
A. Longchar
Cell and Tumor Biology Laboratory, Department of Zoology, North-Eastern Hill University, Shillong-793022,
India
S.B. Prasad
Cell and Tumor Biology Laboratory, Department of Zoology, North-Eastern Hill University, Shillong-793022,
India
Research Journal of Mutagenesis
Year: 2016 | Volume: 6 | Issue: 1 | Page No.: 1-21







ABSTRACT
Cisplatin is a potent anticancer drug which has been in use against a variety of cancers for a long time. The present study was carried out to assess the protective ability of ascorbic acid on cisplatin-induced mutagenic effects in tumor-bearing mice. The increase in the frequency of chromosomal aberrations, micronuclei and sperm abnormality were studied as markers of mutagenicity. Indicators of oxidative stress relatives like lipid peroxidation, reduced glutathione and the activities of enzymes such as catalase, glutathione-S-transferase and glutathione reductase were also studied to understand the possible mechanism(s) underlying this amelioration. Pre-treatment with ascorbic acid in combination group significantly reduced cisplatin-induced mutagenicity, markedly decreased lipid peroxidation along with increased level of glutathione and activities of antioxidant enzymes particularly in the liver of mice. The overall studies suggest that ascorbic acid with its antioxidant and free radical scavenging properties may play a part in its protective role in the abatement of cisplatin-induced mutagenesis and oxidative damage in the normal tissues of mice.
PDF Abstract XML References Citation
Received: September 11, 2015;
Accepted: November 09, 2015;
Published: December 09, 2015
How to cite this article
A. Longchar and S.B. Prasad, 2016. Ascorbic Acid (Vitamin C)-Mediated Protection on Mutagenic Potentials of Cisplatin in Mice Bearing Ascites Dalton’s Lymphoma. Research Journal of Mutagenesis, 6: 1-21.
DOI: 10.3923/rjmutag.2016.1.21
URL: https://scialert.net/abstract/?doi=rjmutag.2016.1.21
DOI: 10.3923/rjmutag.2016.1.21
URL: https://scialert.net/abstract/?doi=rjmutag.2016.1.21
INTRODUCTION
Cisplatin (cis-diamminedichloroplatinum-II) is an inorganic platinum-based cancer chemotherapeutic drug that is extensively used against a variety of malignancies including testicular, ovarian, lung and breast cancer (Frezza et al., 2010). The anticancer activity of cisplatin mainly involves the formation of cisplatin-DNA adducts with inter and intra-strand nuclear DNA cross-links which account for about 90% of the total DNA damage induced by cisplatin to cause cytotoxic effects (Ramadan et al., 2001; Florea and Busselberg, 2011). Although cisplatin has been a mainstay for cancer therapy, its regular use is primarily limited by 2-3 factors i.e., development of acquired resistance by cancer cells to cisplatin and adverse side effects such as nephrotoxicity, ototoxicity, neurotoxicity, etc. and the possible induction of genotoxicity in non-tumor cells of the host (Gentile et al., 1998; Elsendoorn et al., 2001). The clastogenicity of cisplatin in humans and experimental animals has been well documented. Cisplatin induces formation of micronuclei in bone marrow cells and chromosomal aberrations in the germinal cells of mice (Kliesch and Adler, 1987; Adler and El-Tarras, 1989) and also the micronuclei in peripheral blood lymphocytes of testicular patients with various types of cancers (Osanto et al., 1991; Elsendoorn et al., 2001). It is imperative to reduce the drug-induced genotoxicity, a goal that have been tried experimentally by administration of free radical scavengers such as antioxidants (Antunes et al., 1999, 2000a; Attia, 2010; Dos Santos et al., 2012) with varying degree of success.
Ascorbic Acid (AA) or vitamin C has been reported to be effective as a protectant against a variety of toxic chemical agents including heavy metals (Fox, 1975; Holloway and Peterson, 1984). The protective role of ascorbic acid on cisplatin-induced nephrotoxicity (Antunes et al., 2000b; Ajith et al., 2007) and mutagenicity (Giri et al., 1998; Antunes et al., 2000a; Nefic, 2001) have also been reported in both humans and animal models. The use of anticancer drugs such as cyclophosphamide (Nicol and Prasad, 2006), paclitaxel (Park et al., 2012), arsenic trioxide (Yedjou et al., 2009), mitomycin C (Krishnaja and Sharma, 2003), doxorubicin (Viswanatha Swamy et al., 2011) and chlorambucil (Kalita et al., 2014) in combination with vitamin C have also been examined.
Ascorbic acid or vitamin C is involved in various biological processes including free radical scavenging function and plays an important role in the metabolism and detoxification of many endogenous and exogenous compounds (Henson et al., 1991). Mechanisms of antioxidative action of ascorbic acid operate through direct scavenging and blocking of reactive oxygen species (ROS) (Griffiths and Lunec, 2001). The role of ascorbic acid in general medicine and in anticancer therapy has been discussed (Verrax and Calderon, 2008). It has been demonstrated that treatment with ascorbic acid (vitamin C) significantly reduced the genotoxicity of well-known mutagens (Cabrera, 2000; Blasiak and Kowalik, 2001; Siddique et al., 2005; Misra and Choudhury, 2006). However, large amounts of ascorbic acid has been reported to cause some genotoxic effects in in vitro test systems (Shamberger, 1984; Nefic, 2008) because ascorbic acid has both antioxidant and pro-oxidant activities (Bijur et al., 1997).
Cisplatin-based combination chemotherapy regimens may be used as front-line therapy in the treatment of various cancers. Most of the studies involving implication of ascorbic acid in the modulation of antitumor effects or toxicities induced by various anticancer drugs have been done separately in cancer cell lines/tumors in vivo or in normal animals, respectively. Since cancers may alter the metabolic and endocrine equilibrium in the host (Barbosa et al., 2009), it is reasonable that the ascorbic acid-mediated protective strategies on cisplatin-induced mutagenic effects should be tested in tumor-bearing hosts which may also propose that inspective protective agents are useful in enhancing therapeutic efficacy. Our recent studies have shown that the combination treatment using ascorbic acid with cisplatin show better antitumor activity and reduced tissue toxicities in the hosts (Amenla et al., 2013; Longchar and Prasad, 2015). Thus, the present studies were undertaken to further examine the modulatory effects of combination treatment of cisplatin with ascorbic acid on cisplatin-induced mutagenic potentials in tumor-bearing mice.
MATERIALS AND METHODS
Chemicals: Cisplatin solution (1 mg mL–1 of 0.9%, NaCl) was obtained from Biochem Pharmaceutical Industries, Mumbai, India. L-ascorbic acid (vitamin C) was purchased from HiMedia laboratories, Mumbai, India. Reduced glutathione (GSH), 5,5ʹ-dithiobis-2-nitrobenzoic acid (DTNB), 1-chloro-2,4-dinitrobenzene (CDNB), nicotinamide adenine dinucleotide phosphate reduced (NADPH) and bovine serum albumin (BSA) were purchased from Sigma Chemical Co., St. Louis, Mo, USA. Ethylenediamine tetra-acetic acid (EDTA), 2-thiobarbituric acid (TBA) and other chemicals used in the experiments were of analytical grade and purchased from SRL Pvt. Ltd., Mumbai, India.
