plant extracts regarding their effect on insoluble proteins. As an example, a plant extract from privet leaves was characterised regarding its cross-linking potential of collagen and the reaction mechanism could be clearly identified.
MATERIALS AND METHODS
Hide powder: Hide powder was prepared from limed, air-dried cattle pelts which were milled on a cutting mill (Retsch, SM 2000) to a particle size of approximately 2 mm. It was ensured that no denaturation occurred during milling.
Plant materials: For the trials commercially available extractsof tara, valonea and myrobalane (SCRD, France), quebracho (Silvateam, Italy) and wattle (Seta sun, Brazil) were used. Privet leaves (Ligustrum vulgare), olive leaves (Olea europea), tormentil (Potentilla erecta), rhubarb radices (Rheum rhabarbarum) and oak bark (Quercus specie) were extracted in water at 50°C. The extracts were concentrated in a rotary evaporator and finally freeze dried. Glutaraldehyde (25%) was purchased from Carl Roth, Germany.
Preparation of cross-linked hide powder at different pH: Hide powder was initially soaked in a defined volume of 0.4 M McIlvain buffer at different pH in the range of pH 2-9. The plant extract was solved in water and added to the buffered hide powder in a volume ratio of 1:1 (final concentration 5% volume). Samples were shaken for 6 h at 30°C, than centrifuged. The supernatant was discarded and the samples were either washed 3 times with excess of water and finally soaked in phosphate buffer at pH 7 or remained unwashed, to evaluate the reversibility of any effects. Cross-linking with glutaraldehyde as reference was done at 1% final concentration of pure glutaraldehyde.
Differential Scanning Calorimetry (DSC) measurement: The DSC measurements were performed with a DSC 1 device (Mettler-Toledo). Approximately 6 mg (calculated on dry weight) of wet cross-linked hide powder, either unwashed (pH corresponds to the pH during cross-linking) or washed and soaked in 0.1 M phosphate buffer (pH 7) were placed in an aluminum pan and hermetically closed. Temperature scans were run from 10-125°C with a rate of 5 K min–1. From the endotherms Tonset and Tpeak were calculated. The results for the different pH are expressed as difference (ΔT) between the denaturation temperature (Tpeak) of a cross-linked sample and a sample without any crosslinking.
Enzymatic proteolysis: Cross-linked hide powder was washed in an excess of water to remove remaining cross-linker. The samples were dried and 25 mg were soaked in 1 mL 0.2 M ammonium hydrogen carbonate buffer at pH 8. Samples were denatured by heating at 85°C for 5 min and cooled down to room temperature. A 60 U of the proteolytic enzyme α-chymotrypsin from bovine pancreas E.C. 3.4.21.1 (Sigma-Aldrich) solved in 300 μL water were added and the samples were incubated at 37°C for 5 h. The enzymatic catalysed degradation was stopped with an inhibitor cocktail (Sigma-Aldrich). The supernatant was discarded and the samples were washed twice and dried. The remaining solid samples were reweighed and the difference to the initial mass was calculated as proportion of cross-linked sample in relation to non-cross-linked sample.
Determination of bound ε-amino groups before acid hydrolysis: Five milligrams of sample were incubated in 200 mL 0.5 M ammonium hydrogen carbonate buffer pH 8 200 mL of 0.5% trinitrobenzenesulfone acid (TNBS) were added20. The samples were incubated at 40°C for 2 h. The TNBS binds to free ε-amino-groups. After this derivatisation step the samples were hydrolysed with 6 N HCl at 60°C for 6 h, diluted with water and absorption was measured photometrically at 400 nm. Quantification was done by calibration with an alanine standard. The amount of ε-amino-groups bound by the cross-linking agent was calculated based on the number of free ε-amino-groups of cross-linked and non-cross-linked samples.
