
Nagwa G-E. Mohammady
Department of Botany and Microbiology, Faculty of Science, Alexandria University, Egypt
Christopher W. Rieken
Josephine Bay Paul Center, Marine Biological Laboratory, Woods Hole, MA, USA
Scott R. Lindell
Marine Resource Center, Marine Biological Laboratory, Woods Hole, MA, USA
Christopher M. Reddy
Department of Marine Chemistry and Geochemistry, Oceanographic Institute, Woods Hole, MA, USA
Hala M. Taha
Department of Botany and Microbiology, Faculty of Science, Alexandria University, Egypt
Connie Pui Ling Lau
Marine Resource Center, Marine Biological Laboratory, Woods Hole, MA, USA
Catherine A. Carmichael
Department of Marine Chemistry and Geochemistry, Oceanographic Institute, Woods Hole, MA, USA
Research Journal of Phytochemistry
Year: 2012 | Volume: 6 | Issue: 2 | Page No.: 42-53







ABSTRACT
A variety of batch cultured marine microalgae was selected as lipid producer candidates towards the biofuel invention. The strains were characterized and compared for their growth and potential lipid production. The results showed that, under basal cultivation conditions, Tetraselmis chuii is the highest grown and lowest lipid producer among the investigated species. Furthermore, a linear correlation between the alga's cell size and the lipid content was demonstrated. However, after being transferred into nitrogen deficient nutrient medium, T. chuii cells were equally sized and attained the maximum size after 15 days of cultivation. In addition, the lipid amount produced by the alga showed an increase by about 35% compared to the basal growth medium.
PDF Abstract XML References Citation
Received: November 02, 2011;
Accepted: February 06, 2012;
Published: February 23, 2012
How to cite this article
Nagwa G-E. Mohammady, Christopher W. Rieken, Scott R. Lindell, Christopher M. Reddy, Hala M. Taha, Connie Pui Ling Lau and Catherine A. Carmichael, 2012. Age of Nitrogen Deficient Microalgal Cells is a Key Factor for Maximizing Lipid Content. Research Journal of Phytochemistry, 6: 42-53.
URL: https://scialert.net/abstract/?doi=rjphyto.2012.42.53
URL: https://scialert.net/abstract/?doi=rjphyto.2012.42.53
INTRODUCTION
Algal lipids have been suggested as a potential fuel substitute (McGinnis et al., 1997; Abdeshahian et al., 2010; Jegannathan et al., 2011) due to their accumulation inside the cells at the end of the growth stage (Mohammady, 2007; Mohammady and Fathy, 2007; Tonon et al., 2002). Biodiesel and different biohydrocarbons are most constituent fuels extracted from algal lipid (Chisti, 2006; Dayananda et al., 2006; Ilavarasi et al., 2011). Therefore, microalgal strains with high lipid content are of great interest in the search for a sustainable feedstock for the production of biofuels (Chojnacka and Marquez-Rocha, 2004; Kotchoni and Gachomo, 2008). Griffiths and Harrison (2009) postulated that high growth rate and lipid productivity are the main key for choosing algal species for bio-fuels industry. Many different models of microalgae are characterized by their capability to produce high lipid content (De Carvalho et al., 2009; Ming et al., 2012), such as the chlorophyte Tetraselmis suecica (Montero et al., 2010), T. chuii (Mohammady, 2004), the haptophytes Pavlova lutheri and Isocrysis sp. (Brown et al., 1993), the eustigmatophyte Nannochloropsis salina (Mohammady, 2007; Mohammady and Fathy, 2007; Mohammady, 2008), the diatoms Thalassiosira weissflogii (Mann and Yentsch, 1989), Chaetoceros muelleri (McGinnis et al., 1997) and many cyanobacteria (Basova, 2005).