Tumor maintenance: Swiss albino mice inbred colony is being maintained in the laboratory under conventional condition at 24±2°C with free access to standard food pellet diets (Amrut Laboratory, New Delhi) and drinking water ad libitum. Ascites Dalton’s Lymphoma (DL) tumor is being maintained in laboratory in in vivo condition in 10-12 weeks old mice by serial intraperitoneal (i.p.) transplantation of approximately 1×107 viable tumor cells per animal (in 0.25 mL phosphate-buffered saline (PBS), pH 7.4). Early sign of tumor development was easily visible by the increase in belly size after 3-4 days of tumor transplantation. Tumor transplanted hosts usually survive for 19-21 days.
The maintenance and use of the mice and the experimental protocol of the present study has been approved by the Institutional ethical committee of North-Eastern Hill University, Shillong, India.
Drug treatment schedule: The treatment of tumor-bearing mice with cisplatin (10 mg kg–1 b.wt.) and ascorbic acid (1% in drinking water) has been used previously in our study (Amenla et al., 2013) and the same treatment schedule was followed in the present study also. Three groups (i.e., control, cisplatin and AA plus cisplatin combination) of tumor-transplanted mice were selected for the present study as AA alone treatment group did not show any significant changes as compared to control group in our previous investigation. Tumor-transplanted mice were randomly divided into three groups consisting of 10 mice in each group. Group-I mice served as tumor-bearing control and received normal saline only. Group-II mice were administered with a single dose of cisplatin (10 mg kg–1 b.wt.) on the 10th day post-tumor transplantation. Group-III mice received AA through drinking water from the 5th day post-tumor transplantation and were administered with cisplatin (i.p., 10 mg kg–1 b.wt.) on the10th day of tumor growth. After 24, 48, 72 and 96 h of cisplatin treatment (i.e., on 11, 12, 13 and 14th day of tumor transplantation) two animals were sacrificed by cervical dislocation and the bone marrow and other tissues were used for the mutagenic and biochemical investigation as detailed below and the experiments were repeated thrice.
Chromosomal aberration analysis: Chromosomal aberration analysis was carried out following the method of Sharma and Sharma (1994). Mice in different groups were subjected to mitotic arrest initiated 2 h prior to sacrificing by i.p injection of colchicine (4 mg kg–1 b.wt.). Bone marrow was collected from femur by flushing in PBS (pH 7.4) using a syringe. Bone marrow cells were collected by centrifugation (1000 rpm, 4°C for 5 min). The 5 mL of pre-warmed hypotonic solution was added to the cell pellet and incubated for 25 min at 37°C followed by gentle mixing of few drops of Carnoy’s fixative (methanol: glacial acetic acid, 3:1, v/v) and centrifuged at 1000 rpm for 5 min. The supernatant was discarded and the pellets were dissociated slowly by gentle tapping. To this, 5 mL of chilled Carnoy’s fixative was added and incubated at 4°C for 30 min. Fixation was repeated twice with an interval of 30 min. Finally, the cells were re-suspended in a small volume (0.5 mL) of fixative. A few drops of this suspension was dropped on cleaned, grease-free chilled slides, burnt on a flame, air-dried and stained the following day with freshly prepared working Giemsa stain in Sorensen’s buffer (pH 6.8) and mounted in DPX. Three hundred good metaphase spreads were examined per animal and chromosome aberrations were classified into the general categories of chromatid breaks, exchanges, isochromatid breaks, chromosomal fragments and sister chromatid unions.
Micronucleus assay: Micronuclei were studied in different groups of mice following the method of Schmid (1976). Both femura of mice were removed and bone marrow was flushed out in PBS (pH 7.4) in a centrifuge tube. Bone marrow cells were collected by centrifugation (1000 rpm, 4°C for 5 min). The cell pellet was treated with a weak hypotonic solution (0.075 M KCl/saline, 1:9, v/v). After centrifugation, cells were fixed in Carnoy’s fixative and repeated twice. Cells were re-suspended in a small volume of fixative solution. A drop of this suspension was smeared onto wet chilled slide, air-dried and stained with Giemsa in Sorensen’s buffer (pH 6.8). One thousand cells were examined from each treatment group under the microscope (Leica) for the presence of micronuclei.
Sperm abnormality assay: Male mice in different groups were sacrificed after 10 days of cisplatin treatment. The cauda epididymis were removed and placed in physiological saline. It was then minced into pieces and kept undisturbed for 20 min for diffusion of spermatozoa. The spermatozoa were spread on a clean slide, air-dried, fixed in absolute methanol for 15 min and then stained with 1% aqueous eosin-Y on the following day. Five hundred sperms from each mouse were examined for the abnormalities in sperm head and tail shapes following the criteria as close as possible to those established by Wyrobek and Bruce (1975).
Lipid peroxidation assay: The extent of lipid peroxidation (LPO) was measured in terms of malondialdehyde (MDA) concentration in different tissues of mice using the method of Buege and Aust (1978) as described in detail earlier (Longchar and Prasad, 2015).
Reduced glutathione and GSH-related enzymes assay: Total glutathione (GSH) was determined in the tissues (liver, kidney and testes) collected from DL-bearing mice under different experimental conditions using the method of Sedlak and Lindsay (1968), as explained and used earlier (Longchar and Prasad, 2015).
The same tissues were washed in ice-cold physiological saline and used for GSH enzymatic activity assay. A 10% (w/v) tissue homogenate was prepared in a motor-driven Teflon-pestle homogenizer at 0±2°C after the addition of 0.1 M potassium phosphate-1 mM Na2 EDTA buffer (pH 6.5) for glutathione-S-transferase assay, 0.2 M potassium phosphate-2 mM Na2 EDTA buffer (pH 7.5) for glutathione reductase assay and 1% Triton X-100 for catalase assay. The tissue homogenates were centrifuged at 27000×g for 20 min at 4°C in a cooling centrifuge and the respective supernatants were used as the enzyme source.
Glutathione-S-transferase (GST; EC 2.5.1.18): Glutathione-S-transferase activity was assayed following the method of Habig et al. (1974). The assay volume (1.0 mL) contained 850 μL of 0.1 mol L–1 potassium phosphate-1 mmol L–1 Na2 EDTA buffer (pH 6.5), 50 μL of 30 mmol L–1 CDNB in 95% ethanol and 50 μL of 30 mmol L–1 reduced glutathione in deionised water. The overall reaction (maintained at 30°C) was started by adding 50 μL tissue supernatant as the enzyme source and the increase in absorbance was monitored at 340 nm. One unit of enzyme activity is defined as the amount of enzyme that catalyses the conjugation of 1 μmol of CDNB per minute.
