
Selvaraj Venkatesan
Cavinkare Research Center, Chennai, India
Sankar Jayaganesh
UPASI Tea Research Foundation, Nirar Dam BPO, Valparai-642 127, Tamil Nadu, India
Research Journal of Phytochemistry
Year: 2010 | Volume: 4 | Issue: 2 | Page No.: 67-77







ABSTRACT
The excess potassium (K) application to the soil reduces the availability of magnesium (Mg) and vice versa due to antagonism. A pot culture experiment was carried out in which pots were filled with 4 kg of soil to which magnesium sulphate was added externally so as to have 100, 250, 500, 750, 1000, 2000, 5000 and 10000 mg of Mg kg-1 of soil. Excess magnesium has induced some toxicity symptoms like development of coppery color along the marginal veins at the initial stage. The mid rib region was also slightly affected. Extensive coppery color developed all over the leaf surface and defoliation of leaf occurred during the final staged of toxicity. The plant, which received 10000 mg of Mg2+ kg-1 died on 20th day after imposing treatments and the plants, which received 5000 mg of Mg2+ kg-1, died on 45th day. The surviving plants were uprooted and separated into leaves, stem, root and soil, on which chemical analysis was carried out. Surprisingly, at any given treatment the accumulation of Mg in root was higher than that of leaf and stem. This study confirmed the antagonism existing between K and Mg and synergism existing between the P and Mg. The amino acid content increased in tea leaf up to 1000 mg treatment and then gradually decreased.
PDF Abstract XML References Citation
How to cite this article
Selvaraj Venkatesan and Sankar Jayaganesh, 2010. Characterisation of Magnesium Toxicity, its Influence on Amino Acid Synthesis Pathway and Biochemical Parameters of Tea. Research Journal of Phytochemistry, 4: 67-77.
URL: https://scialert.net/abstract/?doi=rjphyto.2010.67.77
URL: https://scialert.net/abstract/?doi=rjphyto.2010.67.77
INTRODUCTION
Magnesium is an activator of several enzymes involved in carbohydrate metabolism and synthesis of nucleic acids. Availability of magnesium in soils considered to be important because it is the only metal existing in chlorophyll molecule. The magnesium content of tea harvestable shoots varies between 0.2 and 0.3%. Although, the magnesium removal through leaf harvesting is 6 to 8 times lesser than that of potassium, the higher potassium input strengthens the antagonism leading to magnesium deficiency (Nayyar, 2002; Venkatesan, 2006), which started appearing in many parts of the South Indian tea gardens. The correction is done through foliar application of mixture of zinc sulphate, magnesium sulphate, manganese sulphate, boric acid and naphthalene acetic acid (Verma and Palani, 1997). However, potassium rate cannot be reduced because the aim was to have sustainability in tea productivity. About 150 to 300 kg of K2O is added every year to overcome potassium deficiency, while only very little quantity of magnesium is supplied to the soil. Hence, our scientific department has recommended the soil application of MgSO4 recently (Verma and Palani, 1997; Venkatesan, 2006). It is necessary , to be well informed about the critical and toxicity levels of magnesium and the typical symptoms on tea leaves. While the magnesium deficiency in tea is well recorded and recognized, there is no information on its toxicity symptoms in tea, particularly under South Indian conditions. Although, magnesium is very much needed for tea growth, higher concentration would retard growth and development of plants by interfering with certain metabolic process. The toxic effect of magnesium on other plants, mainly serpentine soil has been reported by many researchers (Brady et al., 2005; Loganathan et al., 2005; Kayama et al., 2005). Many researchers have proved that the soil applied magnesium improved the free amino acid content and quality of made tea (Ruan et al., 1999; Ma et al., 2005). However, this is the first attempt to find out the influence of magnesium on amino acid synthesis pathway in tea under South Indian condition. The main objectives of the present study were: to determine the distribution and accumulation of soil applied magnesium; to fix the critical toxic limit of the magnesium; to find out the interaction of magnesium with other essential elements in plant and soil and to document its influence on biochemical parameters.