There are many ways for analyzing total lipids; gravimetric (Bligh and Dyer, 1959) and analytical methods (Nichols et al., 1988, 1989) are common examples. However, fluorescence spectrometric method using Nile Red stain was reported as a rapid technique for screening and comparing the lipid accumulated in different algal strains (Cooksey et al., 1987; Lee et al., 1998). Since the increased fluorescence intensity could be correlated with increased numbers of lipid droplets inside the algal cells (Cooksey et al., 1987) and corresponded to the distinct sites of lipid accumulation in microalgae (Lee et al., 1998). The regulation of algal lipid metabolism is greatly influenced by many factors such as cell growth phase (Mohammady, 2007; Chiu et al., 2009), nutrient limitation (Griffiths and Harrison, 2009; Ilavarasi et al., 2011) and environmental stresses (Guschina and Harwood, 2006). Cultivating microalgae in nitrogen deficient growth medium has been shown to accumulate lipid inside the algal cell (Elsey et al., 2007; Mohammady and Fathy, 2007; Wang et al., 2008; Pulz and Gross, 2004; El-Shoubaky et al., 2008) postulated that successful algal biotechnology mainly depends on choosing the right alga with relevant properties for specific culture conditions and products. Accordingly, optimizing a specific alga for producing economical biofuels in competition with the fossil fuels is the main challenge facing scientists.
It is well known that microalgae used in aquaculture industry are recommended as a feedstock for biofuel production due to their high lipid content. In this respect, the objective of this study was to identify the most promising microalga (cultivated in the experimental hatchery at the Marine Biological Laboratory, MA, USA) for biofuel production and how to justify between growth rate and maximum lipid production of the selected strain.
MATERIALS AND METHODS
Maintenance of microalgae: A collection of unialgal cultures was batch grown in the experimental hatchery at the Marine Biological Laboratory, MA, USA. Starter cultures were obtained from the NOAA Milford Laboratory in Milford Connecticut. The study was carried out on the following strains (followed by their accession number in the CCMP collection): Class Chlorophyceae:
| • | Tetraselmis chuii (CCMP884), 2- T. striata (CCMP), 3- T. sp. (CCMP961); Class Bacillariophyceae:1- Chaetoceros calcitrans (CCMP1315), 2- Chaetoceros muelleri (CCMP1425), 3- Thalassiosira weissflogii (CCMP1047); Class Haptophyceae: |
| • | Isocrysis sp. clone C. Iso (CCMP 463), 2- Iso. sp. (clone T. Iso.) (CCMP 1324) |
| • | Pavlova lutheri (CCMP1047) |
Growth conditions: Microalgae cultures were cultivated in 250 mL glass Erlenmeyer flasks using standard F2 media. Algae were incubated in a climatic chamber under continuous light conditions using fluorescent light tube providing a photosynthetic active radiation expressed in photon flux of 31 μmol. photons m-2 sec-1. The flasks held on an oscillating shaker (100 rpm) at 19±1°C.
Nitrogen-deficient growth medium: Tetraselmis chuii was cultivated in nitrogen-free F2 growth medium, for 15 days, under the same previous conditions.
Growth measurement: The cell density was measured daily with a spectrophotometer (Spectronic 20 Genesys) using 680 nm wavelength (Lee et al., 1998). The cell number was also counted using a haemocytometer. A correlation between the algal cell number and Optical Density (OD) was developed. However, results here are represented as cell number and the subsequent growth rate was calculated according to Robert (1979) as follows:
R = (3.322/t2-t1)x(log N2/N1) |
Where:
| 3.322 | = | Growth constant |
| t1 | = | Time at the beginning of the experiment |
| t2 | = | Time at the end of the experiment |
| N1 | = | Number of cells mL-1 culture at t1 |
| N2 | = | Number of cells mL-1 culture at t2 |
Preparation of nile red stain: According to Lee et al. (1998), a stock solution of Nile Red stain {9-diethlamino-5H-benzo(α)phenoxa-phenoxazine-5-one} was prepared by adding 2.5 mg of Nile Red to 100 mL of acetone. The solution is kept in a brown-colored bottle and stored in the dark at room temperature. Nile Red was added to the sterilized medium to give final concentration of 0.5 μg dye mL-1.
