
James A. Timbilla
Queensborough Community College, City University of New York, 222-05 56th Avenue, Bayside, 11364,
New York, USA
K. Yeboah-Gyan
Department of Theoretical and Applied Biology, Kwame Nkrumah University of Science and Technology, Kumasi, Ghana
B.W.L. Lawson
Department of Theoretical and Applied Biology, Kwame Nkrumah University of Science and Technology, Kumasi, Ghana
T. Hartmann
Institute of Pharmaceutical Biology, Technical University of Braunschweig, D-38106, Braunschweig, Germany
Research Journal of Phytochemistry
Year: 2015 | Volume: 9 | Issue: 1 | Page No.: 1-15







ABSTRACT
Since the 1970s, the pestilence of dry season populations of the polyphagous grasshopper Zonocerus variegatus (Orthoptera: Pyrgomorphidae) has impacted negatively on agriculture and forestry in Sub-Saharan Africa. Current chemoecological knowledge indicates that outbreaks of Z. variegatus may be due to sequestration of protective Pyrrolizidine Alkaloids (PAs) from the introduced perennial shrub, Chromolaena odorata (Asteraceae; Eupatorieae). A better knowledge of the impact of the plant PA system on the performance of Z. variegatus should help to develop efficient strategies for controlling outbreaks of Z. variegatus. The influence of the accessibility of a PA source (dried roots of C. odorata) on the development of Z. variegatus reared on cassava foliage as sole nutrient source was studied. The comparative studies, in which females and males were analyzed in separate trials, revealed that in the presence of the PA source Z. variegatus showed: (1) A slightly higher survival rate, (2) Significantly increased body weight and size (increased length of the thorax shield) and (3) Females produced heavier egg pods with more but lighter eggs. Eggs produced in the presence of the accessible PA source were found to contain PAs in concentrations known to efficiently protect eggs of PA sequestering Lepidoptera (e.g., arctiid moths) against various predators and parasitoids. Participation of a female-to-male-transfer of PAs could be excluded. The results suggest that in Z. variegatus, as in PA adapted leaf beetles and Lepidoptera, the acquisition of the plant PA system and its integration into the insect’s biology is associated with specific functional and behavioral adaptations.
PDF Abstract XML References Citation
Received: January 13, 2015;
Accepted: February 28, 2015;
Published: July 10, 2015
How to cite this article
James A. Timbilla, K. Yeboah-Gyan, B.W.L. Lawson and T. Hartmann, 2015. A Grasshopper and its Beneficial Drug Source: The African Story of Zonocerus variegatus and the Neophyte Chromolaena odorata. Research Journal of Phytochemistry, 9: 1-15.
URL: https://scialert.net/abstract/?doi=rjphyto.2015.1.15
URL: https://scialert.net/abstract/?doi=rjphyto.2015.1.15
INTRODUCTION
In the dry season Zonocerus variegatus (Orthoptera: Pyrgomorphidae) is a major grasshopper pest of crops in the forest and savannah areas of West and Central Africa (Toye, 1982; Boppre and Fischer, 1994; Fischer and Boppre, 1997). During the last decades outbreaks of Z. variegatus have been repeatedly observed in areas with high abundance of the introduced Siam weed (Chromolaena odorata) (Asteraceae, tribe Eupatorieae) and the suggestion has been made that the spread of this weed could be a factor boosting the grasshopper outbreaks (Toye, 1974; Chapman et al., 1986; Popov, 1988). Chromolaena odorata is a perennial shrub native to South and Central America. During the last century it established as a serious pest in the humid tropics of South East Asia, Africa and Pacific Islands. It reached Africa in the late 1930s and became a pest weed in the 1970s (Bennet and Rao, 1965; Modder, 1984; Muniappan and Marutani, 1988; Fischer and Boppre, 1997). The plant can form dense, extensive thickets that have been described as suitable but not exceptional aggregation and oviposition sites for Z. variegatus (Chapman et al., 1986; Modder, 1997). In fact, all life stages of Z. variegatus are attracted olfactorily to the open inflorescences of C. odorata (Modder, 1984, 1986). This attraction is correlated with a strong feeding stimulation causing Z. variegatus to feed actively on the flower heads. Subsequently it was found that the attractive principle is related to Pyrrolizidine Alkaloids (PAs) present in C. odorata and that various other PA-containing plants as well as the pure alkaloids are lures for all life stages of Z. variegatus (Boppre et al., 1984; Boppre, 1991). Obviously dry season populations of Z. variegatus benefit from the local all-dominance of C. odorata thickets as PA source when it is in bloom. This intriguing plant-insect relationship is apparently the result of a ‘happy meeting’ (Modder, 1996) of two pest organisms in the forest clearings of West Africa. Probably not the abundance of C. odorata itself but its role as an accessible and inexhaustible source of PAs makes the benefit for Z. variegatus.