MATERIALS AND METHODS
Pot grown UPASI-9 tea plants (one year old) were used for this study with nine treatments including an untreated control. The pots were filled with 4 kg of soil to which magnesium sulphate was added externally so as to have 100, 250, 500, 750, 1000, 2000, 5000 and 10000 mg of Mg kg-1 of soil. The experiment was conducted in triplicate manner during the year of 2008-2009 at UPASI Tea Experimental Farm. A moisture meter (theta meter type HH1) was used to maintain the soil moisture at 20%. The plants were monitored everyday and the visual toxicity symptoms were recorded. The experiment was continued for a period of 150 days and the plants were uprooted on 150th day and were separated into roots, stem and leaves. The soil samples were air dried and passed through 2 mm sieve (Klose and Tabatabai, 2000). Separated vegetative parts were oven dried at 60°C and homogenized with mortar and pestle. About 1 g from every plant part was digested with HNO3/HClO4 mixture (Haunter et al., 1987) and analysed for Fe, Zn, Mn, Ca, Mg, K and P (Bhargava and Raghupathi, 2001) using ion exchange chromatography (Dionex ICS 2500). The metal standards used in this study were traceable to NIST (National Institute for Standards and Technology).
Glutamate Synthase Assay
Exactly, 0.5 g of plant material was homogenized with 5 mL of 100 mM phosphate buffer (pH 7.5) containing 1 mM disodium EDTA, 1 mM dithioerythrietol and 1% polyvinyl pyrolidone. The homogenate was centrifuged at 10,000 rpm for 30 min at 4°C. The supernatant was collected and used as enzyme source. The activity was assayed spectrophotometrically as described by Van de casteele et al. (1975). The reaction mixture consisted of 1 mL of 5 mM glutamine, 1 mL of 5 mM 2-oxoglutarate, 1 mL of 0.25 mM NADPH, 0.2 mL of enzyme extract and 1.8 mL of 50 mM tris HCl buffer. The absorbance was measured at 340 nm.
Amino Transferase Enzymes
About 1 g of plant tissue was homogenized with 0.2 M potassium phosphate buffer (pH 7.5). The homogenate was centrifuged at 25,000 g for 15 min to get the enzyme fraction. To the treatment about 0.5 mL of aspartic acid substrate solution was added and warmed up in a water bath at 37°C for 3 min. Then, 0.2 mL of enzyme extract was added and incubated at 37°C for 60 min and then 0.5 mL of dinitrophenylhydrazine solution was added. To the control about 0.5 mL of substrate, 0.5 mL of DNPH and 0.1 mL enzyme extract were added. The mixture was kept at room temperature for 20 min and then 0.4 N NaOH was added. After 10 min the absorbance was read at 510 nm (Bergmeyer and Burnt, 1974).
Nitrate Reductase Activity
The leaf samples were washed with tap water and finally with distilled water to remove the foreign materials. Immediately after washing, the leaf materials were blotted dry before being assayed. The NR activity was assayed in vivo as described by Sarmah et al. (1987), where potassium nitrate was used as substrate. A sliced (1 cm) sample (200 mg) of leaf suspended in a thunberg tube containing 5 mL of medium comprising 0.1 M phosphate buffer at pH 7.5, 0.02 M KNO3 6% (w/v), purified polyclar-AT and two drops of chloramphenicol (0.5 mg mL-1). The tubes were evacuated for 1 min and then vacuum was released. The amount of nitrite formed during 4 h of incubation period was estimated using colorimetric technique as described by Bremner (1965) at 550 nm using UV-Visible Spectrophotometer (GBC make, Model: 918). The activity was expressed in micromoles of nitrite formed g-1 fresh weight of leaf h-1 (μmol NO2 g-1 FW h-1).
Biochemical Parameters
Leaf samples of known quantity weighed in a mortar was ground with 25 mL of methanol. It was filtered and made up to 50 mL in a volumetric flask-using methanol. This solution was diluted five times with methanol and used for the estimation of chlorophyll A, B and carotenoids (Welburn, 1994) by measuring the absorbance at 470, 653 and 666 nm, respectively using UV-VIS spectrophotometer (GBC 918).