Glutathione reductase (GR; EC 1.6.4.2): Glutathione reductase activity was assayed according to Smith et al. (1988). Total assay volume (1.0 mL) contained 500 μL of 0.2 mol L–1 potassium phosphate-2 mmol L–1 Na2 EDTA buffer (pH 7.5), 250 μL of 3 mmol L–1 DTNB in 0.01 mol L–1 phosphate buffer, 150 μL water, 50 μL of 2 mmol L–1 NADPH in 10 mmol L–1 Tris-HCl buffer (pH 7.0) and 50 μL of enzyme source. The reaction was initiated by the addition of 50 μL of 20 mM GSSG to the reaction mixture and the decrease in absorbance at 37°C was monitored at 340 nm. One unit of enzyme activity is defined as the amount of enzyme that catalyses the reduction of 1 μmole of NADPH per minute.
Catalase (CAT; EC 1.11.1.6): Catalase activity was determined following the method of Aebi (1984). The assay volume (3.0 mL) contained 20 μL of tissue supernatant as the enzyme source and 1.98 mL of 50 mol L–1 phosphate buffer (pH 7.0) with 1.0 mL of 30 mol L–1 H2O2. The CAT activity was measured by monitoring a decrease in absorbance of H2O2 at 240 nm. One unit of enzyme activity is expressed as 1 μM of H2O2 decomposed per minute.
The protein concentration was determined in the same supernatant used for various enzyme assays following the method of Lowry et al. (1951). The particular enzyme specific activity was calculated by dividing the number of units of that enzyme by milligrams of protein present in that assay of tissue supernatant.
Statistical analysis: The values have been expressed as Mean±Standard Deviation (SD) and all determinations were repeated thrice. One-way analysis of variance (ANOVA) was used to evaluate the difference among multiple groups followed by a post hoc test (Tukey test). A p-value ≤0.05 was considered as statistically significant.
RESULTS
Chromosomal aberration analysis: Various types of chromosomal aberrations such as chromatid breaks, isochromatid breaks, exchange, chromosomal fragments and sister chromatid union were observed in bone marrow cells under different treatment conditions (Fig. 1). Maximum frequency of various aberrations was seen at 24 h of treatment. Mean aberrant metaphases decreased appreciably with the duration of treatment from 24-96 h in cisplatin alone (Fig. 2a) as well as in combination treated groups (Fig. 2b).
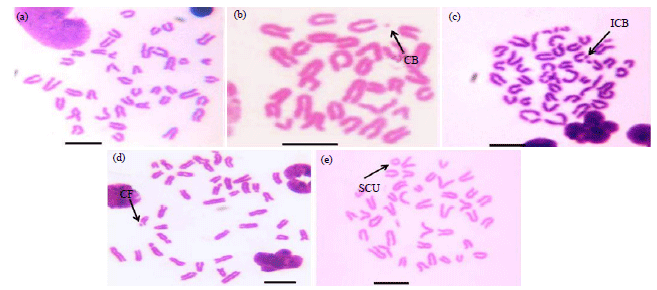 | |
| Fig. 1(a-e): | Representatives of bone marrow metaphase chromosome spreads of mice showing (a) Normal set of chromosomes and different types of chromosomal aberrations under different treatment conditions; different types of aberrations induced by cisplatin are (b) Chromatid break (CB), (c) Isochromatid break (ICB), (d) Chromosomal fragment (CF) and (e) Exchange (Exch) and Sister Chromatid Union (SCU). Scale bar = 5 μm |
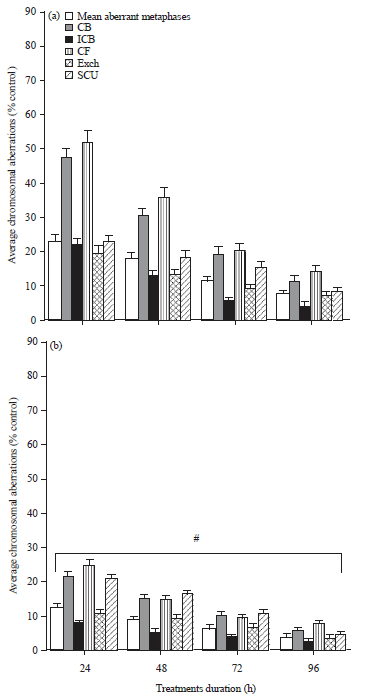 | |
| Fig. 2(a-b): | Quantitative analysis of mean aberrant metaphases i.e., CB: Chromatid break, ICB: Isochromatid break, CF: Chromosomal fragment, Exch: Exchanges, SCU: Sister chromatid union after treatment with (a) Cisplatin and (b) AA plus cisplatin. Results are expressed as Mean±SD, n=3, #p≤0.05 as compared to cisplatin. AA: Aascorbic acid |
Cisplatin alone treatment revealed maximal increase in the frequency of chromosomal aberrations than the other treated groups with chromatid breaks and chromosomal fragments occurring more frequently in the bone marrow cells of mice (Fig. 2). Comparative analysis at corresponding periods of treatment revealed that the chromosomal aberrations were significantly lesser in the group of mice treated with combination of AA plus cisplatin than those treated with cisplatin alone (Fig. 2).
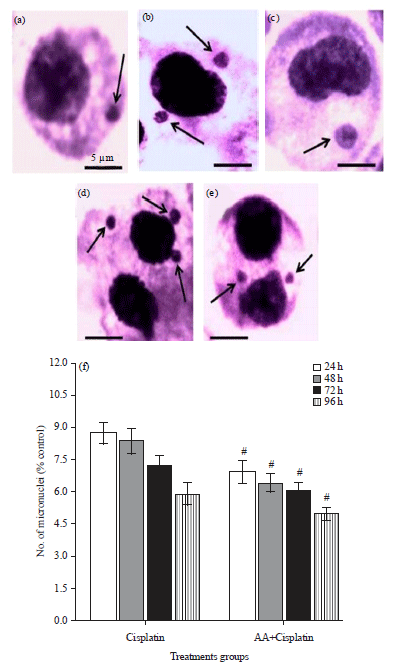 | |
| Fig. 3(a-f): | (a-e) Photomicrographs of bone marrow cells of mice showing micronuclei (arrows) induced by cisplatin and (f) Quantitative percentage analysis of micronuclei at different treatment conditions. Results are expressed as Mean±SD, n = 3, #p≤0.05 as compared to cisplatin |
Micronucleus study: The incidence of micronuclei in bone marrow cells of mice significantly increased after cisplatin treatment. One or more micronuclei were observed after cisplatin alone (Fig. 3a-e) and combination treatment (24-96 h) with maximum frequency of micronuclei at 24 h of treatment (Fig. 3f). However, AA pre-treatment in the combination group significantly decreased the frequency of micronuclei at corresponding time points as compared to cisplatin alone treatment (Fig. 3f).
Sperm abnormality study: Cisplatin induced various types/shapes of sperm abnormalities which include hookless head, looping mid-piece, microhead, balloon-like head, double tailed, incorrect head-neck connection, diffused head, banana head, amorphous head, etc (Fig. 4b-j).