Pyrrolizidine alkaloids are ester alkaloids produced constitutively by plants as chemical defense against herbivores, particularly insects. The occurrence of more than 400 structures is restricted to a few unrelated angiosperm taxa. About 95% of the species containing PAs belong to only five families, viz., the Asteraceae (i.e., Senecioneae and Eupatorieae), the Boraginaceae, the Apocynaceae, the genus Crotalaria within the Fabaceae and Orchidaceae (Hartmann and Witte, 1995). The PAs are strong feeding deterrents for most herbivores and part of fascinating interactions between PA producing plants and adapted insects that during evolution recruited the plant defense system and utilize it for their own benefit (Boppre, 1986; Schneider, 1987; Hartmann, 1999, 2004). Well studied examples are certain leaf beetles (Pasteels and Hartmann, 2004) and among the Lepidoptera, a number of arctiid moths as well as danaine and ithomiine butterflies (Eisner et al., 2002). The acquisition of the plant PA system and its integration into insect-specific defenses is associated with highly specific biochemical, physiological and behavioral adaptations.
A better mechanistic understanding of the relationship between Z. variegatus and its plant PA sources should promote strategies to control outbreaks of this pest insect (Boppre and Fischer, 1994; Fischer and Boppre, 1997). In this study it was examined that the effect of a PA source on growth and development of Z. variegatus. Zonocerus variegatus is highly polyphagous but unlike many food generalists, only a few plants, such as cassava (Manihot esculenta) and citrus guarantee optimal growth, development and egg production, when offered as sole diet (Bernays et al., 1975; Modder and Singh, 1976; McCaffery et al., 1978). Zonocerus variegatus has the peculiarity of regularly changing its food plant due to the requirement of mixed diet for proper development (Vuillaume, 1953; Modder, 1984, 1986; Modder and Tamu, 1996). The present feeding strategy was rearing Z. variegatus on: (1) Fresh cassava foliage as sole diet which allowed excellent performance and egg production, (2) Identical conditions but free access of the insects to a PA source (i.e., dry chopped roots of C. odorata). In comparative trials the following aspects were studied and discussed: (1) Influence of PA accessibility on: (1) Survival rate, (2) Body weight and length of thoracic shield, (3) Effect of PA sequestration on various characteristics of egg production. In addition the transmission of PAs to eggs directly from females and possibly, from males via females was investigated.
MATERIALS AND METHODS
Origin and rearing of insects: Collections and maintenance of newly hatched nymphs of Z. variegatus (resulting from the same batch of eggs at a particular oviposition site) were made in fallow fields in Cape Coast and Kumasi in August 2001 according to the procedure developed by Cobbinah and Tuani (1992). Each cohort of Z. variegatus was fed with freshly cut cassava foliage maintained in earthen pots filled with sand and watered daily. All the rearing was done at the Crops Research Institute (CRI), Kumasi under ambient conditions.
The insectary measured 8x5 m with 8 cages each measuring 2×2×3 m, four on either side with a meter walkway in-between. The floor of each cage was cemented and had block walls of 1 m height with the remaining 2 m covered with metal mesh and roofed with translucent fiber glass sheets. The cages were thoroughly cleaned every other day and provided with fresh supply of feed. River bed sand was sterilized at 250°C for 30 min and spread to the dimensions 0.5×0.5×0.2 m on the floor at one corner of each cage and moistened with water for oviposition when the insects turned adults.
Feeding experiments: Three comparative feeding experiments were conducted. All trials were started with third instar larvae reared on cassava as sole food. The treatments, each trial with 40 specimens of males and females, were as follows: (1) Larvae fed on cassava as sole food source and (2) Larvae fed on cassava as sole food source and given continuous access to C. odorata roots as PA source. Both sexes were reared together but data collection was done separately for each sex. Roots of C. odorata were chosen because they are a rich source of PAs (Biller et al., 1994) and available throughout the year. They were chopped into 10 cm pieces that were offered to the insects (Fig. 1). The insects generally only nibbled at the root cortex to get access to the PAs. Cassava leaves and C. odorata roots were replaced by fresh materials every two days. The first experiment was conducted with an insect culture from Nsanfo, near Cape Coast, while the second and third experiments were conducted with insects collected from fallow fields near Kwadaso, Kumasi.
Parameters measured and data analysis
Survival rate: For experiments 1 and 2 mortality was counted at weekly intervals till death of the last specimen (16-22 weeks).
Weight: For experiments 1 and 3 the body weight (fresh weight) was determined 30 days after onset of the experiment. All the weight measurements were done after starving the insects for 24 h (Wigglesworth, 1972).
 | |
| Fig. 1: | Dry roots of Chromolaena odorata showing nibbling by adult Zonocerus variegatus |
Lengths of thorax shield (from end pro-thorax to end meta-thorax) were determined by means of a Venier caliper immediately after the specimens of the respective trial turned adult.
Oviposition: Egg pods were collected from the female populations of each treatment and counted. Additionally, egg pods of experiment 3 were weighed and the fresh and dry weights of resulting egg masses measured. Numbers of eggs per pod were counted and the egg masses weighed. In experiment 3, data was limited to length of thorax and egg production.
Alkaloid analysis: Eggs and isolated egg pods were weighed and then ground in 0.2-2 mL 1 M HCl in a mortar, extracted for 2-3 h and then centrifuged. The supernatant was saved and the pellet dissolved in a small volume of HCl and again extracted. The combined supernatants were extracted with 2 mL dichloromethane, the aqueous phase was recovered, mixed with excess of Zn dust and stirred for 3 h at room temperature for complete reduction of the PA N-oxides. Then the mixture was made basic with 25% ammonia and applied to an Extrelut (Merck) column (size adapted to 1.4 mL solution/g extrelut). The Pas (free bases) were eluted with dichloromethane (6 mL g-1 extrelut). The solvent was evaporated and the residue dissolved in 10-100 μL of methanol prior to GC or GC-MS. Routine Gas Chromatography (GC) was performed as described previously (Witte et al., 1992; Hartmann et al., 2004a). Quantitative analyses were performed via the FID signals with heliotrine as internal standard.