Determination of bound amino groups after acid hydrolysis: The samples were hydrolysed with 6 N HCl at 110°C for 20 h, dried and resolved in lithiumcitrate buffer. Amino acid composition was determined from cross-linked and non-cross-linked samples by an amino acid analysator (Biochrom 30+) with pre-column derivatisation with ninhydrin according to standard protocols.
The percentage of amino-groups, that formed an acid-stable bond was calculated from the area under the lysine, hydroxylysine or arginine peaks and normalised to the area under the peaks from alanine and valine (not involved in cross-linking). The resulting factor was related to the same factor calculated from non-cross-linked sample. All analytical results are averages of a duplicate.
RESULTS AND DISCUSSION
To characterise the mechanism and the degree of cross linking, different analytical methods were applied to the treated hide powder.
Tara (hydrolysable tannin), wattle (condensed tannin) and olive leaves (secoiridoids) were chosen as examples to explain the cross linking activity of the three different chemical classesof tannins (Fig. 2-5).
Figure 2 shows the denaturation temperatures measured by DSC. This temperature correlates strongly with the cross-linking degree. Depending on the washing step two distinct curves appeared. For example, the denaturation temperature difference ΔT of unwashed samples of hide powder tanned with tara decreases with increasing pH indicating that hydrolysable tannins from tara cross-link only in the acidic pH range. Transferring the sample into pH 7 by washing nearly removes all cross-links. With respect to the protonation and deprotonation of acidic and alkaline polar groups of collagen tara preferentially interacts with protonated amino-groups.
In contrast, the cross-linking effect for unwashed samples treated with wattle decreases slightly with increasing pH value but remains stable at +25 K in neutral and alkaline range. The washed samples show a slight increase in cross linking with increasing pH value suggesting a preferential binding at unprotonated basic amino-groups typically seen for nucleophilic addition reactions. Few cross-links are formed also in the acidic range at protonated amino-groups.
