
Rakesh Singh Sengar
College of Biotechnology, Sardar Vallabh Bhai Patel University of Agriculture and Technology,Meerut-250110, India
Madhu Gautam
Institute of Management Studies, C-238, Bulandshahr Road, Indl, Area, Lal Quan, G.T. Road, Ghaziabad-201009, India
Sanjay Kumar Garg
Department of Plant Science, M.J.P. Rohilkhand University, Bareilly-243006, India
Reshu Chaudhary
College of Biotechnology, Sardar Vallabh Bhai Patel University of Agriculture and Technology,Meerut-250110, India
Kalpana Sengar
College of Biotechnology, Sardar Vallabh Bhai Patel University of Agriculture and Technology,Meerut-250110, India
Research Journal of Phytochemistry
Year: 2008 | Volume: 2 | Issue: 2 | Page No.: 61-68







ABSTRACT
Seedling of mungbean (Vigna radiate) were raised in the pots to evaluate the effect of lead on growth, chlorophyll and nitrate reductase activity. The plants grown with increasing level of lead were reduced in germination in and growth compared to control plants. Chlorophyll content, carotenoids, soluble protein and organic nitrogen contents remained almost unaffected at lower concentration of lead tested. Enzyme activity in vitro and in vivo in excised leaves showed marked increase at lower concentration of lead. Chlorophyll carotenoids soluble protein and organic nitrogen contents remained almost unaffected at lower concentrations of lead tested. Thus lead shows the inhibition or reduction in growth and germination at higher concentration thus leads the decrease in chlorophyll level also. Inhibition in enzyme activity (in vitro and in vivo) also reported in excised and intact mungbean in presence of lead. It seems that nitrate reductase has a different response towards lead pollution in which is more tolerant to lead pollution Vigna radiata.
PDF Abstract XML References Citation
How to cite this article
Rakesh Singh Sengar, Madhu Gautam, Sanjay Kumar Garg, Reshu Chaudhary and Kalpana Sengar, 2008. Effect of Lead on Seed Germination, Seedling Growth, Chlorophyll Content and Nitrate Reductase Activity in Mung Bean (Vigna radiata). Research Journal of Phytochemistry, 2: 61-68.
URL: https://scialert.net/abstract/?doi=rjphyto.2008.61.68
URL: https://scialert.net/abstract/?doi=rjphyto.2008.61.68
INTRODUCTION
Heavy metal pollution of the environment which is increasing day by day has exerted deleterious effects on plants and consequently to man and animals through the food chain (Jones et al., 1973). Although lead is a non-essential element, plants can absorb it from soil, water and air through their roots and leaves (Zimdahl and Koeppe, 1977). The possible adverse effects of heavy metal pollution and their phytotoxic effects have been reported by Angelone and Bini (1992), Antonovies et al. (1971), Broyer et al. (1972), Chaney and Ryan (1994), Chila and Takahashi (1977), Davies (1995), de Abreu et al. (1998), Godzik (1993), Johnson and Eaton (1980), Kumar et al. (1995), Leblova et al. (1986), Laxen and Harrison (1977), Lee et al. (1998), Merakchiuska et al. (1976), Paivoke (2002) and Yang et al. (2000). Inhibitory effect of lead on photosynthesis or photosynthetic components have been reported in soybean (Bazzaz et al., 1975), corn (Bazzaz et al., 1975; Carlson et al., 1975), Zea mays (Eun et al., 2000; Stefanov et al., 1995), spruce (Keller and Zuber, 1970; Marschner et al., 1996), barley (Stiborova et al., 1987), maize leaves (Parekh and Puranik, 1992; Sinha et al., 1988a; Stiborova et al.,1987), maize seedlings (Iqbal and Mushtaq, 1987; Stiborova et al., 1987) and maize roots (Seregin et al., 2004). Inhibition of respiration in corn root tip (Koeppe, 1977; Miller and Koeppe, 1971), accumulation of lead in Indian mustard by soil applied chelating agents (Blaylock et al., 1997) and lead uptake in Raphanes sativua (Lane and Martin, 1977) has also been reported. The metal is known to inhibit protein content (Jana and Choudhuri, 1984) and chlorophyll in maize (Parekh and Puranik, 1992; Sinha et al., 1994), mung bean (Prassad and Prassad, 1987), Pisum sativum (Sengar and Pandey, 1996), nitrate reductase activity Pea leaves (Sinha et al., 1988b) and submerged aquatic plants (Jana and Choudhuri, 1984) have also been reported. The sole objective of the article under references is to exam the possibilities of physiological and biochemical changes in the metabolic activities of plant brought about by the lead contaminants. The knowledge of physiological and biochemical basis of phytotoxicity thus gained may help in understanding the possible constraints in the role of lead pollutants in the plants. In the present investigation an attempt is made here to elucidate the effect of lead on seed germination, seedling growth and nitrate reductase activity. The mungbean seeds occupy a very important position in Indian agriculture.