The GC-MS data was obtained with a Hewlett Packard 5890A gas chromatograph equipped with a 30 mx0.32 mm analytical column (ZB1, Phenomenex). The capillary column was directly coupled to a triple quadrupole mass spectrometer (TSQ 700, Finnigan). The conditions applied were: Injector and transfer line were set at 250°C; the temperature program used was: 100°C (3 min) to 310°C at 6°C/min. The injection volume was 1 μL. The split ratio was 1:20, the carrier gas flow was 1.6 mL min-1 He and the mass spectra were recorded at 70 eV.
Statistical analysis: Data comparisons on growth parameters were subjected to the t-test while the Keplan Meir survival curves were used to compare mortalities using the GraphPad Prism® Software Version 4.00 for Windows (Miller, 2003). Results were significant at p<0.05.
RESULTS
Survival rate of Z. variegatus in the presence and absence of a PA sources: The grasshoppers developed properly if reared on cassava as sole nutrient source. If pieces of dried roots of C. odorata were offered together with cassava the hoppers were periodically found nibbling at the surface of the dry roots obviously ingesting PAs from the alkaloid rich cortex tissue (Fig. 1). In the course of experiments 1 (Origin of insect culture: Cape Coast) and 2 (Origin of culture: Kumasi) mortality of the cohorts were evaluated separately for males and females reared on cassava in the presence (+PA) and absence (-PA) of C. odorata roots. The results are illustrated in Fig. 2. In both experiments the survival rates of males and females that had access to the C. odorata were somewhat higher. In experiment 1, 50% mortality was observed within 5 weeks for males and females in the -PA population but not until 10 weeks (males) and 12 weeks (females) in the +PA population. In Experiment 2 the differences were less obvious, i.e., 50% mortality of females after 13 weeks (-PA) and 16 weeks (+PA) and of males after 6 weeks (-PA) and 8 weeks (+PA). In both experiments the major differences in mortality between the +PA and the -PA populations occurred during the first few weeks. The total life span of the +PA populations were higher than of the -PA populations.
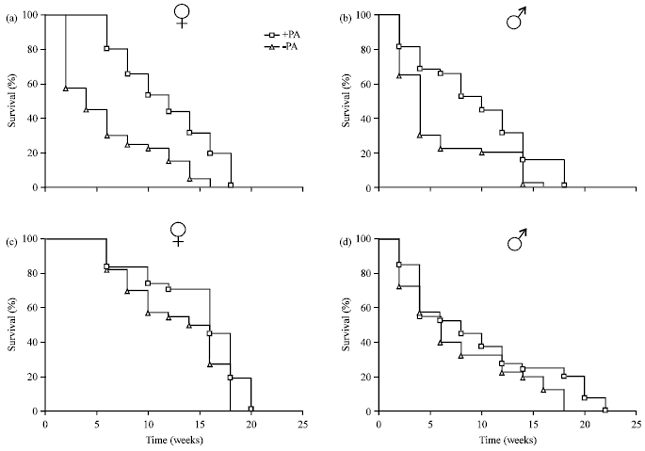 | |
| Fig. 2(a-d): | Survival of Zonocerus variegatus reared on cassava foliage as sole diet in the absence (-PA) and presence (+PA) of dry Chromolaena odorata roots. The experiment was started with 3rd instar larvae (to), (a) Experiment 1 (females), (b) Experiment 1 (males), (c) Experiment 2 (females) and (d) Experiment 2 (males), n = 40 for each trial |
Effect of a PA source on growth rate: The results obtained from experiment 1 with insects from Cape Coast indicate that Z. variegatus reared on cassava in presence (+PA) of dried C. odorata roots gained significantly more weight than those reared in its absence (-PA) (Fig. 3). Within the duration of the experiment females gained mean weights of 0.51 and 0.24 g each over their original weights, constituting 72.9% (presence of PA source) and 34.0% (absence of PA source) weight gain, respectively. A similar pattern was observed with males: Weight gain of 0.38 g (59.4%) (presence of PA source) in comparison to only 0.20 g (34.3%) (absence of PA source). With both sexes the observed weight changes were significantly higher in the PA-treated insects (p<0.05).
In experiment 2 with insects from Kumasi, the female Z. variegatus reared in the presence of the PA source also gained significantly more weight than those reared in the absence of PAs (Fig. 3). The mean weight increase was 0.51 and 0.35 g, i.e., 68.5 and 54.7%, respectively, for the two treatments. Under the same conditions the mean weight gain of males was 0.48 g (72.7%) and 0.39 g (69.6%) for the two differences. However, the differences were not significant.
Further experiments performed in the same manner as experiments 1 and 2 but containing an additional trial with cassava plus leaves of Gliricidium sepium in the presence and absence of C. odorata roots revealed no significant differences.
