
M. Barros
Departament of Biosciences and Biochemistry, Faculty of Chemistry, Universidad de la Rep�blica, Avda. Gral. Flores 2124, CP 11800, Montevideo, Uruguay
A.M.B. Cantera
Laboratory of Hydrolytic Enzyme, Institute of Biological Chemistry, Faculty of Sciences, Universidad de la Rep�blica, Equals 4225, CP 11400, Montevideo, Uruguay
Research Journal of Phytochemistry
Year: 2014 | Volume: 8 | Issue: 4 | Page No.: 172-187







ABSTRACT
Characterization studies were performed to a proteolytic extract, prepared from latex of Ficus luschnathiana (Miq.) Miq., a Moraceae family tree. The mechanism of action was determined, corresponding to serine proteinases behaviour. Studies of optimum pH and temperature showed a maximum catalytic activity at pH 8.0 and 75°C, respectively. The proteolytic activity exhibited good stability when incubated at different pH and temperature conditions. At 7°C and pH 7.5, the extract kept a 100% of the initial activity for 5 months. When comparing the effect of pH on the stability, maximum values were obtained at pH 6.5, with 98% of the initial activity remaining after 5 h of incubation at 60°C. After 1 min at 100°C total inactivation occurred. Regarding the potential use in the dairy industry, milk clotting activity was determined obtaining a very effective coagulant action. Analysis protocols which included molecular exclusion chromatography, electrophoretic methods and mass spectrometry revealed a complex protein profile. It was found that the molecules responsible for the proteolytic activity of the extract were several proteins of different molecular weight and isoelectric points, most of which still retained the activity in presence of SDS and reducing agents. These proteic components displayed a pH-dependant tendency to form aggregates. Using the proteolytic activity of the crude extract under controlled hydrolysis conditions on whey proteins, yielded peptidic fractions with antimicrobial activity against a S. aureus strain. It was found that fractions of peptide concentration of 0.96 μmol mL-1 inhibited growth by 94%.
PDF Abstract XML References Citation
Received: August 15, 2014;
Accepted: October 13, 2014;
Published: November 13, 2014
How to cite this article
M. Barros and A.M.B. Cantera, 2014. Characterization of the Proteolytic Activity of Latex of Ficus luschnathiana
and its Application in the Generation of Whey Antimicrobial Peptides. Research Journal of Phytochemistry, 8: 172-187.
URL: https://scialert.net/abstract/?doi=rjphyto.2014.172.187
URL: https://scialert.net/abstract/?doi=rjphyto.2014.172.187
INTRODUCTION
The wide range of biotechnological applications of proteolytic enzymes have generated in recent years increased efforts to search for new sources that provide activity profiles adaptable to current demands. This, coupled with the convenience of using natural resources in each region, has turned much of the interest of researchers towards the native flora.
The plant proteolytic enzymes have received particular attention due to the property to maintain its activity in a wide range of temperature and pH (Devaraj et al., 2008). They play an important role in regulating the biological processes in plants, such as stress response, recognition of pathogen, mobilization of storage protein during germination, senescence, etc. (Singh et al., 2008).
The latex is considered a major source of plant proteases and is well-known its use in folk medicine as well as in different industrial processes (Liggieri et al., 2009); for example, latices from Asclepias spp. are used in wound healing and the treatment of some digestive disorders (Obregon et al., 2009) and it has been reported a variety of traditional uses and pharmacological properties of Ficus religiosa (Singh et al., 2011). This vegetal secretion is a milky fluid that contains, either in solution or suspension, a mixture of substances which can be enzymes such as glycosidases, proteases, phosphatases, amylases, chitinases, glucanases, etc., which would be involved in the mechanisms of plant defense against pathogens and insects (Pereira et al., 1999).
The first aim of this study was the proteolytic activity characterization of the crude extract obtained from the latex de Ficus lushnathiana (Miq.) Miq., Moraceae family laticiferous tree, indigenous to subtropical America (Argentina, Brasil, Paraguay and Uruguay). The genus Ficus contains more than 1800 named species making them one of the largest genuses in the Moraceae family. The ficins (name used to describe proteolytic enzymes of latex of the genus Ficus) isolated from different species possess different characteristic properties (Devaraj et al., 2008). Inhibitors were used to elucidate the mechanistic nature of the occurring enzymes, the optimal reaction conditions (pH and temperature) were determined and stability studies at different storage conditions were performed.
The second aim of the present study consisted of determining of the molecular features of the proteases present in the extract. Using molecular exclusion chromatography, fractions enriched in different proteic components of the extract were obtained. The molecular weight profile as well as the tendency to form aggregates depending on the acidity of the medium were determined for this fractions.
Finally, controlled hydrolysis of whey proteins using the proteolytic activity of the crude extract of F. lushnathiana (Miq.) Miq. were done. Wheys are considered co-products of the dairy industry due to the potential use in the food industry as protein sources of excellent nutritional value and as functional food ingredients. Wheys derived from Mozzarella cheese and casein production were used. Due at that the majority of reported bioactive peptides are derived from milk proteins and have special relevance those with antimicrobial properties (Pellegrini, 2003; Clare et al., 2003), at the obtained peptidic fractions it was evaluated the ability to inhibit the growth of a S. aureus strain.