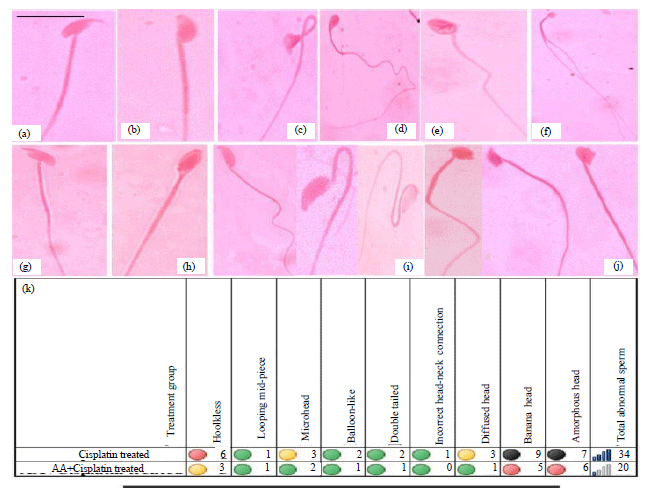 | |
| Fig. 4(a-k): | Representatives of spermatozoa from mice showing various types of abnormalities at different treatment conditions, (a) Normal, (b) Hookless, (c) Looping mid-piece, (d) Microhead, (e) Balloon-like, (f) Double tailed, (g) Incorrect head-neck connection, (h) Diffused head, (i) Banana head,(j) Amorphous head, (k) Quantitative analysis of various types of sperm abnormality after different treatment conditions. Data are shown over percent control. Sperm analysis was carried out on 10th day of cisplatin treatment. Scale bar = 10 μm, Green 0-2, Yellow 3-4, Pink 5-5 and Black 7-9% |
Comparative analysis of sperm abnormalities in mice treated with cisplatin alone showed maximum increase (~34%) in the frequency of sperm abnormalities. However, AA pre-treatment in combination treatment significantly decreased (~20%) the frequency of cisplatin-induced sperm abnormalities in the hosts (Fig. 4k).
Lipid peroxidation: As compared to corresponding control, cisplatin treatment caused an increase in the level of LPO in liver and kidney at different time points. However, in testes a decrease in LPO level was observed after cisplatin treatment (Fig. 5). As compared to cisplatin alone, combination treatment of mice caused significant decrease in the LPO level in liver (24-96 h), kidney (48-96 h) and testes (48-96 h) (Fig. 5).
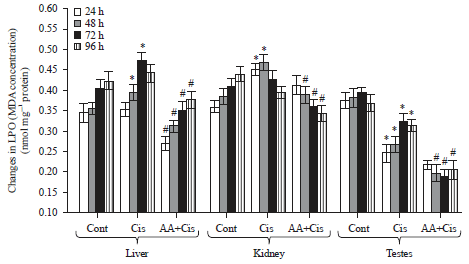 | |
| Fig. 5: | Changes in the level of malondialdehyde reflecting lipid peroxidation (LPO) in the tissues of tumor-bearing mice at different treatment conditions, Cont: Control, Cis: Cisplatin, AA+Cis: Ascorbic acid plus cisplatin. Results are expressed as Mean±SD, Tukey test: n = 3, *p≤0.05 as compared to the corresponding control, #p≤0.05 as compared to cisplatin |
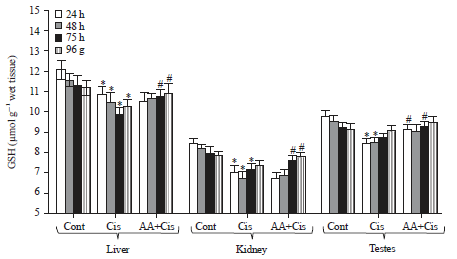 | |
| Fig. 6: | Changes in total reduced glutathione (GSH) level in the liver, kidney and testes at different treatment conditions, Cont: Control, Cis: Cisplatin, AA+Cis Ascorbic acid plus cisplatin. Results are expressed as Mean±SD, ANOVA: n = 3, *p≤0.05 compared to the corresponding control, #p≤0.05 as compared to cisplatin |
Reduced glutathione and GSH-related enzymatic study: The changes in total GSH level in liver, kidney and testes under different treatment conditions are shown in Fig. 6. As compared to corresponding control, cisplatin treatment of tumor-bearing mice resulted in a significant decrease in GSH level in liver (24-96 h), kidney (24-72 h) and testes (24-48 h) of treatment. As compared to cisplatin alone, AA plus cisplatin treatment caused a significant increase in GSH levels in liver and kidney at 72-96 h and in testes during 24-72 h of treatment (Fig. 6).
Glutathione S-transferase (GST; EC 2.5.1.18) activity: As compared to corresponding control, cisplatin treatment caused a decrease in GST activity in liver (24-96 h), while significant increase in GST activity was noted in kidney and testes during 24-48 h of treatment, respectively. As compared to cisplatin alone, combination of AA plus cisplatin treatment caused an increase in GST activity in liver (24-96 h) while differential pattern of decrease was seen in kidney and testes (Fig. 7a).
Glutathione reductase (GR; EC 1.6.4.2) activity: As compared to corresponding control, cisplatin treatment resulted in a decrease in GR activity in liver (24-48 h) and testes (24-96 h). However, in kidney an increase in GR activity was noted during 24-96 h.
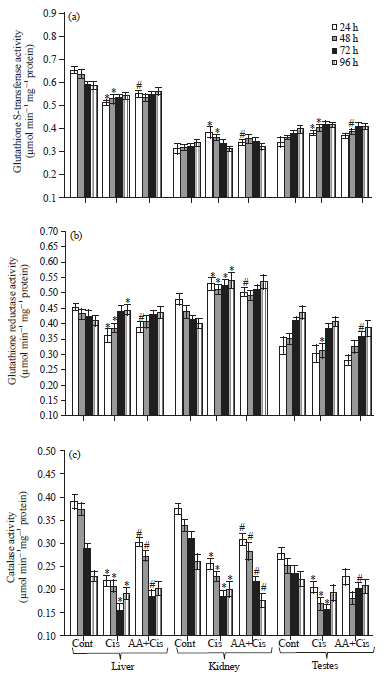 | |
| Fig. 7(a-c): | Changes in (a) Glutathione S-transferase, (b) Glutathione reductase and (c) Catalase activity in the tissues (liver, kidney and testes) at different treatment conditions. Results are expressed as Mean±SD, ANOVA, *p≤0.05 as compared to the corresponding control, #p≤0.05 as compared to cisplatin |
As compared to cisplatin alone, combination treatment caused an increase in GR activity in liver and decrease in kidney and testes at different time points (Fig. 7b).
Catalase (CAT; EC 1.11.1.6) activity: As compared to corresponding control, cisplatin treatment of tumor-bearing mice caused a time-dependent decrease in catalase activity in liver, kidney and testes during 24-72 h of treatment. As compared to cisplatin alone, combination treatment of mice caused an increase in catalase activity in liver and testes during 24-96 h and in kidney at 24-72 h of treatment (Fig. 7c).