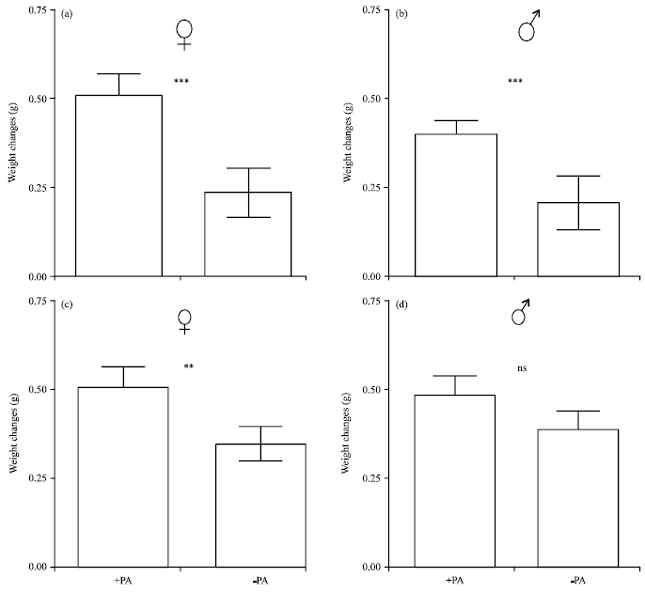 | |
| Fig. 3(a-d): | Weight gain of 3rd instar larvae of Zonocerus variegatus transferred to cassava diet with (+PA) and without (-PA) access to dried roots of Chromolaena odorata. All trials were analyzed 30 days after application of diet, (a) Experiment 1 (females), (b) Experiment 1 (males), (c) Experiment 2 (females) and (d) Experiment 2 (males), n = 8-36 for each trial, ***p<0.01, **p<0.05, ns: Not significant |
The size the thorax length determined for the individual insects included in experiments 1-3 are summarized in Table 1. In experiment 1 no significant differences were observed between insects of both sexes fed on cassava in the presence (+ PA) or absence (-PA) of C. odorata roots (Table 1). However, with both sexes slightly greater thorax lengths were measured for the insects reared in the presence of the PA source. In experiments 2 and 3, there were significant differences observed in the length of thorax for both sexes with respect to the two treatments, i.e., p<0.05 and p<0.001, respectively.
| Table 1: | Mean thoracic length of adult Zonocerus variegatus reared on cassava diet in presence (+PA) or absence (-PA) of dried Chromolaena odorata roots |
 | |
| aParenthesis sample size (n), ns: Not significant, *p<0.05, ***p<0.001 | |
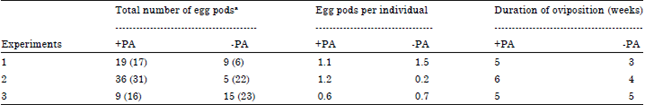
| Table 2: | Production of egg pods and duration of oviposition of female Zonocerus variegatus reared on cassava diet in the presence (+ PA) and absence (-PA) of dried Chromolaena odorata roots |
 | |
| aParenthesis sample size (n) | |
| Table 3: | Mean weight of egg pods and number of eggs per egg pod of Zonocerus variegatus reared on cassava in the presence (+PA) or absence (-PA) of Chromolaena odorata roots (data from Experiment 3) |
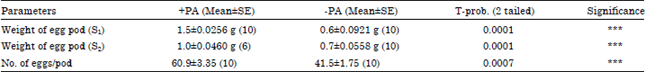 | |
| aParenthesis sample size (n), S1 and S2 designated as Sample 1 and 2, ***Significant at specified probability | |
Effect of a PA source on egg production and weight: In experiments 1 and 2, mated females reared in the presence (+PA) or absence (-PA) of C. odorata roots started ovipositing after 12 weeks while those in experiment 3 started oviposition not before 14 weeks. The total number of egg pods produced by the female populations of the two treatments and the calculated egg pods per single female within the population are shown in Table 2. The result does not show clear differences with exception of experiment 2 where the number of egg pods produced in average by +PA females (i.e., 1.2) is considerably higher than the number obtained with -PA females (i.e., 0.2). The duration of oviposition in experiments 1 and 2 is two weeks extended in the +PA females compared to the -PA females. However, no difference in the duration of oviposition was observed in experiment 3.
The results on egg pods collected at two different times in experiment 3 indicated significantly heavier egg pods in the PA treatments than the non-PA treatment (p<0.05) (Table 3). The egg pods produced by the PA-treated insects were generally bigger than those from the sole cassava treatment which were more slender (Fig. 4). Also, the mean number of eggs in each egg pod of the PA-treatment was significantly higher (60.9 eggs) than the cassava treatment of 41.5 eggs (p<0.05) (Table 3).
 | |
| Fig. 4(a-c): | Eggs pods of Zonocerus variegatus, (a) Broken eggs pod showing the eggs, (b) Egg pods resulting from +PA females and (c) Egg pods resulting from -PA females |
| Table 4: | Mean weight of fresh and dry eggs of Zonocerus variegatus reared on cassava in the presence (+PA) or absence of dried roots of Chromolaena odorata |
 | |
| an = 30 in each trial, bOnly dry weight available in Experiment 2, **p<0.05, ***p<0.001 | |
In contrast to the bigger egg pods and the higher number of eggs per egg pod produced by +PA females the opposite was observed with regard to egg weight. In all experiments tested the eggs produced by ‘-PA females’ were significantly heavier than the eggs deposited by ‘+PA females’ (Table 4). These differences apply for both fresh and dry weights.