The olive leaf extract-tanned hide powder shows a similar behaviour but the slope of increase in denaturation temperature with rising pH values is more pronounced. In the acidic range no cross-linking is observed. This is again typical fora nucleophilic addition at unprotonated amino-groups.
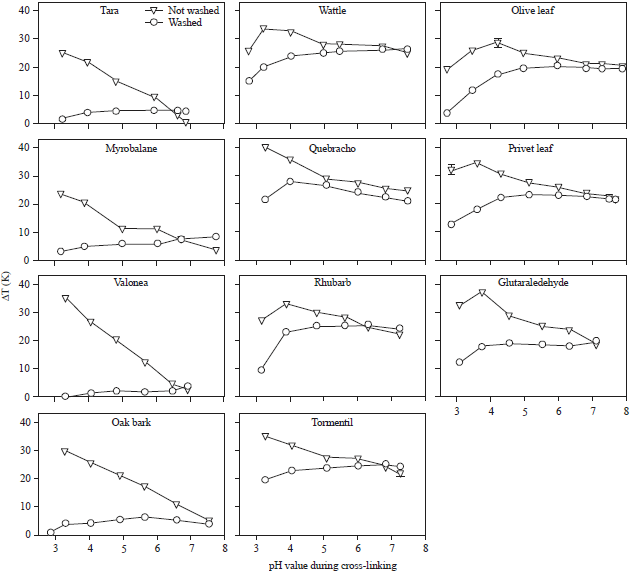 | |
| Fig. 2: | pH dependent change of denaturation temperature of hide powder cross-linked with vegetable tanning extracts relative to non-cross linked control. Samples were either washed and soaked in pH 7 or remained unwashed after cross-linking |
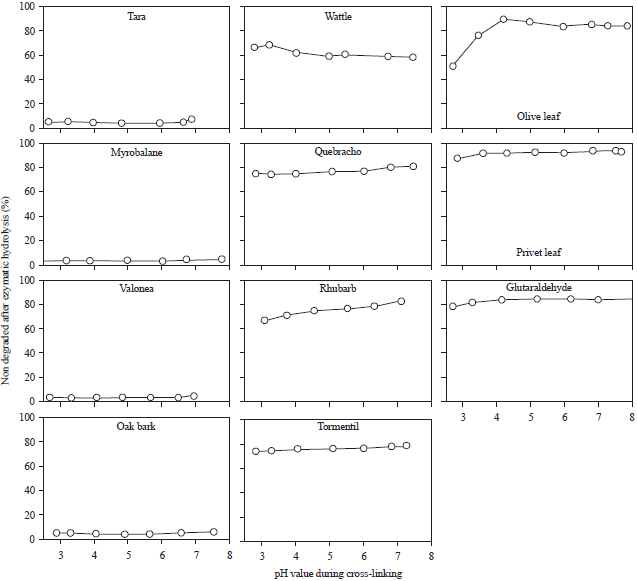 | |
| Fig. 3: | Percentage of non-degraded/insoluble cross-linked hide powder after enzymatic attack by α-chymotrypsin. Enzymatic degradation was performed at pH 8 |
To further characterise the strength of cross-linking, the samples were enzymatically degraded. Figure 3 shows the resistance of tanned hide powder against proteolytic attack by α-chymotrypsin with a maximum enzymatic activity at pH 8. Tara-tanned hide powder showed no resistance against proteolysis, because itis completely detanned at pH 8. The wattle-tanned hide powder was only partly degraded due to the strong tanning effect in the acidic range as well as under alkaline conditions. The olive leaf-tanned hide powder showed elevated resistance with increasing pH value. Samples cross-linked at pH 4 and higher were stable against enzymatic proteolysis.
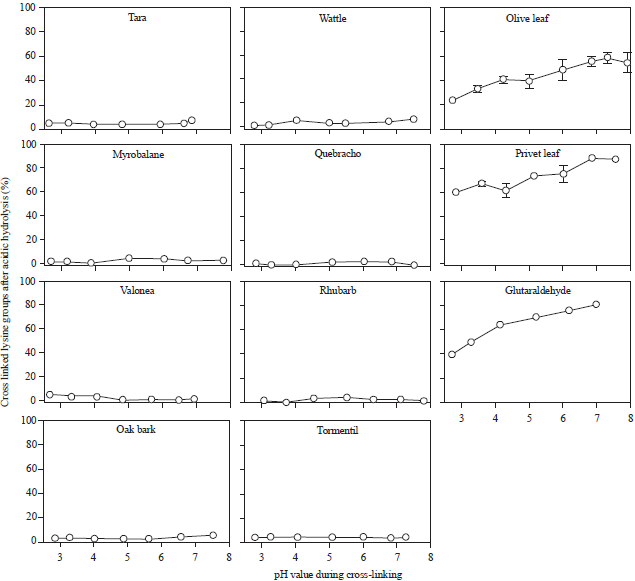
To quantify the number of ε-amino-groups (i.e., lysine side chains), that are involved in the cross-links, the treated and untreated samples were derivatised with TNBS. Figure 4 shows the percentage of bound ε-amino-groups.
Tara contains only a few cross-linked amino-groups indicating that a few ε-amino-groups could be blocked in the acidic range may be by electrostatic interactions. In contrast, wattl ε-tanned hide powder revealed a high number of covalently bound ε-amino-groups. Surprisingly even more bound groups have been found in the acidic range. An increasing number of bound ε-amino-groups with pH values higher than pH 6 indicates again that in olive-tanned hide powder deprotonated groups are preferred.