MATERIALS AND METHODS
The seeds of mung bean (Vigna radiata) used in present investigation were obtained from National seed corporation, New Delhi and study was carried out in 2006. They were surface sterilized with 0.1% bleaching powder (CaOCl2) for about 1-2 min. and then washed thoroughly with distilled water. The seedlings were raised on moist filter paper in petri plates and in small plastic pots containing washed sterilized sand for 15 d either in light approximately 60 wn-2 a mixture of coal white fluorescent tubes and incandescent bulbs or in the dark for chlorophyll biosynthesis investigation at 25 ±2°C. Seedling were watered daily with modified 2 strength Hoagland`s solution containing either KNO3 (10 nm) as sole nitrogen as per experimental design. Lead acetate was included either in the nutrient medium or in the incubation medium as the case may be. The pH in the each case was 6.0.
Mungbean seed soaked in the petridishes containing equal volumes of different concentrations of lead acetate solutions for 24 h arranged in germination towels and allowed to germination for 7 days. On the 7th day the percentage of seeds germination, length of the primary root, shoot and number of lateral roots formed per seedling were recorded. Changes in fresh and dry weights of the seedlings also were recorded.
For the NR activity secondary leaves for uniformly grown seedling were sampled and used for various studies. Lead was supplied to the excised secondary leaves in the incubation medium for 24 h at 25 ±2°C in the light.
In vivo nitrate reductase activity was assayed by the method of Srivastava (1975) and in vitro by the method of Stevens and Oaks (1973) with slight modifications. In intact seedlings nitrate reductase activity was measured only in the leaves as they are believed to be major nitrate reducing organic in most plants.
For the pigments content estimation fresh leaf samples were extracted with 80% acetone. Absorbance of clear supernatant were measured at 440, 645 and 663 nm. Total chlorophyll, carotenoids were calculated by method of Strain and Svec (1966) and Ikan (1969), respectively.
Total solution protein was estimated by the method of Lowry et al. (1951). After precipitating the extract in phosphate buffer (pH 7.0) with an equal volume of 20% TCA followed by solubilization with in NaOH solution. Total organic nitrogen was estimated by the Microkjeldhal method after digesting the dried leaf samples in conc. H2SO4 (Lang, 1958).
The data presented are average ±SD of three duplicate experiments. Employing the t-test the significant differences from the control were statistically evaluated at p = 0.05 and 0.01.
RESULTS
The results indicate that the growth rate of root and shoots were found to be retarded with increasing concentrations of lead acetate (Table 1). The fresh weight of root increased significantly at 0.01 mM lead supply. Further increase in the metal concentration i.e. 2.0 mM effect the root fresh wt. adversely shoot fresh weight also showed significantly a similar pattern of affect though with a lower magnitude. There was decreased fresh weight on shoot at 2.0 mM lead concentration. Dry weight of root increased non-significantly at 0.01 mM concentration decreased as in Table 1. Like root dry weight of shoot also increased slightly but significantly (p = 0.01) and the higher concentration of metal decreased dry weight of shoot significantly.
The effect of Pb of NRA was dependent on the concentration of the metal, mode of its supply and whether the assay was done by the vivo or in vitro method. When the Pb was supplied to the intact seedlings in the nutrient medium in vivo NRA in the leaves was stimulated slightly at 0.01 mM and inhibited (5-20%) at higher concentrations (Table 2). However, when lead was supplied to the seedlings in the nutrient solution. The in vivo enzyme activity in leaves (intact leaves) was inhibited consistently. The inhibition increased with the increase in Pb concentration. However, the in vitro activity of the enzyme either increased or was unaffected at low concentrations of lead while inhibited at higher concentrations (Table 2).