MATERIALS AND METHODS
Materials: All the reagents used were purchased from Sigma-Aldrich.The Ficus luschnathiana (Miq.) Miq. specimens (Herbarium n°MVJB28429) belong to the Museo y JardínBotánicoAtilio Lombardo (Montevideo, Uruguay). Whey derived of Mozzarella cheese production (Wm) was supplied by a national dairy industry (CONAPROLE). Whey derived from casein production (Wc) was prepared by precipitating the casein at pH 4.5 by addition of 2 N HCl.
Crude enzyme extract preparation: Latex, obtained by superficial incisions of stems, was collected on 0.2 M sodium phosphate buffer (pH 7.5). This suspension was centrifuged at 15493xg for 1 h at 10°C in order to discard gums and other insoluble materials.
Preliminary purification of crude extract: About 0.3 mL of crude extract was applied to a column packed with Sephadex G-75 (10.0x1.5 cm), equilibrated with 50.0 mM sodium acetate buffer (pH 5.5), 50.0 mM sodium phosphate buffer (pH 6.4) or 50.0 mM Tris-HCl buffer (pH 8.4), depending on the pH required for subsequent experiments. Protein elution was performed with the same buffer and 1.5 mL fractions were collected and screened for proteolytic activity. The protein profile was measured at 280 nm.
Protein determination: Protein content was determined by the method of Lowry (Layne, 1957) using bovine serum albumin as standard. During chromatographic purification steps, protein concentration of each fraction was estimated firstly by measuring absorbance at 280 nm, wavelength at which the aromatic amino acid residues absorb. Fractions with proteolytic activity were also measured by Lowry method.
Proteolytic activity determination: It was determined as described by Andrews and Asenjo (1986), using azocasein as substrate. The Enzyme Unit (EU) was defined as the amount of enzyme necessary to increase in 1 min one unit the A337 at pH 7.5 and 37°C.
Milk clotting activity: Was performed by visual evaluation of the occurrence of the first clotting flakes as reported by Castro and Cantera (1995). The Milk Clotting Unit (MCU) was defined as the amount of enzyme needed to form the first detectable coagulated milk in 1 min under the experimental conditions.
Effect of inhibitors: Crude extract was incubated with each of the inhibitors solutions for 1 h at 25°C and the remaining proteolytic activities were determined. The inhibitors used and their final concentrations were L-trans-epoxysuccinyl-leucylamide-(4-guanidino)-butane (E-64) 5.0 and 10.0 μM, pepstatin A 1.0 μM, phenylmethylsulphonyl fluoride (PMSF) 0.5 and 1.0 mM, ethylenediaminetetraacetic acid (EDTA) 5.0 and 10.0 mM and iodoacetamide 100.0 μM. In the case of pepstatin A, pH of the crude extract was adjusted to 6.0 with glacial acetic acid while for the others pH 7.5 was used. Inhibition percentages were calculated considering a control without inhibitor.
Optimal conditions of pH and temperature: To determine the optimum pH proteolytic activities were measured at 37°C at pH 5.2, 6.0, 6.4, 6.7, 7.1, 7.6, 7.9, 8.8 and 9.2 using the following buffer solutions: 0.5 M sodium acetate buffer (pH 5.0 and 5.5), 0.5 M sodium phosphate buffer (pH 6.0, 6.5, 7.0, 7.5 and 8.0) and 0.5 M Tris-HCL buffer (pH 8.5 and 9.0). To determine the optimum temperature proteolytic activities were measured at pH 7.5 at 8, 24, 30, 37, 50, 60, 70, 75, 80, 85 and 90°C.
Stability studies: To determine the effect of temperature on the stability of protease preparation at pH 7.5, crude extract was incubated at -20, 7, 22, 37, 60 and 100°C, respectively. Aliquots were taken at different intervals and the remaining activities were determined at 37°C. Intervals between the aliquots were counted in days for the lower temperatures (-20 to 37°C), in hours for 60°C and in minutes for 100°C due to increased protein destabilization speed with increasing temperature. To determine the effect of pH, crude extract was incubated at pH 5.2, 6.5, 7.5 and 9.9, respectively, at 60°C. Glacial acetic acid was used to achieve the pH 5.2 and 6.5. The pH 9.9 was obtained adding 2 N NaOH. Aliquots were taken at different intervals and the remaining activities were measured at pH 7.5 according the activity assay described before.
pI measurement: The pI of the proteins was determined by isoelectric focusing using Phast System equipment (Pharmacia, Uppsala, Sweden) IEF 3-9. Gels were revealed by silver staining according to the manufacturer's instructions.