About 0.5 g of leaf sample was weighed and ground with ethyl alcohol. The contents were filtered and the filtrate was made up to 50 mL with ethyl alcohol. This alcoholic extract was used for the estimation of polyphenols (Dev Choudhury and Goswami, 1983) and amino acids (Moore and Stein, 1948) and the absorbance was recorded at 700 and 570 nm, respectively using UV-VIS Spectrophotometer (GBC 918).
Statistical analysis was carried out by the standard method (Gomez and Gomez, 1984).
RESULTS AND DISCUSSION
Generally, magnesium toxicity stress is more difficult to develop. When, Mg2+ is plentiful the plants generally take up the ion and store it (Stelzer et al., 1990). However, if this is followed by drought then ionic concentrations within the cell can increase dramatically. High cytoplasmic Mg2+ concentrations block a K+ channel in the inner envelope membrane of the chloroplast, in turn inhibiting the removal of H+ ions from the chloroplast stroma. This leads to an acidification of the stroma that inactivates key enzymes in carbon fixation, which leads to the production of oxygen free radicals in the chloroplast and then cause oxidative damage (Wu et al., 1991).
The plants, which received 10000 mg of Mg2+ kg-1 of soil showed toxicity symptoms on 5th day after imposing treatments. The plants grown in the soil containing 5000 mg of Mg2+ kg-1 soil showed toxicity symptoms on 12th day after imposing treatments. Coppery color developed along the marginal veins at the initial stage and subsequently the mid rib region was slightly affected (Fig. 1a, b). Finally, extensive coppery color had appeared all over the leaf surface and defoliated. The plant, which received 10000 mg of Mg2+ kg-1, died on 20th day after imposing treatments and those received 5000 mg of Mg2+ kg-1 died on 45th day.
 | |
| Fig. 1: | Magnesium toxicity symptoms (a) initial stage and (b) final stage |
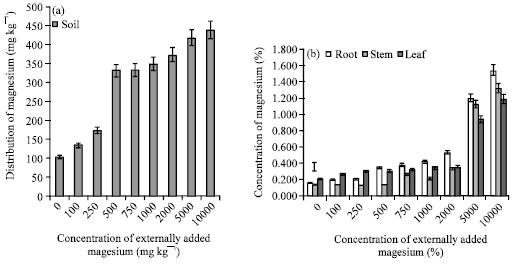 | |
| Fig. 2: | (a) Levels of available magnesium in soil after 150 days of external addition. (b) absorption and distribution of magnesium to various plant parts, The error bars represent the relative standard deviation |
Throughout the study, the bottom most leaves of the plant were showing toxicity symptoms. The soil pH increased from 4.3 to 5.2 when, magnesium concentration was increased. Toxicity symptoms appeared a little later (120th and 140th) on the plants established with 2000 and 1000 mg Mg kg-1 of soil, respectively.
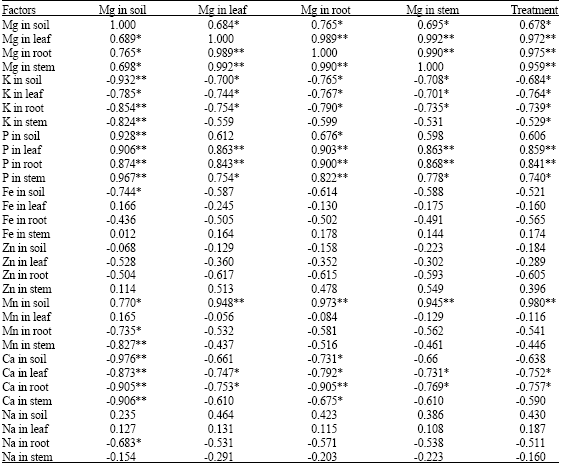
The magnesium content estimated in soil and plant parts are given in Fig. 2a and b, at any given treatment the accumulation of Mg in root was higher than that of leaf and stem. Similar kind of observation was reported by Brady et al. (2005). Statistical analysis among the externally added magnesium and magnesium content of soil, root, stem and leaves exhibited positive and significant correlation coefficients (Table 1). This indicates that the tea plants could accumulate higher magnesium when supplied in excess quantity.