The nitrogenous pigment chlorophyll and non-nitrogenous carotenoid in the excised leaf segments to varying. Concentrations of Pb for 24 h had very little effect on total chlorophyll and carotenoid
| Table 1: | The effect of lead on seed germination and seedling growth in mungbean |
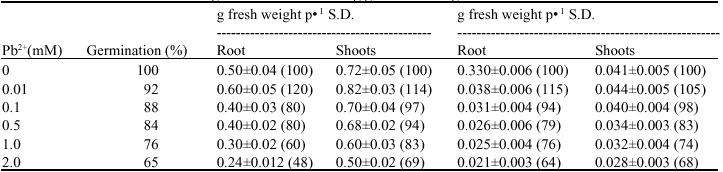 | |
| Vigna seeds were grown for 7 days as described in material and methods. Roots and shoots were dissected out from uniformly grown sampled plants and dried in oven for 2 days before dry weight measurements. ±: The standard error, +: The significance from the control at p = 0.05 **: The significant differences from control at p = 0.01 | |
| Table 2: | the effect of lead on in vivo in in vitro nitrate reductase activity in the Vigna radiata |
 | |
| The desired concentration of lead was included either in the incubation medium 1/10 strength Hoagland`s Solution (excised leaves) or in the nutrient solution $ strength Hoagland`s Solution (intact leaves). In either case the medium/solution contained 10 mM KNO3 as the sole nitrogen source. Values related to control as a percent are given in parentheses | |
| Table 3: | The effect Pb on the total chlorophyll and carotenoid in the Vigna radiata leaves |
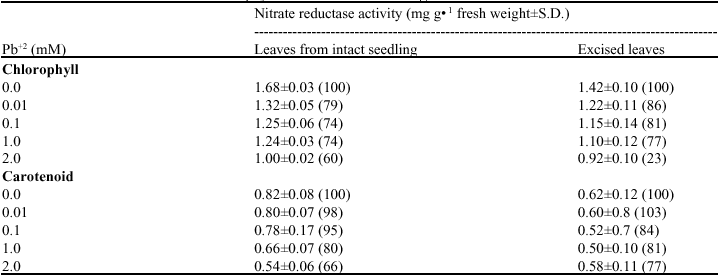 | |
| The desired concentration of lead was included either in the incubation medium 1/10 strength Hoagland`s Solution (excised leaves) or in the nutrient solution $ strength Hoagland`s Solution (intact leaves). In either case the medium/solution contained 10 mM KNO3 as the sole nitrogen source. Values related to control as a percent are given in parentheses | |
| Table 4: | The effect of Pb on the total soluble protein and total organic nitrogen in the Vigna radiata leaves |
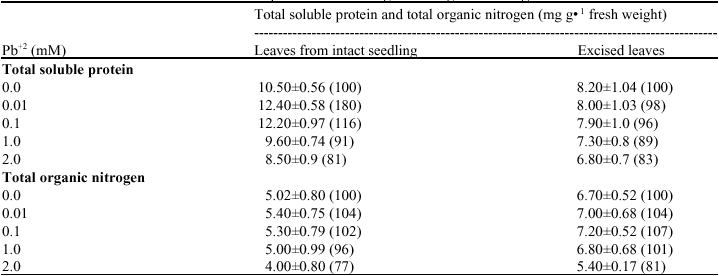 | |
| The desired concentration of lead was included either in the incubation medium 1/10 strength Hoagland`s Solution (excised leaves) or in the nutrient solution $ strength Hoagland`s Solution (intact leaves). In either case the medium/solution contained 10 mM KNO3 as the sole nitrogen source. Values related to control as a percent are given in parentheses | |
contents (Table 3). However, when the Pb was supplied to the seedlings, the chlorophyll content decreased, the reduction generally was correlated with the Pb concentration. Although the total carotenoid content was also slightly reduced by Pb, there was no correlation between Pb concentration and the fall in carotenoid content.