Native electrophoresis: Samples were prepared by adding 10% glycerol and 0.25% (w/v) bromophenol blue and then loaded on 11% polyacrylamide gel. Electrophoresis was performed in a vertical gel apparatus (Hoefer SE 250 Mighty Small II) using 25 mMTris, 0.2 M glycine (pH 8.6), as reservoir buffer. Current was kept constant at 50 mA. The gels were stained with Coomassie Brilliant Blue R 250 in methanol/acetic acid/water (1/1/8, v/v/v) solution.
Molecular mass determination: Electrophoresis was performed in a vertical gel apparatus (Hoefer SE 250 Mighty Small II) on a 12% polyacrylamide gel, in the presence of sodium dodecyl sulphate (SDS-PAGE). Protein samples were prepared by adding 50% (v/v) of sample buffer (0.19 M Tris-phosphate buffer, 0.4 M β-mercaptoethanol, 6% SDS, 30% glycerol, 0.4% bromophenol blue), followed by heating for 5 min in boiling water bath. Current was kept constant at 50 mA. The gels were stained with Coomassie Brilliant Blue R 250 in methanol/acetic acid/water (1/1/8, v/v/v) solution.
Zymogram: Unstained gels from PAGE were contacted for 10 min at 50°C with an agarose gel imbibed with a 1.0% casein solution. After incubation, the agarose gel was dehydrated and stained using Coomassie Brilliant Blue R-250 in methanol/acetic acid/water (1/1/8, v/v/v) solution. Unstained bands revealed proteolytic activity. To detect active proteases in the presence of SDS and reducing agent, SDS-PAGE was performed treating samples as described above in 2.12 but without heating and using a 12% polyacrylamide gel supplemented with 0.2% casein instead. After each electrophoresis, gels were incubated for 2 h at 22°C before staining with Coomassie solution. Unstained bands revealed proteolytic activity.
Mass spectrometry: The protein profile was performed by Matrix Assisted Laser Desorption Ionization (MALDI) and Time of Fly (TOF) analysis (Voyager DE-Pro, AbiSciex.).
pH-dependentoligomeric states of the proteins: After the preliminary purification of crude extract with Sephadex G-75 at pH 5.5 and pH 8.4, fractions with highest proteolytic activity obtained at each pH were, respectively lyophilized and then redissolved in 0.30 mL of distilled water. Both resulting samples were applied on HiLoad 16/60 Superdex 75 preparative grade column equilibrated with 50 mM sodium acetate buffer (pH 5.5) or 50 mMTris-HCl buffer (pH 8.4), respectively, at a flow rate of 1.0 mL min-1. Column was calibrated with thyrogloblin (670 kDa), aldolase (158 kDa), albumin (67 kDa), ovalbumin (44 kDa), chymotrypsinogen A (25 kDa), myoglobulin (17 kDa) and ribonuclease A (13.7 kDa). The resulting equation was log MW = -0.025, EV+3.382; R2 = 0.99 (MW = molecular weight and EV = elution volume).
Hydrolysis of whey proteins: Whey was incubated with the enzymatic preparation under defined pH and temperature conditions (pH = 7.2, 50°C), at specified enzyme/substrate ratio (0.01 EU mg-1). Aliquots were taken at different times (0, 15, 30, 60, 90 and 120 min) and placed in boiling water bath for 5 min to stop the reaction. The progress of hydrolysis was evaluated using TNBS, as described by Spadaro et al. (1979). The degree of hydrolysis (DH%) was defined as the percentage of peptide bonds cleaved, according to the equation:
where, h is the hydrolysis equivalent formed during enzymatic action and htot is the hydrolysis equivalent after acid hydrolysis in 6 N HCl, at 110°C, for 24 h in sealed tubes. Hydrolysis equivalents were calculated using a calibration curve with valine as standard: y = 0.2995x+0.0215, R2 = 0.993.
Fractionation of the products of hydrolysis: Hydrolysis products obtained at different times were fractionated using AMICON Ultra-4 centrifugal filter devices (3 and 10 kDa).
Evaluation of antimicrobial activity: Antimicrobial activity against S. aureus was screened by bioautography applying 50 μL of each fraction obtained in 2.17 on alumina thin-layer plates. Quantification of inhibition was done in microtiter plates with an inoculum of 106 colony forming units per mililiter (CFU mL-1) (Hadacek and Greger, 2000). Bacitracin was used as a positive control.
Determination of the peptidic concentration: The peptidic concentration was determined using o-phthaldialdehyde as described by Church et al. (1983).
Statistical analysis: Reported data was the average values of standard deviations of triplicate assays. Existence of statistically significant differences between results was evaluated by analysis of variance (ANOVA).
RESULTS
Milk clotting activity: The milk clotting activity of the crude extract was compared with that obtained using a trypsin solution. Both enzyme solutions were prepared so as to have the same activity by the azocasein method. Clotting times obtained for both are shown in Table 1. Crude extract showed higher milk clotting activity than the reference solution.