| Table 1: | Correlation co-efficient (r) worked out between magnesium and other elements in various parts of tea plant |
 | |
*Significant at 5% level; **Significant at 1% level | |
While no toxicity symptoms were recorded till 140th day 1000 mg kg-1 treatment, the plants that received 2000 mg kg-1 showed mild toxicity symptoms on 120th day. Hence, the magnesium content of the plant parts that received 2000 mg kg-1 was taken as the critical toxicity limit. Accordingly was fixed as 3435, 3260 and 5285 mg kg-1 for leaf, stem and root, respectively.
Among all the elements studied, influence of Mg was very much pronounced on P content of soil, which increased when the magnesium concentration was increased (Fig. 3a, b). The study conducted elsewhere indicated that magnesium forms a soluble complex with P leading to a drastic increase in soil P (Franco and Coutinho, 2004). Both, the elements may be taken up in a similar way and Mg2+ may act as a phosphatic carrier (Kirkby and Mengal, 1976). This is evidenced by the positive correlation coefficient obtained between P and Mg content of soil (r = 0.928; p = 0.01). However, P estimated in various plant parts also showed the similar kind of trend. This study confirmed the synergism existing between magnesium and phosphorus (Verma, 1995; Verma and Palani, 1997; Franco and Coutinho, 2004).
Already many researchers proqved the antagonism existing between Mg and K in tea soils of South India (Verma and Palani, 1997; Ruan et al., 1998). In this study, also the K content of soil drastically decreased when the concentration of externally added magnesium increased (Fig. 4a, b). The higher amount of magnesium input prevents the potassium uptake in the plant. High cytoplasmic Mg2+ concentrations block the K+ channel in the inner envelope membrane of the chloroplast, in turn inhibiting the removal of H+ ions from the chloroplast stroma (Stelzer et al., 1990; Ding et al., 2006).
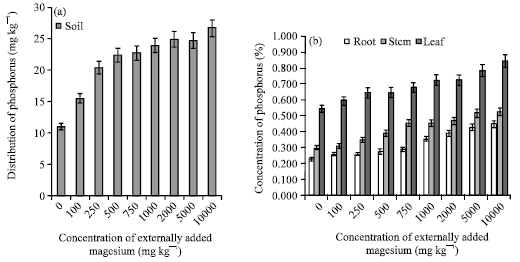 | |
| Fig. 3: | Influence of externally added magnesium (a) on distribution of P in soils (b) on P uptake by various plant parts. The error bars represent the relative standard deviation |
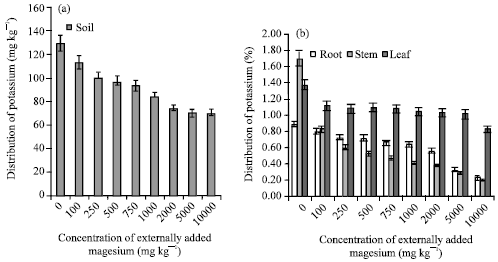 | |
| Fig. 4: | Influence of externally added magnesium (a) on distribution of K in soils (b) on K uptake by various plant parts. The error bars represent the relative standard deviation |
This study proved the antagonistic behavior obtained in the stem and root portion also. This is evidenced by the negative correlation coefficient obtained between K and Mg content of soil, leaf, root and stem (r = -0.932, -0.785, -0.854 and -0.824, respectively; p = 0.01).
The calcium content of soil drastically decreased with increase in concentration of externally added magnesium (Fig. 5a, b). The higher magnesium input prevents the calcium uptake by plant, because both are divalent cations with similar radius to each other (Cofie and Pleysier, 2004; Cornfield and Pollard, 2006).