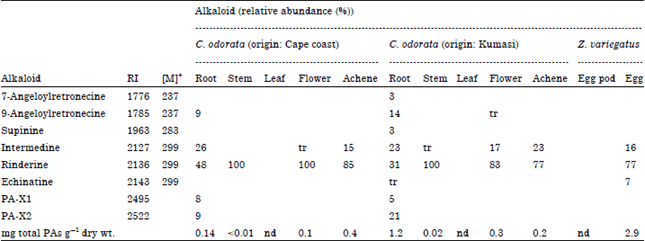
Transmission of plant acquired PAs to eggs of Z. variegatus: GC-MS analysis of two C. odorata provenances, i.e., Cape Coast and Kumasi revealed comparable PA profiles with rinderine and intermedine as major alkaloids in the roots and as exclusive PAs in the aboveground plant parts (Table 5). Roots which in species of the Eupatorieae are the sites of PA biosynthesis (Weber et al., 1999), contained a number of minor alkaloids such as 9-angeloylretronecine, supinine and two unidentified PAs (i.e., X1 and X2). Leaves were found essentially devoid of PAs. Total PA concentrations were relatively low. The highest levels with 0.12% were found in the root samples from Kumasi. The aboveground parts contained substantial amounts (ranging from 0.01-0.04%) only in the inflorescences. Both the PA profiles and total PA concentrations found in the various plant organs correspond well to previous reports on PA profiles of C. odorata collected in Bénin (Biller et al., 1994). Eggs obtained from Z. variegatus reared on cassava which had free access to flowering shoots of C. odorata were found to contain total PAs at a concentration of 2.9 mg g-1 dry wt. which is, in comparison to C. odorata inflorescences, an almost 7 to 30-fold higher level (Table 5 and Fig. 5). Egg pods were devoid of PAs.
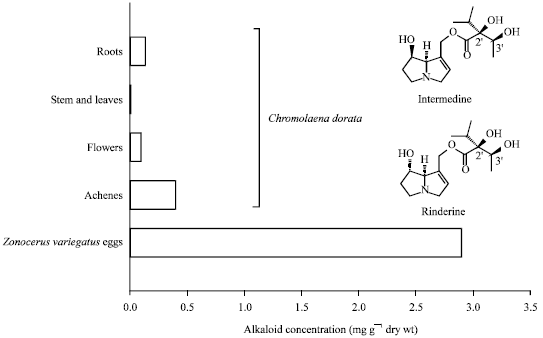 | |
| Fig. 5: | Comparing the concentration of total PAs found in various organs of Chromolaena odorata and eggs of Zonocerus variegatus. Insert: Structure formulas of the major PAs found in the plant and sequestered in the eggs of the grasshopper |
| Table 5: | Pyrrolizidine alkaloids identified by GC-MS in various organs of C. odorata from two distinct geographic origins as well as the eggs of Z. variegatus that as larvae and adults have had excess to flowers of C. odorata |
 | |
| nd: Plant organ was devoid of PAs | |
The relatively high levels of PAs in Z. varigatus eggs caused us to test whether there is a male-to-female transmission of PAs during mating. Females reared on cassava in the presence of dried roots of C. odorata (+PA females) were mated with males reared in absence of a PA source (-PA males). The resulting eggs were found to contain PAs in similar concentrations as found in crossings of +PA females with +PA males. By contrast, only traces of PAs could be detected in eggs resulting from crossings of -PA females and +PA males, indicating an inability of males to donate their PAs to females (Fig. 6).
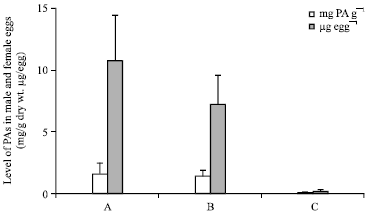 | |
| Fig. 6: | Crossing experiment indicating that females provide their eggs with PAs but do not receive PAs from males during copulation |
DISCUSSION
The results obtained in the present study show that the development of Z. variegatus reared on cassava leaves as sole food source is markedly affected if the hoppers have free access to dry roots of C. odorata as PA source. The parameters studied are influenced positively if the grasshoppers had the choice to nibble at the C. odorata roots. Two major effects were observed. Firstly, the body size of Z. variegatus was higher in the presence of the PA source as demonstrated by an increase of the body weight and the enhanced length of the thorax shield. These positive effects were always detectable. However, they were statistically significant in some but not all trial replications. Secondly, oviposition was affected in a complex manner, i.e., in the presence of C. odorata roots females produced heavier egg pods containing more but lighter eggs. In this case all documented differences were statistically significant.