Fluorescence microscopy: Evaluation of the intracellular Nile Red staining was conducted using fluorescence microscopy. Images of the algal cultures were acquired 10 min after the stain was applied using a 40x/0.75 EC Plan-Neofluar phase contrast objective lens and an Axiocam MRm camera on a Zeiss Axio Imager Z.1 (Carl Zeiss Inc., Thornwood, NY). Three images were acquired at each field of view: a phase contrast image for overall cell morphology, a fluorescence image through Zeiss filter set 46HE (ex 500/25, bs 515LP, em 535/30) for Nile Red localization and a fluorescence image through Zeiss filter set 50 (ex 640/30, bs 660, em 690/50) for chlorophyll autofluorescence. Unstained negative controls were also imaged but no appreciable fluorescence was observed in the 46HE channel. The acquisition parameters and exposure times for all channels were kept constant between samples.
Further examination of T. chuii was performed to correlate cell size with Nile Red fluorescence intensity. Twenty two images were acquired containing a total of 31 cells. The images were analyzed for size and fluorescence intensity using the image analysis software, Image J (http://rsbweb.nih.gov/ij/). Cell size was determined using an intensity-based threshold of the chlorophyll autofluorescence images. An integrated density measurement of the corresponding intracellular Nile Red fluorescence was used as a proxy for lipid content.
Relative total lipid content: In this study, the method described by Lee et al. (1998) was used to determine the total lipid content in the investigated strains. Fluorescence was measured after staining with Nile Red stain on a PTi Quantamaster QM4 scanning spectrofluorometer (Photon Technology International, NJ, USA) using a 75 watt Xenon lamp and 5 nm excitation and emission slits. The sample was excited at a fixed wavelength of 480 nm and an emission spectrum was collected from 490-750 nm at 1 nm resolution and a scan rate of 1 nm sec-1. The relative total lipid content of the culture was calculated from the spectral emission profile after normalization of the chlorophyll peak at 680 nm. The contribution of fluorescence intensity from Nile red was obtained after subtraction of the autofluorescence of unstained algal cells (Lee et al., 1998; Liu et al., 2008).
Statistical analysis: T-test was used to evaluate differences between variables. A value of p≤0.05 was considered statistically significant.
RESULTS
Fluorescence microscopy: Figure 1 illustrates the localization of Nile Red fluorescence (shown in green) in the intracellular lipid droplets in relation to the chlorophyll (shown in red) of the nine studied microalgae grown at the end of their exponential growth phase.
Cell growth: The growth, as expressed in cell number of each individual, is represented in Fig. 2. Data showed that the growth progressively increased over time and the magnitude was more pronounced at the 12th day.