Inhibition studies: Results obtained in inhibition studies are summarized in Table 2. The result obtained after the action of PMSF was the same for the two concentrations tested. The total inhibition obtained suggests that the proteolytic activity of the extract is due to serine proteinases.
| Table 1: | Clotting Times (CT) and Milk Clotting Units (MCU) of a trypsin solution and crude extract (both with 4.4 EU mL-1) |
 | |
| aEach value is the mean of three independent determinations, ±: Indicate the standard deviation | |
| Table 2: | Inhibition percentages of the proteolytic activity |
 | |
| aEach value is the mean of three independent determinations, ±: Indicate the standard deviation | |
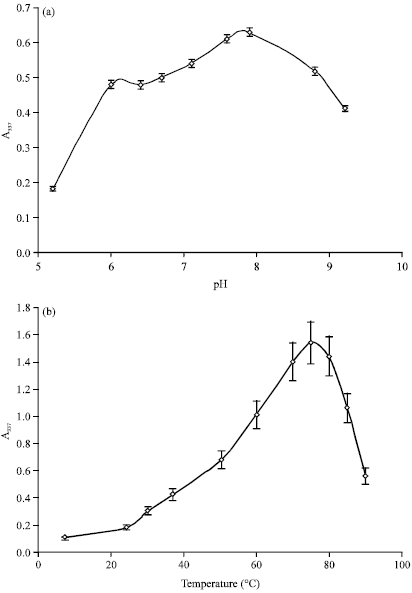 | |
| Fig. 1(a-b): | Effect of (a) pH and (b) Temperature on the hydrolysis of 1.0% azocasein in the presence of the crude extract obtained from the latex of Ficus luschnathiana |
Although this compound can also act on cysteine enzymes, the lack of inhibition observed for E-64and that the activity assay was not affected by the presence of mercaptans confirms the absence of these type of proteinases. Negative results obtained both with EDTA and Pepstatin A indicate a lack of metal and aspartic proteases activity respectively. The percentage of inhibition achieved with iodoacetamide can be explained by the fact that its action is not specific for cysteine enzymes, but is also capable of inhibiting certain serine proteinases (Salvesen and Nagase, 2001).
Optimal conditions of pH and temperature: Figure 1a shows the results of the hydrolyzing activity as a function of pH. Optimal activity was observed at pH 8.0 and each of the results obtained for pH in the range 5.5-9.0 are at least a 60% of the maximum value. Results of temperature-activity are shown in Fig. 1b. The maximum value obtained was at 75°C. Even at 8°C proteolysis was observed (24% compared to that observed in the standard conditions of pH 7.5 and 37°C).
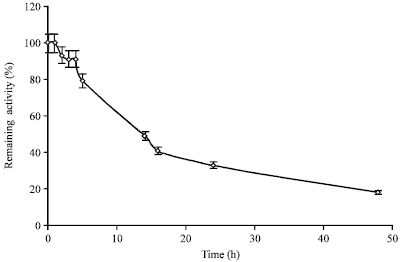 | |
| Fig. 2: | Inactivation kinetics at 60°C of crude extract at pH 7.5 |
| Table 3: | Percentagesa of remaining activity of the extract after incubation at different temperatures |
 | |
| aEach value is the mean of three independent determinations, ±: Indicate the standard deviation | |
| Table 4: | Remaining activity (%)a of the crude extract after incubation at different pH (60°C) |
 | |
| aEach value is the mean of three independent determinations, ±: Indicate the standard deviation | |
Stability at different temperatures: The results of determinations of stability of the extract at different temperatures are summarized in Table 3. Freezer storage (-20°) adversely affects activity of the extract. Moreover, the results suggest the possibility of storage for prolonged times (upto 5 months) at 7°C. Even at room temperature (22°C) the remaining activity values obtained were quite high for two months and after 15 days only 3% of the activity was lost. The behaviour at 60°C is illustrated in Fig. 2. It can be seen that the remaining activity did not change after 1 h of incubation and decreased to 79% after 5 h. The experimental half life observed was 14 h. As at 100°C the extract was completely inactivated after 1 min of incubation.
Stability at different pH: The results of the stability determinations at different pH are summarized in Table 4. In order to visualize promptly the decrease of the activity remaining, the temperature selected for the study was 60°C. The extract was more stable at moderately acidic conditions and the highest values of remaining activity were obtained at pH 6.5. The lowest values on the other hand were obtained in the most alkaline medium. Differences in the slopes of these curves occurred in the first 5 h of incubation, after that all the inactivation rates were similar for all the pH tested.