In excised leaf segments to varying concentrations of Pb had small negative effect on the amount of total soluble protein although there was no correlation between Pb concentration and the decrease (Table 4). On the other hand total organic nitrogen in leaf segments increased with Pb supply except for 0.01 and 1.0 mM, where there was no effect. Supply of Pb to intact seedlings had a small promontory effect on leaf protein. With the exception of 0.5 mM where there was a small decline. The total organic nitrogen was generally unaffected except for 1.0 mM where a slight drop was noticed.
DISCUSSION
In the present study relatively, higher concentration of lead inhibited both seed germination and seedling growth. Earlier heavy metals inhibited seed germination and seedling growth has been reported in maize (Seregin et al., 2004; Stiborova et al., 1987; Vojtechova and Leblova, 1991) and bajra (Prassad and Prassad, 1987). Early seedling growth is also inhibited in rice (Verma and Dubey, 2003; Yang et al., 2000), corn plants (Tung and Temple, 1996; Tu Shu and Brouillette, 1987). Spruce (Vodnik et al., 1999), Barley (Stiborova et al., 1987), tomato and egg plant (Khan and Khan, 1983). Root elongation, stem elongation, leaf elongation and leaf expansion were inhibited by Pb in Alium species and Barley (Juwarkar and Shende, 1986). Variable effect of lead on plant growth have been observed by various workers. Inhibition in fresh weight, dry weight and length of root and shoot of Sesamum indicum Cv. HT-I (Kumar et al., 1992), maize (Iqbal and Mushtaq, 1987) have been observed. These inhibitory effects of lead on growth and biomass accumulation are possibly a consequence of its effect on metabolic processes of plant (Thapa et al., 1988; Van Assche and Clijsters, 1990).
Inhibition in nitrate reductase activity by metal stress has been reported in Sunflower (Helianthuas annus), Sorghum (Pennisetum tyopholdes) leaves, as well as (Venketramana et al., 1978), maize (Sinha et al., 1994), mungbean (Rashid and Mukherji, 1991) and in higher plants (Smarelli et al., 1983).
In the present study, intact and excised leaves of seedlings treated with Pb showed a markedly different pattern in enzyme activity both in vivo as well as in vitro. The lower concentration of lead considerably increased NRA in vivo as well as in vitro in excised tissue of mungbean (Table 2). The possibility of direct influence of Pb on enzyme synthesis can not be ruled out.
The total protein level was not much altered by Pb (Table 3). It is possible that Pb affects a specific protein only chlorophyll content is not much affected in either case of treatment of intact or excised leaf tissues. But at the same time the insensitivity of mature tissue towards heavy metals has been also demonstrated (Koeppe, 1981).
Since protein and organic nitrogen content also remained unchanged, the experiments described that the enzyme nitrate reductase is much more sensitive to Pb pollution than other components on nitrate assimilation such as protein organic nitrogen and chlorophylls. Again the insensitivity of protein or organic nitrogen content to Pb inhibition in intact seedlings may be due to restricted presence of metal at the actual site of action in the leaf.
Although several reports indicates that plants can absorbs lead from the environment (Jarvis and Leung, 2002; Koeppe, 1981; Rudakova et al., 1988) and accumulate of lead was reported in roots (Atkins et al., 1982; Wozny et al., 1982). The accumulation of lead was reported in roots of Allium cepa also (Wierzbicka, 1987, 1994, 1998). Alternatively the intact plants may possess some kinds of detoxification mechanism which apparently lacks in excised leave tissue. The heavy metal pollution of the environment is on the increase at a very fast rate globally. In this context to assess the polluting effect of lead on different parts of the plants has become of absolute importance.
It can be concluded on the basis of the experimental findings of the present study that higher concentration of lead supplied to the plant results not only retareded germination and growth of the seedling of many been it deveases the chlorophyll level and the activity of nitrate reductase enzyme also.
REFERENCES
- Antonovies, J., A.D. Bradhava and R.G. Turner, 1971. Heavy metal tolerance in plants. Adv. Ecol. Res., 7: 1-85.
Direct Link - Blaylock, M.J., D.E. Salt, S. Dushenkov, O. Zakharova and C. Gussman et al., 1997. Enhanced accumulation of Pb in Indian mustard by soil-applied chelating agents. Environ. Sci. Technol., 31: 860-865.