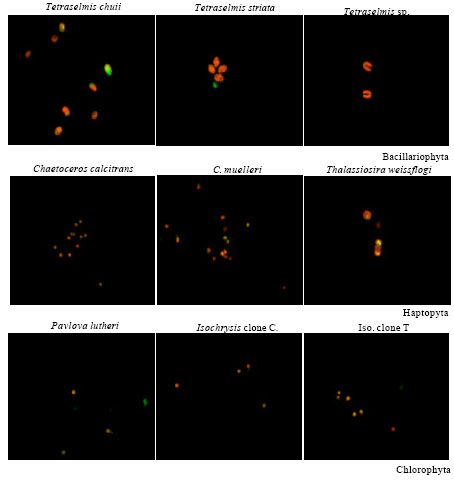 | |
| Fig. 1: | Fluorescence microscopic images of studied microalgae |
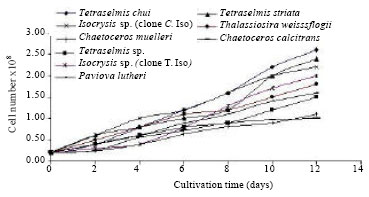 | |
| Fig. 2: | Batch growth of studied microalgae expressed as cell number over cultivation time |
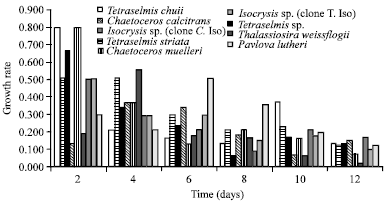 | |
| Fig. 3a: | Batch growth rate of each microalga for every two days of cultivation |
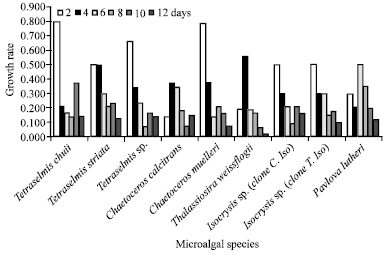 | |
| Fig. 3b: | Batch growth rate of studied microalgae over the cultivation time |
With respect to these results, T. chuii gave the highest cell number/ml over all the tested algae (2.6x108). While C. calcitrans showed the lowest cell number mL-1 (1.0x108). Furthermore, the chlorophytes showed the highest growth between the investigated algae. The highest cell number showed an increase by 13 times, 12 times and 11 times for T. chuii, T. sp. and T. striata, respectively, compared with the initial inoculums. On the other hand, the greatest cell number of C. calcitrans increased by only 5 times. Accordingly, the examined species could be arranged in a descending order as following: T. chuii, T. sp. T. striata, Iso. sp. (clone C. Iso), (clone T. Iso), T. weissflogii, C. muelleri, Pav. lutheri and C. calcitrans.
Growth rate: Figure 3a shows the growth rate of microalgae for each two days of the growth. While, Fig. 3b shows the growth rate of each individual over the cultivation time. At the first two days, the highest growth rate was recorded for both T. chuii and C. muelleri. Consequently, data here recommends T. chuii to be the best growing strain for the growth rate and dense cell number (p≤0.05) among all investigated microalgae. However, the least growth rate was recorded for C. calcitrans. Our results showed that the growth rate of the examined strains arranged in a descending order is; T. chuii, C. muelleri, T. sp., T. striata, Iso. sp. (clone C. Iso), (clone T. Iso), Pav. lutheri, T. weissflogii and C. calcitrans.
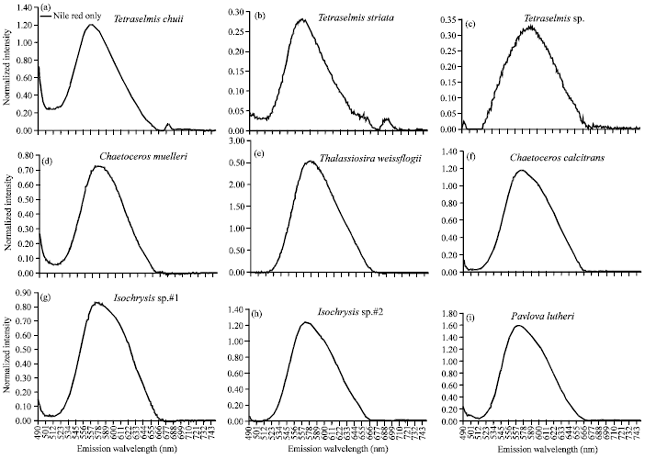 | |
| Fig. 4(a-i): | Fluorescence intensity of Nile Red as a function of lipid content of studied microalgae |
Fluorescence intensity of the lipid content: Since the available amount of microalgae for lipid analysis in this study was small, the Nile red fluorescence method was used to measure the content of lipid droplets. Screening of lipid production of microalgae using Nile Red staining has identified the examined strains to have capability to produce lipid within a range of previously published data. In this work, the corrected fluorescence spectrum was derived by subtracting the spectra before and after the Nile Red addition. Therefore, the intensity of corrected spectrum is corresponded to the intracellular lipid amount of each microalga. Figure 4 shows the fluorescence intensity corresponding to the lipid concentration in each species.