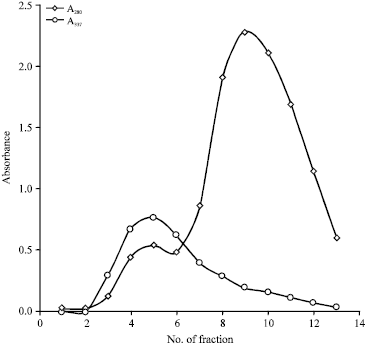 | |
| Fig. 3: | Chromatographic elution profile of crude extract on Sephadex G-75 column |
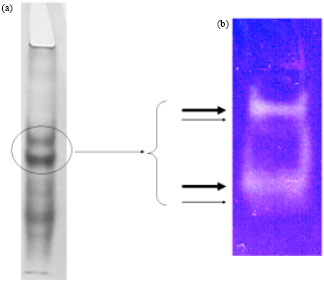 | |
| Fig. 4(a-b): | Native electrophoresis of the most active fractions obtained after the preliminary purification of crude extract by (a) Gel filtration and (b) Zymogram showing four active bands |
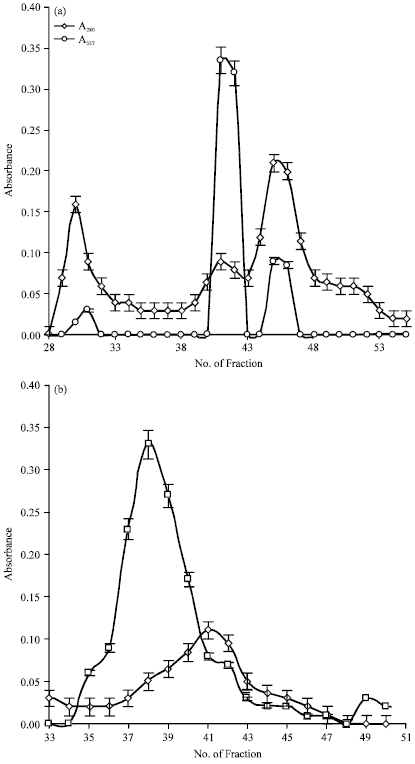
Molecular characteristics of the extract proteins: Molecular analysis of protein species present in the extract revealed a mixture of great complexity. In order to remove several non-proteins components that absorb at 280 nm including the polyphenols, preliminary purification of crude extract by exclusion chromatography with G-75 was performed at pH 6.4 which resulted in the elution profile shown in Fig. 3. The A337 values are the results of proteolytic activity assay performed on each fraction. Fractions 4, 5 and 6 were pooled, lyophilized and then redissolved in 0.25 mL of distilled water. The resulting sample was checked by native electrophoresis (Fig. 4a) and zymogram (Fig. 4b). Coomassie staining showed at least 11 proteins bands.
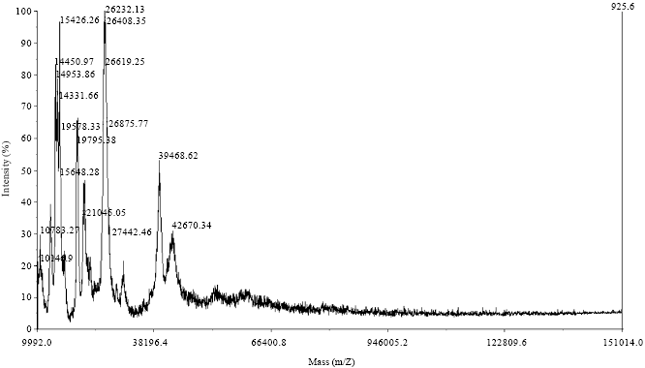 | |
| Fig. 5: | Mass spectroscopy of the pooled 4-6 fractions obtained by exclusion chromatography |
Those with the lowest charge to radius ratio showed significant proteolytic activities evidenced as two broad undyed bands and each of them was closely accompanied by another very faint band. With the aim to determine the molecular weight profile of the pooled fractions mass spectrometry (MALDI-TOF) was carried out. The spectrum obtained is shown in Fig. 5. It can be seen that it is a very complex sample with a great diversity of molecular size compounds of about 10-40 kDa. Compounds with molecular masses lower than 20 kDa probably could have been generated by autolytic processes as was previously reported for ficins of F. carica (Azarkan et al., 2011).
SDS-PAGE of each of the 4, 5 and 6 fractions obtained by exclusion chromatography with G-75 column, with and without a previous treatment of heating for 5 min in boiling water bath (Fig. 6a) provided additional data and apparently contrasting with that obtained by MALDI. These fractions had the same molecular weight profile differing only in the relative amounts of each molecular species. By comparing with molecular weight standards, molecular species were predicted having higher molecular weight values than those found by MALDI. Heat treated samples exhibited a band corresponding to approximately 60 kDa and also bands of lower molecular weight with values equivalent to those determined with MALDI. Samples without thermal treatment exhibited even higher molecular weights, greater than 80 kDa but the 60 kDa band is not present.
Electrophoresis in polyacrylamide gel supplemented with 0.2% casein (Fig. 6b) for non-heat treated samples indicated active proteases in presence of SDS and reducing agent, mainly higher molecular weight species. Also there is a less clearly unstained area, corresponding to a lower molecular weight of about 20 kDa, plus several other very faint bands of molecular sizes in between.