CrossRefDirect Link - Broyer, T.C., C.M. Johnson and R.E. Paull, 1972. Some aspects of lead in plant nutrition. Plant Soil, 36: 301-313.
CrossRef - De Abreu, C.A., M.F. de Abreu and J.C. de Andrade, 1998. Distribution of lead in the soil profile evaluated by DTPA and Mehlich-3 solutions. Bragantia, 57: 185-192.
Direct Link - Eun, S.O., H.S. Youn and Y. Lee, 2000. Lead disturbs microtubule organization in the root meristem of Zea mays. Physiol. Plant, 110: 357-365.
CrossRefDirect Link - Iqbal, J. and S. Mushtaq, 1987. Effect of lead on germination, early seedling growth, soluble protein and acid phosphatase content in Zea mays. Pak. J. Sci. Ind. Res., 30: 853-856.
Direct Link - Jana, S. and M.A. Choudhuri, 1984. Synergistic effect of heavy metal pollutants on senescence in submerged aquatic plants. Water Air Soil Pollut., 21: 351-357.
CrossRef - Jarvis, M.D. and D.W.M. Leung, 2002. Chelated lead transport in Pinus radiata: An ultrastructural study. Environ. Exp. Bot., 48: 21-32.
Direct Link - Johnson, M.S. and J.W. Eaton, 1980. Environmental contamination through residual trace metal dispersal from a derelict lead-zinc mine. J. Environ. Qual., 9: 175-179.
Direct Link - Jones, J.H.P., S.C. Jarvis and S.C. Cowling, 1973. Lead uptake from soils by perennial regress and its relation to the supply of an essential element (sulpur). Plant Soil, 38: 605-619.
CrossRef - Khan, S. and N.N. Khan, 1983. Influence of lead and cadmium on the growth and nutrient concentration of tomato (Lycopersicum esculentum) and egg-plant (Solanum melongena). Plant Soil, 74: 387-394.
CrossRefDirect Link - Koeppe, D.E., 1977. The uptake, distribution and effect of cadmium and lead in plants. Sci. Total Environ., 7: 197-206.
Direct Link - Kumar, G., R.P. Singh and Sushila, 1993. Nitrate assimilation and biomass production in Sesamum indicuml. Seedlings in a lead enriched environment. Water Air Soil Pollut., 66: 163-171.
CrossRefDirect Link - Kumar, P.B.A.N., V. Dushenkov, H. Motto and I. Raskin, 1995. Phytoextraction: The use of plants to remove heavy metals from soils. Environ. Sci. Technol., 29: 1232-1238.
CrossRefDirect Link - Lane, S.D. and E.S. Martin, 1977. A histochemical investigation of lead uptake in Raphanus sativus. New Phytol., 79: 281-286.
Direct Link - Lang, C.A., 1958. Simple microdetermination of Kjeldhal nitrogen in biological materials. Ann. Chem., 30: 1692-1694.
Direct Link - Laxen, D.P.H. and R.M. Harrison, 1977. The highway as a source of water pollution: An appraisal of heavy metal lead. Water Res., 11: 1-11.
Direct Link - Lee, S.Z., L. Chang, H.H. Yang, C.M. Chen and M.C. Liu, 1998. Absorption characteristics of lead onto soils. J. Hazard. Mater., 63: 37-49.
Direct Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Marschner, P., D.L. Godbold and G. Jutschhe, 1996. Dynamics of lead accumulation in mycorrhizal and non-mycorrhizal Norway spruce (Picea abies L. karst.). Plant Soil, 178: 239-245.
Direct Link - Paivoke, A.E.A., 2002. Soil lead alters phytase activity and mineral nutrient balance of Pisum sativum. Environ. Exp. Bot., 48: 61-73.
CrossRef - Parekh, D. and R.M. Puranik, 1992. Inhibition of chlorophyll biosynthesis lead in greening maize leaf segments. Indian J. Exp. Biol., 30: 302-304.
Direct Link - Prassad, D.D.K. and A.R.K. Prassad, 1987. Altered δ-aminolevulinic acid metabolism by lead and mercury in germinating seedlings of Bajra (Pennisetum typhoideum). J. Plant Physiol., 127: 241-249.
CrossRefDirect Link - Rashid, P. and S. Mukherji, 1991. Changes in catalase and ascorbic oxidase activities in response to lead nitrate treatments in mungbean. Ind. J. Plant Physiol., 34: 143-146.