Relative lipid content: Data of relative lipid (the ratio of fluorescence intensity to the chl a content of each species (Fig. 5) shows a distinctive difference; the most promising strains for biofuel production are the bacillariophytes (p≤0.05), followed by haptophytes, with the exception of T. striata. However, the least lipid producer was T. chuii. Therefore, the relative lipid content of the microalgae arranged in a descending order is: C. calcitrans, T. weissflogii, C. muelleri, T. striata, Iso sp. (clone T. Iso), Pav. lutheri, Iso. sp. (clone C. Iso), T. sp., T. chuii.
Optimizing T. chuii for improving accumulation of lipid content: In this study, T. chuii has been recognized as the fastest increasing alga and best economically grown (because of its highest growth rate, shared with C. muelleri) as well among the examined species (p≤0.05).
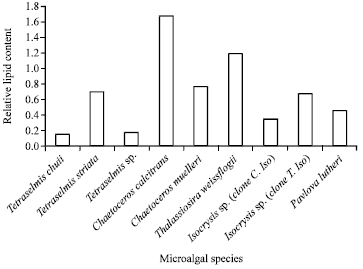 | |
| Fig. 5: | Relative lipid content of the studied microalgae |
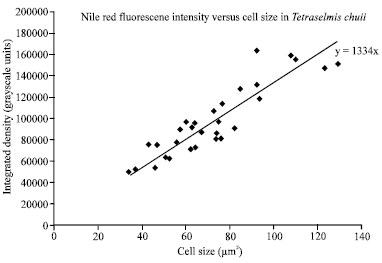 | |
| Fig. 6: | A linear correlation between the cell size of T. chuii and the fluorescence intensity of Nile Red as a function of lipid content |
However, the alga accumulates the least amount of lipid comparing to other strains. The photograph of the culture population of T. chuii using Differential Interference Contrast (DIC) microscopy showed a variation in its cell size. The more the algal cell increases in size, the more intense the emission from the Nile Red staining when observed with fluorescence microscopy. What’s more, a linear correlation between the algal cell size and the fluorescence intensity was demonstrated in Fig. 6, suggesting increased lipid accumulation in the bigger cells. For equality of the algal cell sizes, the cells were transferred into N-deficient growth medium for 15 days. Accordingly, the amount of relative lipid increased by about 35% compared to the controlled culture (Fig. 7).
DISCUSSION
The term microalgae describe a very diverse group of thousands of organisms with different morphological and metabolic properties. In this study, a variety of marine microalgae belonging to different classes were presented for analysis by fluorescence microscopy and spectrofluorometry and screened for their potential for maximum growth and lipid production.
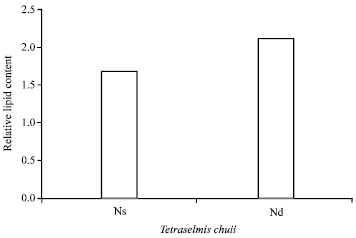 | |
| Fig. 7: | Relative lipid content of T. chuii grown in N-sufficient (Ns) and N-deficient (Nd) F2 growth media |
T. chuii, as the fastest growing alga among the investigated spp, was shown to maximize lipid production when grown under nitrogen deficient conditions. The fluorescence microscopic images presented here indicate that a distinct difference in lipid content was present in the cells of the investigated strains. In this research, microalgae were harvested at the end of exponential growth phase; a transition between highly metabolically active cells and stressed stationary cells. Under these conditions, the results of growth revealed that the best grown group is the chlorophytes followed by the diatoms and finally the haptophytes. A result similar to those recorded by Griffiths and Harrison (2009), who reported that Tetraselmis suecica is characterized by very high growth rate, followed by Nannochloropsis, Pavlova lutheri and Phaeodactylum tricornutum. In this study, T. chuii appeared with highest growth rate after the first two days of cultivation. A result reflects a significant economic value of this alga for biofuel production at the level of a commercial scale. However, the results revealed that T. chuii is the least lipid producer compared to the examined strains. Griffiths and Harrison (2009) stated that many high biomass producers’ microalgae are not high lipid producers. Unlikely, other previous studies screened many algal species for high lipid productivity and found the marine Nannochloropsis sp. (Mohammady and Fathy, 2007; Rodolfi et al., 2009; Doan and Obbard, 2010; Wiyarno et al., 2011) and