DISCUSSION
Developments of chromosomal aberrations, micronuclei and sperm abnormalities in response to chemical exposure are effective cytogenetic biomarkers useful to identify mutagenic agents (Giri et al., 2002; Garcia-Sagredo, 2008). Bone marrow cells are susceptible to oxidative damage and sensitive to clastogenic chemicals so, it has been commonly used for screening of mutagenicity and/or antimutagenicity property of chemicals (Umegaki et al., 1997). The results of present investigation showed development of these mutagenic parameters after cisplatin alone or combination treatment of tumor-bearing mice and supports the earlier findings of its genotoxic properties (Adler and El-Tarras, 1989; Brozovic et al., 2011). The chromosomal aberration pattern revealed that chromatid breaks and fragments occurred more frequently after cisplatin treatment (Fig. 2a). The clastogenicity of cisplatin in bone marrow cells was well investigated by Edelweiss et al. (1995) and Choudhury et al. (2000), who observed that the most impressive effect of a single dose of cisplatin was an increase in the frequency of chromosomal aberrations and in the number of abnormal metaphases after the treatment. The total frequencies of chromosomal aberrations as well as aberrant metaphases were observed to be maximal at 24 h of treatment, which gradually decreased during the later phase of cell cycle (Fig. 2) which also support the view that antitumor agents produce a high frequency of aberrations in rodents 24 h after single exposure, which roughly coincides with the normal length (22-24 h) of the cell cycle (Rosselli et al., 1990). These elevated levels may also be seen as an indicator of an early phase of carcinogenesis, where various genetic alterations are also generated in different tissues. However, decrease in the frequency of aberrations in the later hours of treatment could be due to death of damaged cells, clearance of mutagen from the body and post-replication repair process which might be operating for recovery from the cisplatin-induced damage to DNA. In fact, an involvement of post-replication repair process in cisplatin-induced DNA damage has been established (Sorenson and Eastman, 1988). Rosenberg (1985) proposed that cisplatin lesions on O6 of guanine in normal cells are repaired before replication, while in cancer cells the lesions are not removed because of a deficiency in this repair process, hence mutation rate increases beyond the limits of survivability. This might explain the variation in the frequency of chromosomal aberrations observed in the present study following 24-96 h cisplatin treatments (Fig. 2a). Ascorbic acid (vitamin C) has antioxidant and free radical scavenging activities, suggesting that this vitamin may modulate oxidative DNA damage in mammalian cells (Duthie et al., 1996; Odin, 1997). In the present study, total chromosomal aberrations were decreased appreciably in AA plus cisplatin treated group at corresponding time points (Fig. 2b) showing that ascorbic acid exhibited reduction in the chromosomal aberrations induced by cisplatin which may indicate its anticlastogenic effects against cisplatin-induced mutagenic effects as has also been suggested by others (Giri et al., 1998; Antunes et al., 1999, 2000a; Nefic, 2001). Based on these findings, it may be suggested that decrease in the frequency of chromosomal aberrations after combination treatment is due to the protective effect of ascorbic acid in the abatement of cisplatin-induced mutagens.
Micronuclei are indicators of chromosome instability, since the frequency of micronuclei is higher in tumor cells which show a defective DNA damage repair system and disrupted cell cycle checkpoint machinery (Terradas et al., 2010). An increase in the frequency of micronuclei in bone marrow cells in treated animals could be an indication of induced chromosomal damage. The present finding shows that exposure to cisplatin significantly elevated the frequency of micronuclei in bone marrow cells while pre-treatment with ascorbic acid decreased the frequency on cisplatin-induced micronucleated cells in mice bone marrow (Fig. 3). Moreover, the time-dependent decrease (24-96 h) in frequency of micronuclei observed during combination treatment (Fig. 3f) reveals that ascorbic acid could exert time-dependent antimutagenic effects. In fact, the ability of ascorbic acid to confer marked protection against many toxic chemical agents has been described. El-Refaiy and Eissa (2012) reported the protective effect of ascorbic acid against cadmium-induced cytotoxicity in rat bone marrow cells. Another study in rat bone marrow cells revealed protection by vitamin C or E against chromosomal damage induced by the antitumoral drug doxorubicin (Antunes and Takahashi, 1998).
Anticancer drugs/chemicals induce sperm abnormalities indicating its genotoxicity to germ cells (Wyrobek et al., 1983; Seetharama Rao and Narayana, 2005; Atessahin et al., 2006). Cisplatin has been considered as a potent mutagen causing formation of abnormal male germ cells (Sawhney et al., 2005; Mohammadnejad et al., 2012). In the present study, treatment of mice for 10 days with cisplatin induced various types of sperm abnormalities (Fig. 4b-j). During meiotic phase of spermatogenesis, the spermatocytes are in the G2 phase of the cell cycle (Eddy, 2002) and cisplatin is known to arrest the cell cycle at G2 and M2 phases (Sorenson and Eastman, 1988), therefore, this mechanism may be involved in developing cisplatin-mediated abnormalities in sperms. The abnormalities in sperm morphology have been suggested to be a consequence of chromosomal aberrations (Bruce et al., 1974; Al-Hakkak et al., 1986; Ushijima et al., 2000). Cisplatin treatment developed abnormalities in the sperms (Fig. 4k) suggesting its ability to reach the germ line cells and indicates its potentiality as a germ cell mutagen as well. The development in sperm abnormalities may indicate that cisplatin induced the DNA damage in germ cells leading to altered sperm morphology (Poirier et al., 1992). Several studies show that various species of ROS generated through metal catalysis potentially interact with gene strands causing mutations, thereby inducing changes in sperm morphology (Roy and Rossman, 1992; Hsu et al., 1998; Bench et al., 1999). Therefore, the present mechanism of abnormal production of sperm as observed in the present study may also be an oxidative stress-dependent phenomenon induced by platinum catalysis. However, combination with AA showed a significant decrease (Fig. 4k) in the abnormality which also depicts the similar protective role of ascorbic acid as observed for chromosomal aberration (Fig. 2b). Ayinde et al. (2012) reported that daily consumption of vitamin C could be useful in lowering oxidative stress and spermatozoa deformations in lead exposed albino rats. It has also been reported that co-administration of speman with cisplatin showed significant improvement on the sperm quantity and quality along with enhanced steroidogenesis in mice (Sainath et al., 2011). In fact the use of another antioxidant, i.e., melatonin in combination with cisplatin has also shown chemoprotective effect against cisplatin-induced testicular toxicity in rats (Atessahin et al., 2006) which also supports the present findings.
The combination treatment of mice with ascorbic acid plus cisplatin has much better therapeutic efficacy against murine ascites DL in comparison to cisplatin alone (Amenla et al., 2013). Thus, the results of these mutagenic parameters in the present study showing the significant reduction in cisplatin-induced mutagenic and genotoxic effects in presence of ascorbic acid strongly suggests that while increasing therapeutic efficacy, ascorbic acid effectively ameliorates the mutagenic potentials of cisplatin in the DL-bearing hosts.