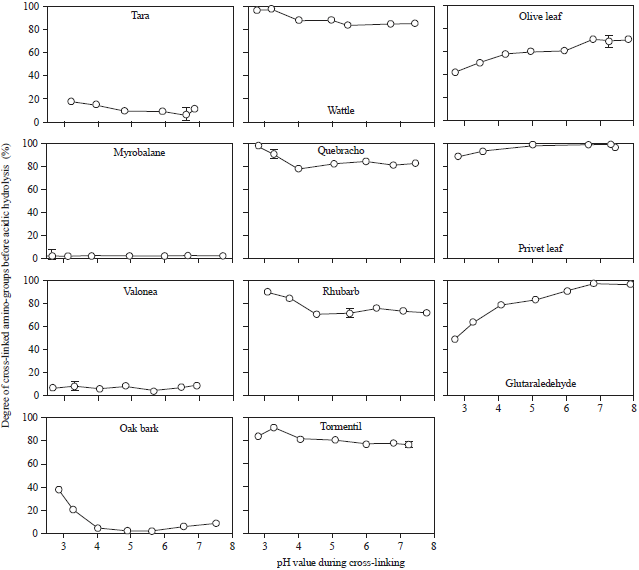
To differentiate between acid-stable and non acid-stable cross-links the samples were investigated by amino acid analysis. The samples were treated with strong acid which leads to breaking of peptide bonds and acid-soluble cross-links, resulting in single amino acids. Acid-stable cross-links can be detected by a decrease in the amount of amino acids that are involved in cross-linking (Fig. 5).
 | |
| Fig. 4: | Percentage of bound ε-amino-groups before acid hydrolysis determined with the TNBS method |
The high amount of bound amino-groups, which could be detected by the TNBS method for wattle-tanned hide powder, disappeared. Wattle tannin induced cross-links are not acid-stable. Only the bonds of hide powder tanned with olive leave extract with stood the strong acidic hydrolysis.
Considering the results and the published data from the literature the different tannin groups can be assigned to the following reaction schemes.
Hydrolysable tannins: Hydrolysable tannins create interactions at protonated amino-groups, which completely disappear with increasing pH values.
If hydrogen bonds play a role, they can either exist between peptide oxygen atoms of the collagen and the H-atoms of hydroxyl groups of the polyphenolic tannin11 or between protonated ε-amino-groups of the collagen and the oxygen atoms of the hydroxyl groups of the polyphenolic tannin12. The preference for protonated amino-groups during cross-linking can also be interpreted by electrostatic interactions.
Condensed tannins: The participation of unprotonated amino-groups in cross-linking indicates that covalent bonds are formed between ε-amino-groups of collagen and quinones, which emerge by enzymatically catalysed oxidation of certain polyphenols. The respective enzymes (polyphenol oxidases) are plants-own enzymes. The quinones can react with collagen via a nucleophilic 1,4-Michael addition creating covalent bonds13. The cross-links are obviously not stable against acid hydrolysis.
Iridoids/secoiridoids: A well-known representative of the secoiridoidsis oleuropein. This and structurally similar compounds are found in olive leaves. It has been proposed that deglycosilation results in ring opening and the formation of two aldehyde groups, which then can cross-link collagen via Schiff-base formation14.
 | |
| Fig. 5: | Percentage of bound ε-amino-groups (lysine) after acid hydrolysis determined with amino acidanalysis |
The Schiff-bases are not stable by themselves but are stabilised by further rearrangements. Oleuropein has additionally two phenolic hydroxyl groups, which can be oxidised by polyphenol oxidases into quinones and react to other covalent bonds14. The tanning effect increases with increasing pH, because the first step, the Schiff-base formation is preferred at unprotonated amino-groups of collagen which is only the case in neutral or basic pH-range. The resulting covalent cross-links are stable against acidic hydrolysis. In summary, the pH dependent cross-linking activity of olive leaves appears as a pattern that is analogous to the pattern of glutaraldehyde-tanned hide powder14.
With this experimental setup a number of vegetable tanning agents (Tara, oak bark, valonea and myrobalane as hydrolysable tannins; wattle, quebracho, tormentil and rhubarb as condensed tannins and olive leaves as secoiridoid) containing plants were investigated. Glutaraldehyde with known reaction mechanism was analyzed as a reference. Each tannin fits into one of the three reaction patterns (Fig. 2-5) described. The classification is consistent with the classification of vegetable tannins based on their chemical structure (Fig. 1).