Tetraselmis suecica (Rodolfi et al., 2009; Montero et al., 2010) to be particularly promising. In this paper, the relative lipid content of T. chuii was proportional to the cell size. A linear correlation between lipid content and cell size of this organism was confirmed. The linkage between our results and what was demonstrated by Cooksey et al. (1987) that increased fluorescence intensity, using Nile Red, could be correlated with increased numbers of lipid droplets inside the algal cells, suggesting increase lipid accumulation over aging in T. chuii. Previous studies stated that lipid accumulation could be increased from logarithmic growth to stationary growth phase (Mohammady and Fathy, 2007; Tonon et al., 2002; Chiu et al., 2009; Ilavarasi et al., 2011). Therefore, culture aging or senescence affects lipid content and composition in many microalgae ( Hu et al., 2008; Ilavarasi et al., 2011). The same conclusion was reported early by Collins and Kalinins (1969) with the green alga Chlorococcum macrostigma. However, lipid composition in the diatom Phaeodactylum tricornutum is not affected by age (Alonso et al., 2000).
Cooksey et al. (1987) demonstrated that the amount of chlorophyll is an indication of cell size. In this study, the ratio of chlorophyll fluorescence to Nile Red fluorescence was used to normalize the results to account for differences in cell size and age and allow detection of individual cells with unusually high lipid levels resulting from natural genetic variation.
Lipid production is partially regulated by algae cellular growth (Ilavarasi et al., 2011), which has been directly linked with nitrogen sufficiency (Sheehan et al., 1998). When, T. chuii was being transferred to N-deficient medium, the cells appeared uniform and attained its maximum size after 15 days. In addition, the total lipid content increase by about 35% compared to the controlled cells. A similar result has been observed in the lipid content of N-starved Isocrysis during various growth phases by (Solomon and Palumbo, 1987; Ilavarasi et al., 2011). From our opinion, the optimization of nitrogen in the growth medium is needed to balance between producing high lipid content while maintaining high growth productivity. In this regard, Illman et al. (2000) suggested that algae may need to be cultivated frequently in nitrogen deficient nutrient medium. While Solomon (1985) stated that algal cells respond as individuals to a lipid trigger rather than gradually increasing the lipid content of the entire culture. However, according Hu et al. (2008) and Ilavarasi et al. (2011), the age of batch culture mode of microalgae may or may not be associated with nutrient depletion, making it difficult to separate true aging effects from nutrient deficiency-induced effects on lipid metabolism. In members of Chlorophyta, nitrogen deficiency correlates with increased lipid content whereas the response of other taxa to nitrogen stress is more varied (Griffiths and Harrison, 2009).
CONCLUSION
Overall view of the present study showed that the growth characterization and lipid content of the examined algae species are within the previous published work. Nitrogen deficiency is enhancing lipid content of T chuii. The optimization of nitrogen in the growth medium is needed to balance between producing high lipid content while maintaining high growth productivity. However, further studies must be carried out on other taxa to understand the mechanism that controls the relationship between algal cell cycle and lipid production in relation to nitrogen deficiency. This should provide useful new information and will enable manipulation of selected algal strains that exhibit rapid growth and lipid accumulation simultaneously to ensure maximum sustainable biofuel production.
ACKNOWLEDGMENT
This study was supported by Lucy B. Lymann research award, Marine Biological Laboratory, Woods Hole, MA, USA. And this is acknowledged and greatly appreciated by the authors.