Lipid peroxidation (LPO) is ascribed to a free radical-mediated chain reaction that damages cell membranes under oxidative stress conditions. Cisplatin is associated with induction of oxidative stress by generation of free radicals and reactive oxygen species (ROS) which are highly toxic to cells (Masuda et al., 1994; Mitazaki et al., 2013). When generation of the ROS overtakes the antioxidant defense of the cells, ROS can react with the polyunsaturated fatty acids of lipid membranes and induce lipid peroxidation (Droge, 2002). The end products of lipid peroxidation, 4-hydroxynonenal (HNE) have been considered to be a second messenger of oxidative stress. Beyond ROS involvement in carcinogenesis, increased ROS level can inhibit tumor cell growth with further increase of oxidative stress, which usually occurs when using several anticancer drugs and thereby overcome the antioxidant defenses of cancer cells finally leading to apoptosis. High concentrations of HNE can also induce apoptosis in cancer cells. Malondialdehyde (MDA), the product of LPO, also interacts with DNA causing strand breaks that in turn develop into chromosomal breaks. These chromosome breaks may appear as micronuclei in the daughter cell after the first cell division (Attia, 2010). In agreement with these report, the present study showed that cisplatin treatment caused significant increase in the LPO levels in tissues (Fig. 5), indicative of increased oxidative stress which may be attributed to tissue toxicities and may be subsequently involved in the formation of chromosomal aberrations in bone marrow cells (Fig. 2). Somani et al. (2000) also reported an increase in MDA content following cisplatin administration leading to impairment in kidney function. All cells maintain antioxidant defenses because of the damaging effects of ROS. High antioxidant intake has been shown to reduce cancer risk and may also mitigate the effects of oxidative DNA damage, which is hypothesized to be casually linked to carcinogenesis. Ascorbic acid is capable of inhibiting the activity of free radical-induced reactions (McCall and Frei, 1999). Pretreatment of ascorbic acid reduced the cisplatin-induced LPO significantly in the tissues (Fig. 5) thus demonstrating protection from cisplatin-induced tissue toxicities in the host. The decrease in LPO also implies the free radical scavenging property of ascorbic acid.
The role of glutathione (GSH) and glutathione-related enzymes in cancer and acquired resistance has been the focus of interest (Arrick and Nathan, 1984; Kidd, 1997). One of the most important intracellular antioxidant systems is the glutathione redox cycle which plays an important role in the detoxification of ROS generated by many endogenous toxins including cisplatin (Hrubisko et al., 1993; Chen et al., 1995), so as to protect cells from the potential toxicity and carcinogenesis. Elevation of GSH levels has been shown to increase the resistance of cancer cells to cisplatin (Meijer et al., 1990; Kartalou and Essigmann, 2001), while depletion of GSH levels could potentiate the cytotoxicity of a variety of antitumor agents (Arrick and Nathan, 1984). Several investigators have demonstrated that cisplatin induces ROS primarily by decreasing the activity of antioxidant enzymes and by depleting intracellular concentrations of GSH (Husain et al., 1998; Ognjanovic et al., 2012). Depletion in the renal GSH level in rats has also been reported in response to oxidative stress caused by cisplatin treatment (Silva et al., 2001). Therefore, the decrease in total GSH levels observed in liver, kidney and testes (Fig. 6) after cisplatin treatment may have a role in altering cellular antioxidant machinery and in lessening the protective ability which may facilitate cisplatin-mediated toxic effect in these tissues. The decrease level of GSH observed in testes of tumor bearing mice (Fig. 6) should also be helping to enhance cisplatin’s mutagenic effects in testes (Fig. 4). The depletion of GSH is also believed to be related to perturbation of intracellular calcium homeostasis by the increased cytosolic calcium which can lead to oxidative stress and cell injury (Pereira and de Oliveira, 2000). A decrease in cellular GSH concentration has been inversely correlated with lipid peroxidation in the liver (Trbojevic et al., 2010). It has been reported that the alterations induced by cisplatin in the tissues (particularly liver and kidney) were characterized by signs of injury such as increase of LPO products and changes in GSH levels (Naziroglu et al., 2004). It may, therefore, be suggested that the observed decrease in GSH level associated with an increase in LPO after cisplatin treatment in the present study could be an early and critical factor in cisplatin-induced cytotoxicity against cancer. Ascorbic acid and GSH are effective water-soluble cytoplasmic antioxidants participating in cellular protection against oxidative stress and toxic agents (Nagyova and Ginter, 1994). The cooperation between ascorbic acid and GSH in various protective effects has been reviewed by Meister (1994), where it was reported that giving ascorbate to animals led to increase GSH levels. In the present study, although initially combination treatment caused a decrease in the GSH level in tissues, it was restored to approximately control levels during 72-96 h of treatment. These elevations of total GSH in the tissues (Fig. 6) after combination treatment of AA plus cisplatin could also represent a protective mechanism in response to various toxic radicals thereby, suggesting the possible involvement of cellular GSH as a mechanistic step in AA-mediated protection against cisplatin-induced mutagenicity. It has been suggested that GSH and ascorbate pools in cells may interact with each other, producing cooperative antioxidative protective effects (Jain et al., 1992). The protective role of ascorbic acid against cisplatin-induced nephrotoxic effects has been noted with possible cooperative involvement of GSH in its protective function (Prasad et al., 2006). A protective role of ascorbic acid in 2,4-dichlorophenol induced teratogenic/carcinogenic toxicity along with significantly increased liver ascorbic acid and GSH levels has also been reported (Nagyova and Ginter, 1995).
Glutathione S-transferases (GSTs) are a family of phase II detoxification enzymes which catalyse the conjugation of GSH to a wide variety of xenobiotics, drugs and carcinogens (Townsend and Tew, 2003). This detoxification ability plays a role in cellular defenses from environmental and oxidative stress. Moreover, overexpression of specific GSTs can also affect chemoresistance (Hamada et al., 1994; Byun et al., 2005), whereas polymorphisms that decrease GST activity are associated with a high risk of developing cancer (Balendiran et al., 2004). In the present study, cisplatin-mediated decrease in GST activity in liver (Fig. 7a) may be a result of increase in free radical generation causing tissue toxicity. However, the increase in GST activity in kidney and testes (Fig. 7a) after combination of AA plus cisplatin treatment may indicate the utilization of GSH in detoxification reactions and reflecting the role of ascorbic acid in scavenging free radicals. An elevated expression of GSTs combined with high GSH levels can increase the rate of conjugation and detoxification of chemotherapy agents, thus reducing their effectiveness (Cullen et al., 2003). Increased GST activity in liver noted after combination of AA plus cisplatin treatment as compared to cisplatin alone (Fig. 7a) may imply GSTs to have peroxidase and isomerase activities which are involved in protection of cells against H2O2-induced cell death (Sheehan et al., 2001). Therefore, the observed increase in GST activity in the tissues may have a role in cellular defense mechanism.
Glutathione reductase (GR) function is also associated with an essential role in the cellular defense mechanism against oxidative stress. GR catalyses the reduction of oxidized glutathione (GSSG) to reduced glutathione (GSH) in presence of NADPH (Sweet and Blanchard, 1991). By generating GSH, GR indirectly participates in the protection of cells against oxidative stress and cytotoxic compounds and is deeply involved in the maintenance of the redox status of cells. Cisplatin-mediated decrease in GR activity observed in the tissues (particularly in liver) (Fig. 7b) may therefore hamper the cell’s ability to convert GSSG to GSH leading to GSSG accumulation within the cytosol, thereby weakening the cellular antioxidant machinery. However, the observed increase in GR activity in kidney after combination of AA plus cisplatin treatment could be in an attempt to maintain the intracellular ratio of reduced and oxidized GSH.