Oligomeric states: The facts that for a single fraction eluted from a gel filtration (pH 6.4) was obtained a broad range of molecular sizes (20 to about 100 kDa) in a SDS-PAGE (pH 8.9) as well as the observed discrepancy between the molecular weight values obtained by MALDI and by electrophoresis (above 60 kDa only for the latter) can not be explained only by quaternary structures.
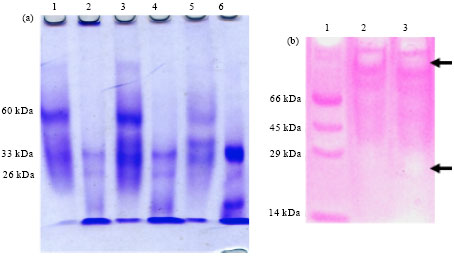 | |
| Fig. 6(a-b): | (a) PAGE-SDS of fractions 4, 5 and 6 without heat treatment for Lanes: 1, 3 and 5, respectively and with a heat treatment of 5 min in boiling water for Lanes: 2, 4 and 6, respectively and (b) Electrophoresis in polyacrylamide gel supplemented with 0.2% casein. Lane 1: Standards, Lanes 2, 3: Fractions 5 and 6, respectively, without thermal treatment. Arrows indicate unstained areas more evident |
A possible explanation is that the molecular species present in the crude extract are grouped in different ways depending on the pH of the medium and that the largest clusters are formed at alkaline pH. Heat treatment is probably capable of breaking the largest aggregates but fails to eliminate the forces that hold together the grouping of 60 kDa or may be that this grouping is reorganized during the electrophoretic run. In order to confirm these hypotheses, an exclusion chromatography of the crude extract at pH 5.5 and 8.4 with a HiLoad 16/60 Superdex 75 preparative grade column pre-calibrated with molecular weight standards was perfomed. The respective elution diagrams are shown in Fig. 7a, b. Three peaks were obtained at pH 5.5, a small one that eluted in a volume corresponding to dead volume (MW = 80 kDa) and two major peaks corresponding to 67 and 44 kDa. At pH 8.4 the resolution of the sample decreased: a large peak eluted in a volume corresponding to MW = 80 kDa, followed by a small plateau (62-70 kDa).
PAGE-SDS (with and without thermal treatment of samples) and zymogram (data not shown) were performed to fractions 41 (F41) and 45 (F45) of the gel filtration done at pH 5.5. For both fractions, molecular sizes profiles evidencing new regroupings were obtained. For F41 without thermal treatment a major band having more than 100 kDa with strong proteolytic activity and two very faint bands of approximately 100 and 39 kDa, respectively were observed while with thermal treatment there was only an intense band of 39 kDa. For F45 without thermal treatment two intense bands, a wide one of about 100 kDa and a thin band of about 80 kDa, both with proteolytic activity while with thermal treatment there were bands corresponding to 42-49 kDa. Additionally, isoelectric focusing was performed to these fractions and results are shown in Fig. 8. While there is a differential enrichment for both samples (F41 had a preponderance of higher pI molecules and in F45 there was a greater abundance of those of lower pI), there was no net resolution, showing both lanes a similar and very complex bands distribution.
 | |
| Fig.7(a-b): | Chromatographic elution profile of crude extract on HiLoad 16/60 Superdex 75 at pH (a) 5.5 and (b) 8.4 |
Hydrolysis of whey proteins: The degree of hydrolysis for each whey are shown in Fig. 9. It can be seen that whey derived from casein production was hydrolyzed to a lesser degree than that from Mozzarella cheese reaching at 120 min of reaction values of 2.40±0.10 and 3.1±0.09%, respectively, that represent a statistically significant difference according to ANOVA.
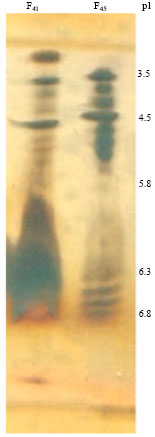 | |
| Fig. 8: | Isoelectric focusing of fraction 41 (F41) and 45 (F45) of the gel filtration performed on HiLoad 16/60 Superdex 75 at pH 5.5 |
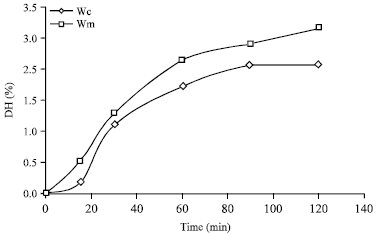 | |
| Fig. 9: | Hydrolysis curves of wheys derived from Mozzarella cheese and casein production (Wm and Wc, respectively). Reactions conditions were pH= 7.2, 50°C and enzyme/substrate ratio = 0.01 EU mg-1 |
Antimicrobial activity: Growth inhibition of a S. aureus strain was observed for both wheys with the aliquots obtained at 90 and 120 min of hydrolysis, particularly with the fractions corresponding to peptide sizes smaller than 10 kDa (Fig. 10a and b). It can be seen that although at 90 min there is a marked antimicrobial action for the fraction corresponding to sizes smaller than 10 but larger than 3 kDa, after 120 min the bioactivity is better for displaced to sizes smaller than 3 kDa. This could be because the large bioactive peptides found in first place are intermediate product of hydrolysis which are degraded afterwards resulting in the antimicrobial molecules smaller than 3 kDa evidenced after 120 min of reaction. Comparing the results for both wheys, it can be seen greater action in the case of Wm. Figure 10c shows the results for the aliquots obtained at 0-30 min of reaction. No significant results were obtained in that period showingthere is no detectable antimicrobial activity prior to hydrolysis. In none of the cases there was antimicrobial activity for fractions above 10 kDa. Although some whey proteins, such as lactoferrin (Recio and Visser, 2000), have antimicrobial activity, its low proportion in the whey, as well as the dilution factors of the method, make their activity undetectable. This shows that the hydrolysis process greatly increases the antimicrobial activity that may have the starting material.