The manganese content of soil increased with increase in concentration of externally added magnesium (Fig. 6a, b), but such a trend did not appear in the plant parts like root, stem and leaf. Manganese shows the properties of both the alkaline earth cations (Mg2+ and Ca2+) and heavy metals (Zn and Fe) and higher magnesium input reported to increase the bivalent form of manganese to the soil (Marschner, 1995).
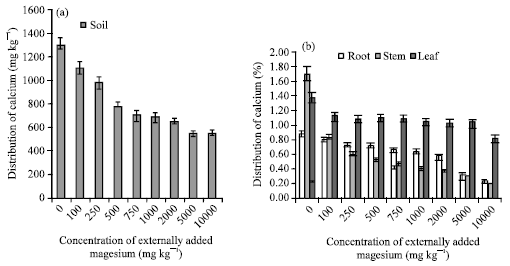 | |
| Fig. 5: | Influence of externally added magnesium (a) on distribution of Ca in soils (b) on Ca uptake by various plant parts. The error bars represent the relative standard deviation |
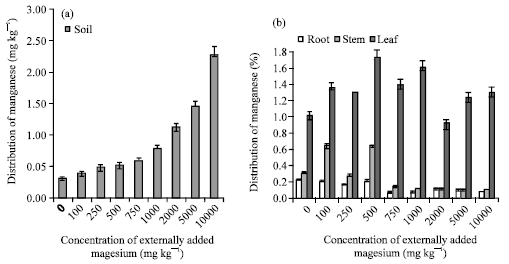 | |
| Fig. 6: | Influence of externally added magnesium (a) on distribution of Mn in soils (b) on Mn uptake by various plant parts. The error bars represent the relative standard deviation |
Other nutrients like zinc, sodium and iron are negatively correlated with externally applied magnesium. However, the correlation coefficients worked out between externally added magnesium and above nutrients were non-significant (Table 1).
Impact of Externally Added Magnesium on Biochemical Parameters
The amino acid content increased in tea leaf up to 1000 mg Mg kg-1 soil treatment and then gradually decreased (Table 2). This indicates that the synthesis and transport of amino acids in tea plants were improved by adequate supply of magnesium; however, the excess supplement of magnesium decreased the amino acid synthesis and bindered transport pathway (Ma et al., 2005). According to Ruan et al. (1998), magnesium application improved the amino acid content in tea leaves, apart from stimulating the nitrogen metabolism.
| Table 2: | Influence of externally added magnesium on amino acid synthesis pathway and biochemical parameters in tea plant |
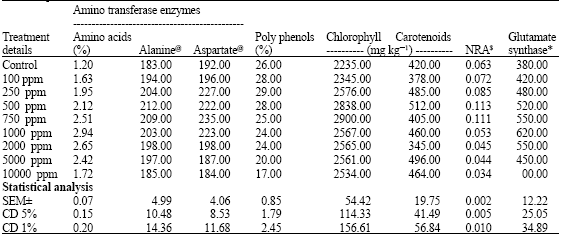 | |
@: μmole of pyruvate formed/min/g of fresh weight of leaf; $: μmol NO2 formed g-1 FW h-1; *: Nano mole of NADPH formed/min/g of fresh weight of leaf’ SEM±: Standard error mean; CD: Critical difference | |
Polyphenols content in tea leaves increased up to 250 mg Mg2+ kg-1 treatment and then drastically decreased. This is due to the inhibitory effect of magnesium on the synthesis of phenolic compounds. The excess amount of magnesium caused the less carbon skeletons supplied due to enhanced nitrogen metabolism (Ruan, 2005; Berkowitz and Wu, 1993).