Catalase (CAT) is an endogenous antioxidant that prevents generation of hydroxyl radicals and protects the cellular constituents from oxidative damage (Hunt et al., 1998). Several evidences revealed that CAT activity is reduced/suppressed in cancerous tissues (Durak et al., 1994; Arruda et al., 1996). The decreased levels of CAT activity in liver, kidney and testes observed after cisplatin treatment (Fig. 7c) may result in increased production of ROS resulting in oxidative stress, which may also explain the cause of enhanced LPO (Fig. 5) as observed in the tissues of cisplatin alone treated mice in present study. The involvement of oxidative stress in cisplatin-induced toxicity may further be supported by the fact that many antioxidants have been reported to prevent cisplatin-induced nephrotoxicity (Ajith et al., 2007) and hepatotoxicity (Abouzeinab, 2013). Negahdar et al. (2005) reported that low levels of CAT activity in breast cancer resulted in higher production of ROS due to inadequate enzyme activity to detoxify high levels of hydrogen peroxide thereby, leading to formation of hydroxyl radicals. Low levels may be due to treatment by anticancer drugs which reduces antioxidants and induces oxidative stress which increases with disease progression (Borek, 2004). Catalase is implicated in destruction of harmful H2O2 generated in excess by different sub-cellular processes and by biotic and abiotic stresses. Induction of catalase activity provides protection against oxidant attacks created by cisplatin and/or its metabolites. The observed increase in CAT activities in the tissues after combination treatment (Fig. 7c) may also be involved to reduce lipid peroxides and play a compensatory regulatory role in decreasing cisplatin-induced oxidative stress and toxicities in the tissues and reflects the antioxidant role of ascorbic acid administration in the initial stages of tumor formation which may be useful as a secondary therapy to prevent the oxidative damage. These results imply that the increased activities of GR and CAT as well as increased concentration of reduced GSH in the tissues after combination treatment of AA plus cisplatin should be very helpful to decrease cisplatin-induced toxicities by elimination of excess hydrogen peroxide production and/or suppression of oxidative stress in the tissues of the host.
CONCLUSION
In conclusion, it may be suggested that cisplatin-induced mutagenicity and genotoxicity is closely associated with the increase of lipid peroxidation in the tissues. Treatment with cisplatin alone causes significant changes in the activity of antioxidant enzymes (GST, GR and CAT) and GSH concentration. Present findings show that pre-treatment with ascorbic acid in combination chemotherapy ameliorates cisplatin-induced oxidative stress and decreases alterations in antioxidant defense system and significantly decreases cisplatin-induced mutagenicity in the hosts.
ACKNOWLEDGMENT
The University Grants Commission, New Delhi, India is gratefully acknowledged for the award of Rajiv Gandhi National Fellowship granted to Ms. Amenla to carry out this research work.
REFERENCES
- Abouzeinab, N.S., 2013. Cytoprotective effect and antioxidant properties of silymarin on cisplatin induced hepatotoxicity in rats: A biochemical and histochemical study. Int. J. Cancer Res., 9: 9-23.
CrossRefDirect Link - Adler, I.D. and A. El-Tarras, 1989. Clastogenic effects of cis-diamminedichloroplatinum: I. Induction of chromosomal aberrations in somatic and germinal cells of mice. Mutat. Res./Fundam. Mol. Mechan. Mutagen., 211: 131-137.
CrossRefDirect Link - Aebi, H., 1984. Catalase in vitro. In: Methods in Enzymology, Packer, L., Academic Press, Cambridge, Massachusetts, United States, ISBN: 9780121820053, pp: 121-126.
CrossRefDirect Link - Ajith, T.A., S. Usha and V. Nivitha, 2007. Ascorbic acid and alpha-tocopherol protect anticancer drug cisplatin induced nephrotoxicity in mice: a comparative study. Clin. Chim. Acta, 375: 82-86.
CrossRefPubMedDirect Link - Antunes, L.M.G. and C.S. Takahashi, 1998. Effects of high doses of vitamins C and E against doxorubicin-induced chromosomal damage in Wistar rat bone marrow cells. Mutat. Res./Genet. Toxicol. Environ. Mutagen., 419: 137-143.
CrossRefPubMedDirect Link - Antunes, L.M.G., M.C.P. Araujo, J.D.C. Darin and M. D.L.P. Bianchi, 2000. Effects of the antioxidants curcumin and vitamin C on cisplatin-induced clastogenesis in Wistar rat bone marrow cells. Mutat. Res./Genet. Toxicol. Environ. Mutagen., 465: 131-137.
CrossRefPubMedDirect Link - Antunes, L.M.G., J.D.C. Darin and M.L.P. Bianchi, 2000. Protective effects of vitamin C against cisplatin-induced nephrotoxicity and lipid peroxidation in adult rats: A dose-dependent study. Pharmacol. Res., 41: 405-411.
CrossRefDirect Link - Atessahin, A., E. Sahna, G. Turk, A.O. Ceribasi, S. Yilmaz, A. Yuce and O. Bulmus, 2006. Chemoprotective effect of melatonin against cisplatin-induced testicular toxicity in rats. J. Pineal. Res., 41: 21-27.
CrossRefPubMedDirect Link - Bijur, G.N., M.E. Ariza, C.L. Hitchcock and M.V. Williams, 1997. Antimutagenic and promutagenic activity of ascorbic acid during oxidative stress. Environ. Mol. Mutag., 30: 339-345.
CrossRefDirect Link - Blasiak, J. and J. Kowalik, 2001. Protective action of vitamin C against DNA damage induced by selenium-cisplatin conjugate. Acta Biochim. Pol., 48: 233-240.
Direct Link - Borek, C., 2004. Dietary antioxidants and human cancer. Integr. Cancer Ther., 3: 333-341.
CrossRefPubMedDirect Link - Bruce, W.R., R. Furrer and A.J. Wyrobek, 1974. Abnormalities in the shape of murine sperm after acute testicular X-irradiation. Mutat. Res./Fundam. Mol. Mechan. Mutagen., 23: 381-386.
CrossRefPubMedDirect Link - Choudhury, R.C., M.B. Jagdale and S. Misra, 2000. Cytogenetic toxicity of cisplatin in bone marrow cells of Swiss mice. Chemother, 12: 173-182.
CrossRefDirect Link - Droge, W., 2002. Free radicals in the physiological control of cell function. Phsyol. Rev., 82: 47-95.
CrossRefPubMedDirect Link - Duthie, S.J., A. Ma, M.A. Ross and A.R. Collins, 1996. Antioxidant supplementation decreases oxidative DNA damage in human lymphocytes. Cancer Res., 56: 1291-1295.
PubMedDirect Link - Giri, A., D. Khynriam and S.B. Prasad, 1998. Vitamin C mediated protection on cisplatin induced mutagenicity in mice. Mutat. Res./Fundam. Mol. Mech. Mutagen., 421: 139-148.
CrossRefDirect Link - Habig, W.H., M.J. Pabst and W.B. Jakoby, 1974. Glutathione S-transferases: The first enzymatic step in mercapturic acid formation. J. Biol. Chem., 249: 7130-7139.
CrossRefPubMedDirect Link - Henson, D.E., G. Block and M. Levine, 1991. Ascorbic acid: Biologic functions and relation to cancer. J. Natl. Cancer Inst., 83: 547-550.