Direct Link - Sengar, R.S. and M. Pandey, 1996. Inhibition of chlorophyll biosynthesis by lead in greening Pisum sativum leaf segments. Biol. Plant, 38: 459-462.
CrossRef - Seregin, I.V., L.K. Shpigun and V.B. Ivaniov, 2004. Distribution and toxic effects of cadmium and lead on maize roots. Russian J. Plant Physiol., 51: 525-533.
Direct Link - Sinha, S.K., H.S. Srivastava and S.N. Mishra, 1988. Nitrate assimilation in intact and excised maize leaves in the presence of lead. Bull. Environ. Contamin. Toxicol., 41: 419-426.
CrossRef - Sinha, S.K., H.S. Srivastava and T.D. Tripathi, 1994. Influence of some growth regulators and divalent cations on the inhibition of nitrate reductase activity by lead in maize leaves. Chemosphere, 29: 1775-1782.
Direct Link - Smarelli, J. and H.C. Wilbur, 1983. Heavy metal inactivation and chelator stimulation of higher plant nitrate reductase. Biochem. Biophys. Acta, 742: 435-445.
Direct Link - Srivastava, H.S., 1975. Distribution of nitrate reductase inageing bean seedlings. Plant Cells Physiol., 16: 995-999.
Direct Link - Stefanov, K., K. Seizova, I. Popova, V.L. Petkov, G. Kimenov and S. Popov, 1995. Effects of lead ions on the phospholipid composition in leaves of Zea mays and Phaseolus vulgaris. J. Plant Physiol., 147: 243-246.
Direct Link - Stevens, D.L. and A. Oaks, 1973. The influence of nitrate in the induction of nitrate reductase in the maize roots. Can. J. Bot., 51: 1255-1258.
CrossRef - Stiborova, M., M. Ditrichova and A. Brezinova, 1987. Effect of heavy metal ions on growth and biochemical characteristics of photosynthesis of barley and maize seedlings. Biol. Planta, 29: 453-467.
CrossRefDirect Link - Tu Shu, I. and J.N. Brouillette, 1987. Metal ion inhibition of corn root plasma membrane ATPase. Phytochemistry, 26: 65-69.
Direct Link - Tung, G. and P.J. Temple, 1996. Uptake and localization of lead in corn (Zea mays L.) seedlings, a study by histochemical and electron microscopy. Sci. Total Environ., 188: 71-85.
CrossRefDirect Link - Van Assche, F. and H. Clijsters, 1990. Effect of metals on enzyme activity in plants. Plant Cell. Environ., 13: 195-206.
Direct Link - Verma, S. and R.S. Dubey, 2003. Lead toxicity induces lipid peroxidation and alters the activities of antioxidant enzymes in growing rice plants. Plant Sci., 164: 645-655.
Direct Link - Vodnik, D., G. Jentschke, E. Fritz, N. Gogala and D.L. Godbold, 1999. Root-applied cytokinin reduces lead uptake and affects its distribution in Norway spruce seedlings. Physiol. Plant, 106: 75-81.
Direct Link - Vojtechova, M. and S. Leblova, 1991. Uptake of lead and cadmium by maize seedlings and the effect of heavy metals on the activity of phosphoenolpyruvate carboxylase isolated from maize. Biol. Plant, 33: 386-394.
CrossRef - Wierzbicka, M., 1987. Lead accumulation and its translocation in roots of Allium cepa L.-autoradiographic and ultrastructural studies. Plant Cell Environ., 10: 17-26.
Direct Link - Wierzbicka, M., 1994. Resumption of mitotic activity in Allium cepa root tips during treatment with lead salts. Environ. Exp. Bot., 34: 173-180.
Direct Link - Wierzbicka, M., 1998. Lead in the apoplast of Allium cepa L. root tips-ultra structural studies. Plant Sci., 133: 105-119.
Direct Link - Yang, Y.Y., J.Y. Jung, W.Y. Song, H.S. Suh and Y. Lee, 2000. Identification of rice varieties with high tolerance or sensitivity to lead and characterization of the mechanism of tolerance. Plant Physiol., 124: 1019-1026.
PubMedDirect Link