In photosynthetic organisms Mg2+ has the additional vital role of being the coordinating ion in the chlorophyll molecule. This role was discovered by R. M. Willstätter, who received the Nobel Prize for Chemistry in 1915 for the purification and finding the structure of chlorophyll. Chlorophyll is a complex molecule possessing a central Mg atom, C20H39 side chain known as phytol and a porphyrin ring which is the main skeleton. The externally added magnesium increased the chlorophyll content of tea leaves up to 750 mg Mg2+ kg-1 and then decreased sharply (Huber and Maury, 1980; Dorenstouter et al., 1985). Carotenoids also showed the same trend as that of chlorophyll which could be due to the equivalent synthetic activity of pigments (Wilkinson and Ohki, 1988).
Influence of Magnesium on Certain Enzyme Activity
The NR activity increased up to 750 mg Mg2+ kg-1 and then gradually decreased. According to Ruan et al. (1998), the activity of NR has direct positive correlation with free amino acid content because the former is responsible for the nitrogen assimilation. In present studies, also we found that the amino acid content varied in accordance with NR activity. In other words, both NR activity and amino acid contents followed the same trend due to magnesium addition.
The amino transferease enzyme catalyses the following reaction in which the 2-oxoglutarate and aspartate/alanine are the final products.
Both aspartate and alanine amino transferease enzymes increased up to 750 mg Mg2+ kg-1 treatment and then gradually decreased. The externally added magnesium increased the formation of 2-oxoglutarate and simultaneously another product aspartate/alanine was increased (Temple et al., 1998; Kiss et al., 2004). Because, the influence of magnesium increased the protein synthesis and increased the accumulation of soluble organic nitrogen, nitrate nitrogen and amide forms of nitrogen in plant parts. Finally, the soluble amide forms of nitrogen converted into plant available form of amino acids.
The glutamate synthase is a unique enzyme playing important role in nitrogen metabolism. It catalyses the following reaction, where two moles of glutamates are formed (Kiss et al., 2004).
The activity of glutamate synthase increased up to 1000 mg Mg2+ kg-1 and then decreased gradually (Ma et al., 2005). The trend was almost similar to that of free amino acid content tea leaves.
CONCLUSION
The soil applied magnesium up to 2000 mg Mg kg-1 soil did not show severe toxicity symptoms in the tea plants. This study confirmed the antagonism existing between the K and Mg and synergism existing between the P and Mg. The amino acid content increased in tea leaf up to 1000 mg Mg kg-1 soil treatment and then gradually decreased. Similar kind of trend observed in amino transferease enzyme and glutamate synthase indicates that externally applied magnesium influences the amino acid synthesis pathway. The major achievement of this study is that the documentation of critical toxicity levels of magnesium, which was 3435, 3260 and 5285 mg kg-1 in leaf, stem and root of tea plant, respectively. Since, soil applied magnesium up to 2000 mg Mg kg-1 soil did not lead to Mg toxicity. It can be concluded that the soil application of magnesium sulphate along with NK application is a viable one.
ACKNOWLEDGMENTS
The authors are thankful to Dr. N. Muraleedharan, Director, UPASI Tea Research Foundation for his constant encouragement throughout the study. Our sincere thanks are also due to Mr. N. Palani, for critically evaluating this manuscript.
REFERENCES
- Bergmeyer, H.U. and E. Burnt, 1974. Methods of Enzymatic Analysis. 3rd Edn., Academic Press, New York, pp: 735.
Direct Link - Berkowitz, G.A. and W. Wu, 1993. Magnesium, potassium flux and photosynthesis. Magnesium Res., 6: 257-265.
PubMedDirect Link - Brady, K.U., A.R. Kruckeberg and H. Bradshaw, 2005. Evolutionary ecology of plant adaptation to serpentine soils. Annu. Rev. Ecol. Evol. Syst., 36: 243-266.
CrossRefDirect Link - Cofie, O.O. and J. Pleysier, 2004. Ion exchange involving potassium-calcium and magnesium-calcium in soil and organic matter fractions. Commun. Soil Sci. Plant Anal., 35: 2417-2431.
CrossRefDirect Link - Cornfield, A.H. and A.G. Pollard, 2006. The relative rates of release of potassium, calcium and magnesium from soils during electrodialysis. J. Sci. Food Agric., 3: 613-615.