Most likely the PAs acquired by the grasshoppers from C. odorata are causing the observed developmental alterations. The relationship of Z. variegatus and PA-containing plants has been well studied. All developmental stages of Z. variegatus are attracted by plant tissues containing PAs or by pure alkaloids (Boppre et al., 1984; Fischer and Boppre, 1997) indicating the ability of the insect to specifically recognize volatiles most likely released as decomposition products of PAs such as hydroxydanaidal and related structures that are also known from certain PA sequestering arctiid moths as male courtship pheromones (Schulz, 1998; Schulz et al., 2004). Grasshoppers lured to a PA source immediately begin eagerly consuming the PA-containing plant food like flowers of C. odorata (Modder, 1984, 1986). However, C. odorata does not allow normal development of Z. variegatus (Bernays et al., 1975; McCaffery et al., 1978). Both leaves and flowers of C. odorata are eaten by all nymphal stages and adults of Z. variegatus (Modder, 1984; Marks and Seddon, 1985) but not as exclusive food. Leaves of other plants are eaten at the same time; Z. variegatus needs a mixed diet with C. odorata as just one item (Modder, 1984, 1986; Chapman et al., 1986). Obviously C. odorata is preferentially exploited as drug source by Z. variegatus as suggested by (Boppre et al., 1984), a phenomenon referred to as pharmacophagy (Boppre, 1984, 1986). Flowers, although less suited for nutrition, are preferred to leaves because they are the better PA source (Table 5). Zonocerus variegatus is capable of sequestering PAs of different structural types such as macrocyclic alkaloids of the monocrotaline type found in many Crotalaria species (Fabaceae) (Bernays et al., 1977) or the senecionine type found in genus Senecio (Asteraceae, tribe Senecioneae) (Lindigkeit et al., 1997) as well as monoesters of the lycopsamine type as those found in C. odorata (Biller et al., 1994). Ingested PAs are specifically detoxified by N-oxidation and safely stored and maintained as non-toxic PA N-oxides (Lindigkeit et al., 1997). Moreover, as shown in this study females provide their eggs with PAs at concentrations that are 10-30-fold higher than those found in their plant PA source.
The briefly sketched relationship of Z. variegatus to PAs shows striking parallels to PA sequestering lepidopterans, particularly the well studied arctiid moths (Boppre, 1986; Hartmann, 1999; Eisner et al., 2002). Polyphagous arctiid larvae locate PA plants just incidentally but recognize them specifically through sensitive taste receptors in their mouthparts that are phagostimulatory (Bernays et al., 2002a, b). Like Z. variegatus, arctiid larvae absorb PAs as free bases and efficiently detoxify them by N-oxidation. The respective enzyme has been characterized as a flavin-dependent monooxygenase that specifically N-oxidize PAs but no other alkaloids (Lindigkeit et al., 1997; Naumann et al., 2002). As already discussed Z. variegatus cannot develop properly on its preferred PA source C. odorata. Exactly the same has been demonstrated for arctiid moths with polyphagous larvae. In the grassland of the southwestern United States larvae of E. acrea, for instance, cannot survive or develop properly on their major PA sources, e.g., Crotalaria pumila (Hartmann et al., 2005) or Senecio longilobus (Singer et al., 2004), although the PAs, themselves, do not affect larval development. Like Z. variegatus polyphagous arctiid larvae exploit PA-containing plants preferentially as “Drug source” and not for nutrition.
The role of plant-acquired pyrrolizidine alkaloids in the performance of arctiid moths has been most completely elucidated in the species Utetheisa ornatrix (Eisner et al., 2002). Larvae of this moth develop monophagously on Crotalaria spp. which functions as both nutrient and PA source. Sequestered PAs are retained through metamorphosis. At mating the male advertises to the female his alkaloidal load through the PA derived male courtship pheromone, hydroxydanaidal. This is emitted from a pair of scent brushes (coremata) which it everts during close-range precopulatory interaction with the female. Females appear to favor males having higher alkaloid levels (Conner et al., 1990). At mating, the male transmits a portion of its PAs to the female during insemination. At oviposition these alkaloids together with the female’s own load are transmitted to the eggs (Dussourd et al., 1988; Iyengar et al., 2001). This sophisticated physiological and behavioral adaptation most impressively demonstrates the importance of an efficient protection of the eggs, the most endangered life-stage in the development of an insect. Efficient egg protection by PAs of biparental origin was also demonstrated for polyphagous arctiids like Creatonotos transiens (Von Nickisch-Rosenegk et al., 1990) and Estigmene acrea (Hartmann et al., 2004b). The alkaloid endowment has been shown to protect the eggs of U. ornatrix against predation by coccinellid beetles (Dussourd et al., 1988), ants (Hare and Eisner, 1993) and chrysopid larvae (Eisner et al., 2000) and parasitization by the chalcidoid wasp, Trichogramma ostriniae (Bezzerides et al., 2004). Zonocerus variegatus males do not transmit PAs to the female (Fig. 6) but the average PA level (3 mg g-1 or 8 μg egg-1) is in the same order of magnitude as reported for arctiids, i.e., U. ornatrix 7 mg g-1 or ca 0.3-1.1 μg egg-1 (Eisner et al., 2000) and E. acrea 1.7-4 mg g-1 or 0.03-0.24 μg egg-1 (Hartmann et al., 2004b). Thus, eggs of Z. variegatus provided with PAs should be similarly well protected against egg predators or parasitoids as arctiid eggs although the predacious species may be different. In this context the striking effects of the accessible PA source on the egg characteristics (Table 3) of cassava reared Z. variegatus should be noticed. It is known that the size of the egg pods and the number of eggs per pod may vary from large pods with more than 100 eggs per pod to small pods with less than 30 eggs per pod (Tamu, 1995).