The extract of privet leaves were not used as tanning agents so far. As the olive tree privet is a member of the plant family of the Oleacea and their leaves contain e.g., oleuropein and structural similar compounds like ligustrosid and ligustalosid A and B21,22, but also a lot of other iridoids, secoiridoids and flavonoids21-23. In order to classify its reaction mechanism with collagen, it was investigated in the same way.
The tannins of privet leaves obviously belong to the third group. They cross-link covalently and the cross-links are acid stable. The tanning effect seems to be very strong compared to olive leaves, which were applied in the same concentration.
The methods of the investigations of polyphenol-protein-interactions with soluble proteins cannot be transferred to insoluble protein-tannin interactions. Especially in the field of leather making the investigation of interactions between hides and vegetable tannins cover a period of about 100 years. In the past there was a controversy whether covalent cross-links play a role in the interaction of vegetable polyphenolic tannins with collagen24,25. Currently, there are only scarce studies about the exact chemical interactions between insoluble collagen and polyphenols. Some recent studies could be found, which deals rather with kinetic questions of diffusion behavior26 of the large polyphenolic molecules and the binding site within the hierarchical collagen structure27.
Regarding the interaction of iridoids with insoluble hide collagen, Ding et al.28 investigated the reaction of genipin with hide powder and also the pH-dependence of the cross-linking effects. The results are in line with our observations. However, the researchers did not determine the amount of bound amino-groups. Therefore, only little insights have been gained into the cross-linking mechanisms.
From the pH dependence and the comparison of bound amino-groups before and after acidic hydrolysis new insights into the different reaction mechanisms of chemically different vegetable tannins could be generated: Reversible non-covalent effects appearing with hydrolysable tannins, covalent bonds, which are not stable against acid hydrolysis with condensed polyphenols and acid stable covalent bonds with iridoids and secoiridoids.
The cross-linking activity of oleuropein from privet leaf is well described14 and a reaction mechanism has been suggested. However, the investigation was done with bovine serum albumin, a soluble globular protein and the study aimed to find out the action of plant own enzymes for activation of the secoiridoide. The ability of privet leaf extract as excellent cross-linker for hide collagen had not yet been described.
CONCLUSION AND FUTURE RECOMMENDATION
By combining the use of hide powder as collageneous matrix with vegetable extracts in cross-linking trails we were able to identify tanning agents, that form stable, covalent bonds with non-soluble proteins. Moreover, the pH dependent setup und well selected analytical methods allowed to identify the reaction mechanisms of cross-linking. All plant extracts investigated here could be classified into three groups of interaction with a characteristic pattern of analytical results. The third group showed cross-linking properties that are different from those of the hydrolysable and condensed tannins. These new tannins belong to the chemical group of iridoids or secoiridoids. In contrast to condensed tannins they induce covalent cross-links, which are stable against acid hydrolysis.
The methods applied here can be used to characterise the cross-linking effect of unknown substances for example during the development of new synthetic retanning agents or new vegetable crosslinkers for food and pharmaceutical applications. Additionally, the knowledge about the influence of pH values during cross-linking is important for the optimisation of the diffusion and binding of tannins during the tanning process.
REFERENCES
- Beart, J.E., T.H. Lilley and E. Haslam, 1985. Polyphenol interactions. Part 2. Covalent binding of procyanidins to proteins during acid-catalysed decomposition; observations on some polymeric proanthocyanidins. J. Chem. Soc. Perkin Trans., 2: 1439-1443.
CrossRefDirect Link - Peter, M.G., 1989. Chemical modifications of biopolymers by quinones and quinone methides. Angewandte Chemie Int. Edn., 28: 555-570.
CrossRefDirect Link - Kroll, J., H.M. Rawel and S. Rohn, 2003. Reactions of plant phenolics with food proteins and enzymes under special consideration of covalent bonds. Food Sci. Technol. Res., 9: 205-218.