REFERENCES
- Abdeshahian, P., M.G. Dashti, M.S. Kalil and W.M.W. Yusoff, 2010. Production of biofuel using biomass as a sustainable biological resource. Biotechnology, 9: 274-282.
CrossRefDirect Link - Alonso, D.L., E.H. Belarbi, J.M. Fernandez-Sevilla, J. Rodrigues-Ruiz and E.M. Grima, 2000. Acyl lipid composition variation related to culture age and nitrogen concentration in continuous culture of the microalga Phaeodactylum tricornutum. Phytochemistry, 54: 461-471.
CrossRef - Bligh, E.G. and W.J. Dyer, 1959. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol., 37: 911-917.
CrossRefPubMedDirect Link - Chiu, S.Y., C.Y. Kao, M.T. Tsai, S.C. Ong, C.H. Chen and. C.S. Lin, 2009. Lipid accumulation and CO2 utilization of Nannochloropsis oculata in response to CO2 aeration. Bioresour. Technol., 100: 833-838.
CrossRefPubMedDirect Link - Ming, L.C., R. Nurliyana, A.B. Syah, M.N. Azizah, H.L. Sim and M.Y. Hirzun, 2012. Identification and biochemical composition of a green microalgae. Asian J. Biotechnol., (In press).
Direct Link - Chojnacka, K. and F.J. Marquez-Rocha, 2004. Kinetic and stoichiometric relationships of the energy and carbon metabolism in the culture of microalgae. Biotechnology, 3: 21-34.
CrossRefDirect Link - Cooksey, K.E., J.B. Guckert, S.A. Williams and P.R. Callis, 1987. Fluorometric determination of the neutral lipid content of microalgal cells using Nile red. J. Microbiol. Methods, 6: 333-345.
Direct Link - De Carvalho, P.R., M.C.G. Pita, E.P. Neto and C.X. De Mendonca, 2009. Efficiency of PUFAs incorporation from marine sources in yolk egg's laying hens. Int. J. Poult. Sci., 8: 603-614.
CrossRefDirect Link - Dayananda, C., R. Sarada, T.R. Shamala and G.A. Ravishankar, 2006. Influence of nitrogen sources on growth, hydrocarbon and fatty acid production by Botryococcus braunii. Asian J. Plant Sci., 5: 799-804.
CrossRefDirect Link - Doan, T.T.Y. and J.P. Obbard, 2010. Improved Nile Red staining of Nannochloropsis sp. J. Applied Phycol., 23: 895-901.
CrossRef - Elsey, D., D. Jameson, B. Raleigh and M.J. Cooney, 2007. Fluorescent measurement of microalgal neutral lipids. J. Microbiol. Methods, 68: 639-642.
CrossRefPubMedDirect Link - El-Shoubaky, G.A., A.M.Y. Moustafa and E.A.E. Salem, 2008. Comparative phytochemical investigation of beneficial essential fatty acids on a variety of marine seaweeds algae. Res. J. Phytochem., 2: 18-26.
CrossRefDirect Link - Jegannathan, K.R., E.S. Chan and P. Ravindra, 2011. Biotechnology in biofuels-A cleaner technology. J. Applied Sci., 11: 2421-2425.
CrossRef - Griffiths, M.J. and S.T.L. Harrison, 2009. Lipid productivity as a key characteristic for choosing algal species for biodiesel production. J. Applied Phycol., 21: 493-507.
CrossRefDirect Link - Guschina, I.A. and J.L. Harwood, 2006. Lipids and lipid metabolism in eukaryotic algae. Prog. Lipid Res., 45: 160-186.
CrossRefPubMedDirect Link - Hu, Q., M. Sommerfeld, E. Jarvis, M. Ghirardi, M. Posewitz, M. Seibert and A. Darzins, 2008. Microalgal triacylglycerols as feedstocks for biofuel production: Perspectives and advances. Plant J., 54: 621-639.