Inhibitionquantification was done in microtiter plates. This results as well as the peptide concentration of fractions with the highest antimicrobial activity from Wm, are summarized in Table 5. The greater antimicrobial effect is observed for the fraction of lower peptidic concentration,indicating that the observed bioactivity is mainly due to the peptides of less than 3 kDa.
| Table 5: | Growth inhibition percentage of a S. aureus strain for the aliquots obtained from Wm at 120 min of hydrolysis |
 | |
| aEach value is the mean of three independent determinations, ±: Indicates the standard deviation | |
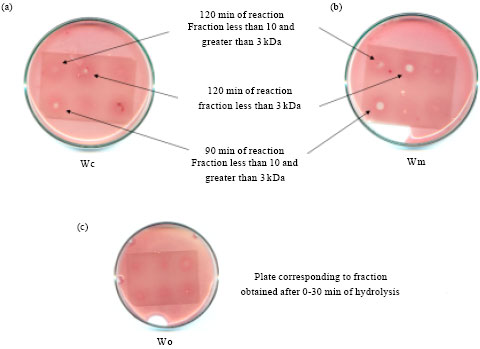 | |
| Fig. 10(a-c): | Petri dishes containing hydrolysate fractions applied on thin-layer plates. (a) Fractions from whey derived of Mozzarella cheese production (Wm), after 90 and 120 min of hydrolysis. (b) Fractions from whey derived from casein production (Wc), after 90 and 120 min of hydrolysis. (c) Fractions corresponding to the first 30 min of hydrolysis, from Wm (up) and Wc (below) |
But this does not rule out the possibility that also in the fraction corresponding to sizes between 3-10 kDa antimicrobial activity is due also to the fraction below 3 kDa, since the filtering process involves a balance between the permeate and retentate of components of lower size than the membrane pore.
DISCUSSION
The crude extract of F. luschnathiana (Miq.) Miq. latex has a marked proteolytic activity with an action model corresponding to serine proteinases. The high milk clotting activity observed opens the possibility of its use in the cheese industry, as well as in other biotechnological industries. A similar behaviour of high milk clotting activity was previously reported for religiosin B, a serine protease from Ficus religiosa, by Kumari et al. (2012).
The proteolytic action reaches its maximum at pH 8.0 and 75°C. The fact that significant values of activity were observed in a wide range of pH and temperature opens a broad spectrum of applications. The optimum values of pH and temperature found in this study are similar to those reported for other serine proteases derived from latex. Indicain, a dimeric serine protease from Morus indica showed maximum values at pH 8.5 and 80°C (Singh et al., 2008).
Stability studies showed the ability of the extract to remain unchanged for long periods at refrigeration temperature which would be economically beneficial for an industrial application because imply important economic and energy savings. The stability at high temperatures (60°C) allows its use in processes that require hours of continuous proteolytic processing. Brunius and Sundbom (1987) studied the stability of a trypsin solution from bovine pancreas at 37°C and pH 7.3. These authors reported that in these conditions the activity decreases to less than 10% after 24 h meanwhile the serinic activity in the crude extract from Ficus even after 48 h at 60°C at pH 7.5 still conserved 18% of its initial activity. The greater thermostability found for the crude extract from Ficus enables the possibility of its application in processes that require high temperatures for extended periods. Moreover, the inactivation after 1 min at 100°C makes it possible to stop the reaction without the addition of chemicals that may be incompatible in the food industry. Regarding pH, acid environment affected the protein structure of enzymes to a lesser extent than alkali.
Multiple proteases of the same family are quite often reported in latex bearing plants. However, the cause of such multiplicity has not been extensively explained (Sharma et al., 2012). The proteolytic activity of crude extract of Ficusis due to several proteic components of different molecular weight and isoelectric points, most of them still retained the activity in presence of SDS and reducing agents, making it suitable for its use in the detergent industry. These proteic components exhibited pH-dependant clustering forms, with a tendency to form larger aggregates at basic pH. In summary, the tendency to form aggregates of variable composition coupled with the possibility that some components have quaternary structure and/or different degrees of glycosylation resulted in an extract with a high level of analytical complexity. Anyway, current interest of the authors of this study lies not in obtaining pure enzymes but in evaluating the potential use of the crude extract as such and in a future work the isolation of the present peptidases.