CrossRefDirect Link - Ding, Y., W. Luo and G. Xu, 2006. Characterisation of magnesium nutrition and interaction of magnesium and potassium in rice. Ann. Applied Biol., 149: 111-123.
Direct Link - Dorenstouter, H., G.A. Pieters and G.R. Findenegg, 1985. Distribution of magnesium between chloroplhyll and other photosynthetic functions in magnesium deficient sun and shade leaves of poplar. J. Plant Nutr., 8: 1089-1101.
CrossRefDirect Link - Franco, H.C.J. and E.L.M. Coutinho, 2004. Evaluation of the agronomic effectiveness of phosphorus sources making use of different methods. Cientifica, 32: 58-64.
Direct Link - Huber, S.C. and W.J. Maury, 1980. Effects of magnesium on intact chloroplasts. Plant Physiol., 65: 350-354.
Direct Link - Kayama, M., A.M. Quoreshi, S. Uemura and T.A. Koike, 2005. Differences in growth characteristics and dynamics of elements absorbed in seedlings of three spruce species raised on serpentine soil in Northern Japan. Ann. Bot., 95: 661-672.
CrossRefDirect Link - Kirkby, E.A. and K. Mengal, 1976. The role of magnesium in plant nutrition. Z. Pflanzern. Bodenk., 139: 209-222.
CrossRef - Klose, S. and M.A. Tabatabai, 2000. Urease activity of microbial biomass in soils as affected by cropping systems. Biol. Fert. Soils, 31: 191-199.
CrossRef - Loganathan, P., A.D. Mitchell, J.A. Hanly and T.M. Aye, 2005. An improved procedure for determining magnesium fertiliser dissolution in field soils. Commun. Soil Sci. Plant Anal., 36: 2121-2128.
CrossRefDirect Link - Ma, L., J.Y. Ruan, Y. Yang, W. Han and Y. Shi, 2005. Effect of magnesium nutrition of the formation and transport of free amino acids in tea plants. Proceedings of International Symposium on Innovation in Tea Science and Sustainable Development in Tea Industry, Nov. 10-15, Tea Research Institute, Chinese Academy of Agricultural Sciences, Hangzhou, China, pp: 151-158.
- Ruan, J.Y., X. Wu and R. Hardter, 1999. Effect of potassium and magnesium nutrition on the quality components of different types of tea. J. Sci. Food Agric., 79: 47-52.
Direct Link - Ruan, J.Y., X. Wu, Y. Ye and R. Hardter, 1998. Effect of potassium, magnesium and sulphur applied in different forms of fertilisers on free amino acid content in leaves of tea (Camellia sinensis L.). J. Sci. Food Agric., 76: 389-396.
Direct Link - Kiss, S.A., E. Stefanovits-Banyai and M. Takacs-Hajos, 2004. Magnesium-content of rhizobium nodules in different plants: The importance of magnesium in nitrogen-fixation of nodules. J. Am. College Nutr., 23: 751-753.
Direct Link - Temple, S.J., C.P. Vance and J.S. Gantt, 1998. Glutamate synthase and nitrogen assimilation. Trends Plant Sci., 3: 51-56.
CrossRefDirect Link - Wellburn, A.R., 1994. The spectral determination of chlorophyll a and chlorophyll b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol., 144: 307-313.
Direct Link - Wilkinson, R.E. and K. Ohki, 1988. Influence of manganese deficiency and toxicity on isoprenoid syntheses. Plant Physiol., 87: 841-846.
PubMedDirect Link - Wu, W., J. Peters and G.A. Berkowitz, 1991. Surface charge-mediated effects of Mg2+ on K+ flux across the chloroplast envelope membrane are associated with the regulation of stromal pH and photosynthesis. Plant Physiol., 97: 580-587.
Direct Link - VandeCasteele, J.P., J. Lemal and M. Coudest, 1975. Pathways and regulation of glutamate synthesis in a Corynebacteriurn sp. overproducing glutamate. J. Gen. Microbiol., 90: 178-180.
PubMedDirect Link