CrossRefDirect Link - Brown, E.M., R.J. Latona and M.M. Taylor, 2015. Powdered hide model for vegetable tanning II: Hydrolyzable Tannin. J. Am. Leather Chem. Assoc., 110: 19-22.
Direct Link - Hansel, R. and O. Sticher, 2010. Pharmakognosie-Phytopharmazie. Springer, Berlin, Germany, ISBN: 978-3-642-00962-4, Pages: 1451.
Direct Link - Mavlyanov, M., S.Y. Islambekov, A.I. Ismailov, D.N. Dalimov and N.G. Abdulladzhanova, 2001. Vegetable tanning agents. Chem. Nat. Compd., 37: 1-24.
CrossRefDirect Link - Spencer, C.M., Y. Cai, R. Martin, S.H. Gaffney and P.N. Goulding et al., 1988. Polyphenol complexation-some thoughts and observations. Phytochemistry, 27: 2397-2409.
CrossRefDirect Link - Endres, H., 1964. Theory of interaction of vegetable tannins with collagen. Leather Sci., 11: 455-462.
Direct Link - Konno, K., C. Hirayama, H. Yasui and M. Nakamura, 1999. Enzymatic activation of oleuropein: A protein crosslinker used as a chemical defense in the privet tree. Proc. Natl. Acad. Sci., 96: 9159-9164.
CrossRefDirect Link - McManus, J.P., K.G. Davis, J.E. Beart, S.H. Gaffney, T.H. Lilley and E. Haslam, 1985. Polyphenol interactions. Part 1. Introduction: Some observations on the reversible complexation of polyphenols with proteins and polysaccharides. J. Chem. Soc. Perkin Trans., 2: 1429-1438.
CrossRefDirect Link - Siebert, K.J., N.V. Troukhanova and P.Y. Lynn, 1996. Nature of polyphenol-protein interactions. J. Agric. Food Chem., 44: 80-85.
CrossRefDirect Link - Brown, E.M., R.J. Latona, M.M. Taylor and A.G. Gehring, 2014. Powdered hide model for vegetable tanning. J. Am. Leather Chem. Assoc., 109: 8-13.
Direct Link - Siebert, K.J., 2006. Haze formation in beverages. LWT-Food Sci. Technol., 39: 987-994.
CrossRefDirect Link - Rawel, H.M., D. Czajka, S. Rohn and J. Kroll, 2002. Interactions of different phenolic acids and flavonoids with soy proteins. Int. J. Biol. Macromol., 30: 137-150.
CrossRefDirect Link - Habeeb, A.F.S.A., 1966. Determination of free amino groups in proteins by trinitrobenzenesulfonic acid. Anal. Biochem., 14: 328-336.
CrossRefDirect Link - Romani, A., P. Pinelli, N. Mulinacci, F.F. Vincieri, E. Gravano and M. Tattini, 2000. HPLC analysis of flavonoids and secoiridoids in leaves of Ligustrum vulgare L. (Oleaceae). J. Agric. Food Chem., 48: 4091-4096.
CrossRefDirect Link - Tattini, M., C. Galardi, P. Pinelli, R. Massai, D. Remorini and G. Agati, 2004. Differential accumulation of flavonoids and hydroxycinnamates in leaves of Ligustrum vulgare under excess light and drought stress. New Phytol., 163: 547-561.
CrossRefDirect Link - Diak, J., 1988. Phytochemical study of privet (Ligustrum vulgare L.). Herba Polonica, 34: 103-108.
Direct Link - Covington, A.D., T.H. Lilley, L. Song and C.S. Evans, 2005. Collagen and polyphenols: New relationships and new outcomes. Part 1. Flavonoid reactions for new tanning processes. J. Am. Leather Chem. Assoc., 100: 325-335.
Direct Link - Ding, K., M.M. Taylor and E. Brown, 2006. Effect of genipin on the thermal stability of hide powder. J. Am. Leather Chem. Assoc., 101: 362-367.
Direct Link