Enzymes present in the crude extract are capable of hydrolyzing proteins ofwheys derived from Mozzarella cheese production and from casein production, yielding higher degrees of hydrolysis for the former.As a result of the hydrolytic process performed under controlled pH and temperature, fractions enriched with antimicrobial peptides capable of inhibiting upto 94% the growth of a S. aureus strain were obtained. These could help to solve problems of resistance to traditional antibiotics, an ever-increasing global threat that involves the main pathogens and drugs.
ACKNOWLEDGMENTS
This study was performed under the project CYTED IV-22.The authors acknowledge the support provided by JulianGago of Museum and Botanical Garden Atilio Lombardo, who contributed with their knowledge of indigenous flora and provided samples. The MALDI-TOFF analyses were conducted with support from the Pasteur Institute of Montevideo, Uruguay.Language corrections were done by Ph.D. Ana Acevedo.
REFERENCES
- Andrews, B.A. and J.A. Asenjo, 1986. Synthesis and regulation of extracellular β (1-3) glucanase and protease by Cytophaga sp. In batch and continuous culture. Biotechnol. Bioeng., 28: 1366-1375.
CrossRef - Azarkan, M., A. Matagne, R. Wattiez, L. Bolle, J. Vandenameele and D. Baeyens-Volant, 2011. Selective and reversible thiol-pegylation, an effective approach for purification and characterization of five fully active ficin (iso)forms from Ficus carica latex. Phytochemistry, 72: 1718-1731.
CrossRefDirect Link - Brunius, G. and B.L. Sundbom, 1987. The effect of Ca2+ on the thermal stability of trypsin in phosphate-buffered saline solution used for harvesting of human embryonic lung fibroblast cultures. J. Biol. Standardization, 15: 265-270.
CrossRefDirect Link - Castro, S. and A.M.B. Cantera, 1995. A rapid and inexpensive procedure for the determination of proteolytic activity. Biochem. Educ., 23: 41-43.
CrossRef - Church, F.C., H.E. Swaisgood, D.H. Porter and G.L. Catignani, 1983. Spectrophotometric assay using o-phthaldialdehyde for determination of proteolysis in milk and isolated milk proteins. J. Dairy Sci., 66: 1219-1227.
CrossRefDirect Link - Devaraj, K.B., L.R. Gowda and V. Prakash, 2008. An unusual thermostable aspartic protease from the latex of Ficus racemosa (L.). Phytochemistry, 69: 647-655.
CrossRefDirect Link - Hadacek, F. and H. Greger, 2000. Testing of antifungal natural products: Methodologies, comparability of results and assay choice. Photochem. Anal., 11: 137-147.
CrossRefDirect Link - Kumari, M., A. Sharma and M.V. Jagannadham, 2012. Religiosin B, a milk-clotting serine protease from Ficus religiosa. Food Chem., 131: 1295-1303.
CrossRefDirect Link - Obregon, W.D., C.S. Liggieri, S.A. Trejo, F.X. Aviles, S.E. Vairo-Cavalli and N.S. Priolo, 2009. Characterization of papain-like isoenzymes from latex of Asclepias curassavica by molecular biology validated by proteomic approach. Biochimie, 91: 1457-1464.
CrossRefDirect Link - Pellegrini, A., 2003. Antimicrobial peptides from food proteins. Curr. Pharm. Des., 9: 1225-1238.
CrossRef - Pereira, L.S., V.M. Gomes, K.V.S. Fernandes, M.P. Sales and J. Xavier-Filho, 1999. Insecticidal and antifungic proteins of the latex from Manihot glaziovii Muell. Arg. Rev. Bras. Bot., 22: 27-30.
Direct Link - Recio, I. and S. Visser, 2000. Antibacterial and binding characteristics of bovine, ovine and caprine lactoferrins: A comparative study. Int. Dairy J., 10: 597-605.
CrossRefDirect Link - Sharma, A., M. Kumari and M.V. Jagannadham, 2012. Religiosin C, a cucumisin-like serine protease from Ficus religiosa. Process Biochem., 47: 914-921.
CrossRef - Singh, D., B. Singh and R.K. Goel, 2011. Traditional uses, phytochemistry and pharmacology of Ficus religiosa: A review. J. Ethnopharmacol., 134: 565-583.
CrossRefDirect Link - Singh, V.K., A.K. Patel, A.J. Moir and M.V. Jagannadham, 2008. Indicain, a dimeric serine protease from Morus indica cv. K2. Phytochemistry, 69: 2110-2119.
CrossRefDirect Link - Spadaro, A.C.C., W. Draghetta, S.N. del Lama, A. Camargo and L.J. Greene, 1979. A convenient manual trinitrobenzenesulfonic acid method for monitoring amino acids and peptides in chromatographic column effluents. Anal. Biochem., 96: 317-321.
CrossRefDirect Link