It appears likely that the presence of PAs increases the pod size and number of eggs but a conclusive proof requires additional experiments with pure PAs in order to exclude possible influence of other compounds contained in the dried root of C. odorata applied as PA source. Such experiments are also needed to confirm the various effects observed in presence of the PA source. It is intriguing to assume that PA sequestration may affect the development of Z. variegatus in a different but also meaningful manner as already demonstrated for the above discussed arctiid moths.
ACKNOWLEDGMENTS
The authors are greatly indebted to Prof. Dr. Michael Boppré and Mr. Ottmar W. Fischer, both of the Forstzoologisches Institute, Freiburg for helpful comments on the original document. We are also thankful to Klaus Kiesel (Freiburg) and Claudine Theuring (Braunschweig) for excellent technical assistance. The work by Till Beuerle (Braunschweig) for performing the GC-MS analyses is appreciated. The field work was done with funds from the British Ecological Society. The work carried out in Braunschweig was supported by a grant of the ‘Deutsche Forschungsgemeinschaft’ to T.H.
REFERENCES
- Bernays, E.A., J.A. Edgar and M. Rothschild, 1977. Pyrrolizidine alkaloids sequestered and stored by the aposematic grasshopper, Zonocerus variegatus. J. Zool., 182: 85-87.
CrossRefDirect Link - Bernays, E., R. Chapman and T. Hartmann, 2002. A highly sensitive taste receptor cell for pyrrolizidine alkaloids in the lateral galeal sensillum of a polyphagous caterpillar, Estigmene acraea. J. Comp. Physiol. A., 188: 715-723.
CrossRefDirect Link - Bernays, E.A., R.F. Chapman and T. Hartmann, 2002. A taste receptor neurone dedicated to the perception of pyrrolizidine alkaloids in the medial galeal sensillum of two polyphagous arctiid caterpillars. Physiol. Entomol., 27: 312-321.
CrossRef - Bezzerides, A., T.H. Yong, J. Bezzerides, J. Husseini, J. Ladau, M. Eisner and T. Eisner, 2004. Plant-derived pyrrolizidine alkaloid protects eggs of a moth (Utetheisa ornatrix) against a parasitoid wasp (Trichogramma ostriniae). Proc. Nat. Acad., 101: 9029-9032.
PubMed - Boppre, M., 1986. Insects pharmacophagously utilizing defensive plant chemicals (Pyrrolizidine alkaloids). Naturwissenschaften, 73: 17-26.
CrossRefDirect Link - Boppre, M., U. Seibt and W. Wickler, 1984. Pharmacophagy in grasshoppers? Zonocerus attracted to and ingesting pure pyrrolizidine alkaloids. Entomologia Experimentalis Applicata, 35: 115-117.
CrossRefDirect Link - Chapman, R.F., W.W. Page and A.R. McCaffery, 1986. Bionomics of the variegated grasshopper (Zonocerus variegatus) in West and Central Africa. Ann. Rev. Entomol., 31: 479-505.
CrossRefDirect Link - Cobbinah, J.R. and G.K. Tuani, 1992. Antifeedant effect of Jatropha curcas L. seed oil and extracts on the variegated grasshopper, Zonocerus variegatus L. (Orthoptera: Arcridae). J. Univ. Sci. Tech., 12: 136-139.
CrossRef - Conner, W.E., B. Roach, E. Benedict, J. Meinwald and T. Eisner, 1990. Courtship pheromone production and body size as correlates of larval diet in males of the arctiid moth, Utetheisa ornatrix. J. Chem. Ecol., 16: 543-552.
CrossRefDirect Link - Dussourd, D.E., K. Ubik, C. Harvis, J. Resch, J. Meinwald and T. Eisner, 1988. Biparental defensive endowment of eggs with acquired plant alkaloid in the moth Utetheisa ornatrix. Proc. Nat. Acad. Sci., 85: 5992-5996.
PubMed - Fischer, O.W. and M. Boppre, 1997. Chemoecological Studies Reveal Causes for Increased Population Densities of Zonocerus (Orth: Pyrgomorphidae) and offer new means for Management. In: New Strategies in Locust Control, Krall, S., R. Peveling and D.B. Diallo (Eds.). Birkhauser Verlag Basel, Switzerland, ISBN: 13-9783764354428, pp: 267-282.
- Hare, J.F. and T. Eisner, 1993. Pyrrolizidine alkaloid deters ant predators of Utetheisa ornatrix eggs: Effects of alkaloid concentration, oxidation state and prior exposure of ants to alkaloid-laden prey. Oecologia, 96: 9-18.
CrossRefDirect Link - Hartmann, T., 1999. Chemical ecology of pyrrolizidine alkaloids. Planta, 207: 483-495.
CrossRefDirect Link - Hartmann, T., 2004. Plant-derived secondary metabolites as defensive chemicals in herbivorous insects: A case study in chemical ecology. Planta, 219: 1-4.
CrossRefDirect Link - Hartmann, T., C. Theuring, T. Beuerle, L. Ernst, M.S. Singer and E.A. Bernays, 2004. Acquired and partially De novo synthesized pyrrolizidine alkaloids in two polyphagous arctiids and the alkaloid profiles of their larval food-plants. J. Chem. Ecol., 30: 229-254.
CrossRefDirect Link - Hartmann, T., C. Theuring, T. Beuerle and E.A. Bernays, 2004. Phenological fate of plant-acquired pyrrolizidine alkaloids in the polyphagous arctiid Estigmene acrea. Chemoecology, 14: 207-216.