CrossRefPubMedDirect Link - Ilavarasi, A., D. Mubarakali, R. Praveenkumar, E. Baldev and N. Thajuddin, 2011. Optimization of various growth media to freshwater microalgae for biomass production. Biotechnology, 10: 540-545.
CrossRefDirect Link - Kotchoni, S.O. and E.W. Gachomo, 2008. Biofuel production: A promising alternative energy for environmental cleanup and fuelling through renewable resources. J. Boil. Sci., 8: 693-701.
CrossRefDirect Link - Lee, S.J., B.D. Yoon and H.M. Oh, 1998. Rapid method for the determination of lipid from the green alga Botryococcus braunii. Biotechnol. Tech., 12: 553-556.
CrossRefDirect Link - McGinnis, K.M., T.A. Dempster and M.R. Sommerfeld, 1997. Characterization of the growth and lipid content of the diatom Chaetoceros muelleri. J. Applied Phycol., 9: 19-24.
CrossRefDirect Link - Mohammady, N.G., 2004. Total, free and conjugated sterolic forms in three microalgae used in mariculture. Z. Naturforsch., 59: 619-624.
PubMed - Mohammady, N.G.E., 2007. Different light spectral qualities influence sterol pool in Porphyridium cruentum (Rhodophyta). Am. J. Plant Physiol., 2: 115-121.
CrossRefDirect Link - Mohammady, N.G.E. and A.A. Fathy, 2007. Humic acid mitigates viability reduction, lipids and fatty acids of Dunaliella salina and Nannochloropsis salina grown under nickel stress. Int. J. Bot., 3: 64-70.
CrossRefDirect Link - Montero, M., M. Aristizabal and G. Reina, 2010. Isolation of high-lipid content strains of the marine microalga Tetraselmis suecica for biodiesel productionby flow cytometry and single-cell sorting. J. Applied Phycol., 23: 1053-1057.
CrossRef - Nichols, P.D., A.C. Palmisano, J.K. Volkman, G.A. Smith and D.C. White, 1989. Changes in the lipid composition of Antarctic Sea Ice diatom communities during a spring bloom: an indication of community physiological status. Antarct. Sci., 1: 133-140.
CrossRefDirect Link - Nichols, P.D., J.K. Volkman, A.C. Palmisano, G.A. Smith and D.C. White, 1988. Occurrence of an isoprenoid C25 di-unsaturated alkene and high neutral lipid content in Antarctic Sea Ice diatom communities. J. Phycol., 24: 90-96.
CrossRef - Pulz, O. and W. Gross, 2004. Valuable products from biotechnology of microalgae. Applied Microbiol. Biotechnol., 65: 635-648.
CrossRefDirect Link - Rodolfi, L., G.C. Zittelli, N. Bassi, G. Padovani, N. Biondi, G. Bonini and M.R. Tredici, 2009. Microalgae for oil: Strain selection, induction of lipid synthesis and outdoor mass cultivation in a low-cost photobioreactor. Biotechnol. Bioeng., 102: 100-112.
CrossRefPubMedDirect Link - Tonon, T., D. Harvey, T. Larson and I. Graham, 2002. Long chain polyunsaturated fatty acid production and partitioning to triacylglycerols in four microalgae. Phytochemistry, 61: 15-24.
CrossRef - Wang, B., Y. Li, N. Wu and C.Q. Lan, 2008. CO2 bio-mitigation using microalgae. Applied Microbiol. Biotechnol., 79: 707-718.
CrossRef - Wiyarno, B., R.M. Yunus and M. Mel, 2011. Extraction of algae oil from Nannocloropsis sp.: A study of soxhlet and ultrasonic-assisted extractions. J. Applied Sci., 11: 3607-3612.
CrossRefDirect Link - Illman, A.M., A.H. Scragg and S.W. Shales, 2000. Increase in Chlorella strains calorific values when grown in low nitrogen medium. Enzyme Microb. Technol., 27: 631-635.
CrossRefDirect Link