CrossRefDirect Link - Hartmann, T., C. Theuring, T. Beuerle, N. Klewer, S. Schulz, M.S. Singer and E.A. Bernays, 2005. Specific recognition, detoxification and metabolism of pyrrolizidine alkaloids by the polyphagous arctiid Estigmene acrea. Insect Biochem. Mol. Boil., 35: 391-411.
CrossRefDirect Link - Iyengar, V.K., C. Rossini and T. Eisner, 2001. Precopulatory assessment of male quality in an arctiid moth (Utetheisa ornatrix): Hydroxydanaidal is the only criterion of choice. Behav. Ecol. Sociobiol., 49: 283-288.
CrossRefDirect Link - Lindigkeit, R., A. Biller, M. Buch, H.M. Schiebel, M. Boppre and T. Hartmann, 1997. The two faces of pyrrolizidine alkaloids: The role of the tertiary amine and its N‐oxide in chemical defense of insects with acquired plant alkaloids. Eur. J. Biochem., 245: 626-636.
CrossRefDirect Link - Marks, M.K. and A.M. Seddon, 1985. Interaction between Chromolaena odorata (Compositae) and Zonocerus variegates (Orthoptera: Pyrgomorphidae) with emphasis on achene predation. Afr. J. Agric. Sci., 12: 29-37.
Direct Link - McCaffery, A.R., A.G. Cook, W.W. Page and T.J. Perfect, 1978. Utilisation of food by Zonocerus variegates (L.) (Orthoptera: Pyrgomor phidae). Bull. Entomol. Res., 68: 589-606.
CrossRefDirect Link - Modder, W.W.D., 1984. The attraction of Zonocerus variegates (L.) (Orthoptera: Pyrgomorphidae) to the weed Chromolaena odorata and associated feeding behaviour. Bull. Entomol. Res., 74: 239-247.
CrossRefDirect Link - Modder, W.W.D. and S.R. Singh, 1976. Utilisation of cassava Manihot esculenta, Rev. Zool. Afri., 90: 417-430.
Direct Link - Modder, W. and G.F. Tamu, 1996. The effect of food plants on metabolic reserves, development and fecundity in the African pest grasshopper, Zonocerus variegates (Linnaeus) (Orthoptera: Pyrgomorphidae). Afr. Entomol., 4: 189-196.
Direct Link - Naumann, C., T. Hartmann and D. Ober, 2002. Evolutionary recruitment of a flavin-dependent monooxygenase for the detoxification of host plant-acquired pyrrolizidine alkaloids in the alkaloid-defended arctiid moth Tyria jacobaeae. Proc. Nat. Acad. Sci., 99: 6085-6090.
CrossRefDirect Link - Von Nickisch-Rosenegk, E., D. Schneider and M. Wink, 1990. Time-course of pyrrolizidine alkaloid processing in the alkaloid exploiting arctiid moth, Creatonotos transiens. Z Naturforsch, 45: 881-894.
Direct Link - Pasteels, J.M. and T. Hartmann, 2004. Sequestration of Pyrrolizidine Alkaloids in Oreina and Platyphora Leaf Beetles: Physiological, Ecological and Evolutionary Aspects. In: New Developments in the Biology of Chrysomelidae, Jolivet, P., J.A. Santiago-Blay and M. Schmitt (Eds.). VSP Academic Publ., Netherlands, ISBN-13: 978-9051031423, pp: 677-691.
- Schulz, S., 1998. Insect‐plant interactions-metabolism of plant compounds to pheromones and allomones by lepidoptera and leaf beetles. Eur. J. Organic Chem., 1998: 13-20.
CrossRefDirect Link - Schulz, S., G. Beccaloni, K.S. Brown Jr., M. Boppre, A.V.L. Freitas, P. Ockenfels and J.R. Trigo, 2004. Semiochemicals derived from pyrrolizidine alkaloids in male ithomiine butterflies (Lepidoptera: Nymphalidae: Ithomiinae). Biochem. System. Ecol., 32: 699-713.
CrossRefDirect Link - Singer, M.S., D. Rodrigues, J.O. Stireman III and Y. Carriere, 2004. Roles of food quality and enemy-free space in host use by a generalist insect herbivore. Ecology, 85: 2747-2753.
CrossRefDirect Link - Tamu, G.F., 1995. Location and categorization of Zonocerus variegates L. orthoptera: Acridoidea: Pyrgomorphidae egg pods and evaluation of egg destruction as a control measure. J. Afr. Zool., 1094: 329-338.
Direct Link - Toye, S.A., 1982. Studies on the biology of the grasshopper pest Zonocerus variegatus (L.) (Orthoptera: Pyrgomorphidae) in Nigeria: 1911-1981. Int. J. Trop. Insect Sci., 3: 1-7.
CrossRefDirect Link - Weber, S., W. Eisenreich, A. Bacher and T. Hartmann, 1999. Pyrrolizidine alkaloids of the lycopsamine type: Biosynthesis of trachelanthic acid. Phytochemistry, 50: 1005-1014.
CrossRefDirect Link - Witte, L., P. Rubiolo, C. Bicchi and T. Hartmannt, 1992. Comparative analysis of pyrrolizidine alkaloids from natural sources by gas chromatography-mass spectrometry. Phytochemistry, 32: 187-196.
CrossRefDirect Link