CrossRefPubMedDirect Link - Hsu, P.C., M.Y. Liu, C.C. Hsu, L.Y. Chen and Y.L. Guo, 1998. Effects of vitamin E and/or C on reactive oxygen species-related lead toxicity in the rat sperm. Toxicology, 128: 169-179.
CrossRefDirect Link - Kidd, P.M., 1997. Glutathione: Systemic protectant against oxidative and free radical damage. Altern. Med. Rev., 2: 155-176.
Direct Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Masuda, H., T. Tanaka and U. Takahama, 1994. Cisplatin generates superoxide anion by interaction with DNA in a cell-free system. Biochem. Biophys. Res Commun., 203: 1175-1180.
CrossRefPubMedDirect Link - McCall, M.R. and B. Frei, 1999. Can antioxidant vitamins materially reduce oxidative damage in humans? Free Radic. Biol. Med., 26: 1034-1053.
CrossRefDirect Link - Meister, A., 1994. Glutathione-ascorbic acid antioxidant system in animals. J. Biol. Chem., 269: 9397-9400.
Direct Link - Naziroglu, M., A. Karaoglu and A.O. Aksoy, 2004. Selenium and high dose vitamin E administration protects cisplatin-induced oxidative damage to renal, liver and lens tissues in rats. Toxicology, 195: 221-230.
CrossRefPubMedDirect Link - Nefic, H., 2001. Anticlastogenic effect of vitamin C on cisplatin induced chromosome aberrations in human lymphocyte cultures. Mutat. Res./Fundam. Mol. Mechan. Mutagen., 498: 89-98.
CrossRefDirect Link - Nicol, B.M. and S.B. Prasad, 2006. The effects of cyclophosphamide alone and in combination with ascorbic acid against murine ascites Dalton's lymphoma. Indian J. Pharmacol., 38: 260-265.
Direct Link - Odin, A.P., 1997. Vitamins as antimutagens: Advantages and some possible mechanisms of antimutagenic action. Mutat. Res./Rev. Mutat. Res., 386: 39-67.
CrossRefDirect Link - Rosenberg, B., 1985. Charles F. Kettring prize. Fundamental studies with cisplatin. Cancer, 55: 2303-2316.
CrossRefDirect Link - Sawhney, P., C.J. Giammona, M.L. Meistrich and J.H. Richburg, 2005. Cisplatin-induced long-term failure of spermatogenesis in adult C57/Bl/6J mice. J. Androl., 26: 136-145.
PubMedDirect Link - Sedlak, J. and R.H. Lindsay, 1968. Estimation of total, protein-bound, and nonprotein sulfhydryl groups in tissue with Ellman's reagent. Anal. Biochem., 25: 192-205.
CrossRefPubMedDirect Link - Shamberger, R.J., 1984. Genetic toxicology of ascorbic acid. Mutat. Res./Rev. Genet. Toxicol., 133: 135-159.
CrossRefDirect Link - Sheehan, D., G. Meade, V.M. Foley and C.A. Dowd, 2001. Structure, function and evolution of glutathione transferases: Implications for classification of non-mammalian members of an ancient enzyme superfamily. Biochem. J., 360: 1-16.
CrossRefPubMedDirect Link - Siddique, Y.H., T. Beg and M. Afzal, 2005. Antigenotoxic effects of ascorbic acid against megestrol acetate-induced genotoxicity in mice. Hum. Exp. Toxicol., 24: 121-127.
CrossRefPubMedDirect Link - Silva, C.R., L.M.G. Antunes and M.L.P. Bianchi, 2001. Antioxidant action of bixin against cisplatin-induced chromosome aberrations and lipid peroxidation in rats. Pharmacol. Res., 43: 561-566.
CrossRefPubMedDirect Link - Somani, S.M., K. Husain, C. Whitworth, G.L. Trammell, M. Malafa and L.P. Rybak, 2000. Dose-dependent protection by lipoic acid against cisplatin-induced nephrotoxicity in rats: Antioxidant defense system. Pharmacol. Toxicol., 86: 234-241.
CrossRefPubMedDirect Link - Umegaki, K., H. Uramoto and T. Esashi, 1997. Lack of influence of a long-term high or low vitamin E diet on the oxidative DNA damage in the bone marrow of mice. Int. J. Vitamin Nutr. Res., 67: 149-154.
PubMedDirect Link - Verrax, J. and P.B. Calderon, 2008. The controversial place of vitamin c in cancer treatment. Biochem. Pharmacol., 76: 1644-1652.
CrossRefDirect Link - Viswanatha Swamy, A.H.M., U. Wangikar, B.C. Koti, A.H.M. Thippeswamy, P.M. Ronad and D.V. Manjula, 2011. Cardioprotective effect of ascorbic acid on doxorubicin-induced myocardial toxicity in rats. Indian J. Pharmacol., 43: 507-511.
CrossRefDirect Link - Wyrobek, A.J., L.A. Gordon, J.G. Burkhart, M.W. Francis and R.W. Kapp et al., 1983. An evaluation of the mouse sperm morphology test and other sperm tests in nonhuman mammals: A report of the U.S. environmental protection agency Gene-Tox program. Mutat. Res./Rev. Genet. Toxicol., 115: 1-72.
CrossRefPubMedDirect Link - Wyrobek, A.J. and W.R. Bruce, 1975. Chemical induction of sperm abnormalities in mice. Proc. Natl. Acad. Sci., 72: 4425-4429.
CrossRefPubMedDirect Link - Al-Hakkak, Z.S., H.A. Hamamy, A.M.B. Murad and A.F. Hussain, 1986. Chromosome aberrations in workers at a storage battery plant in Iraq. Mutat. Res. Genet. Toxicol., 171: 53-60.
CrossRefDirect Link - Amenla, L., A.K. Verma and S.B. Prasad, 2013. Dietary ascorbic acid-mediated augmentation of antitumor activity and protection against toxicities induced by cis-diamminedichloroplatinum-(II) in Dalton's Lymphoma-Bearing mice. J. Cancer Res. Updates, 2: 116-130.
Direct Link - Antunes, L.M.G., J.D.C. Darin and M.D.L.P. Bianchi, 1999. Anticlastogenic effect of vitamin C on cisplatin in vivo. Genet. Mol. Biol., 22: 415-417.
CrossRefDirect Link - Arrick, B.A. and C.F. Nathan, 1984. Glutathione metabolism as a determinant of therapeutic efficacy: A review. Cancer Res., 44: 4224-4232.
PubMedDirect Link - Arruda, V.R., T.S. Salles, F.F. Costa and S.T. Saad, 1996. Glutathione peroxidase, reduced glutathione, superoxide dismutase and catalase in red cells of patients with hairy cell leukemia. Neoplasma, 43: 99-102.
PubMed - Attia, S.M., 2010. The impact of quercetin on cisplatin-induced clastogenesis and apoptosis in murine marrow cells. Mutagenesis, 25: 281-288.
CrossRefPubMedDirect Link - Barbosa, A.L.R., C.A. Pinheiro, G.J. Oliveira, M.O. Moraes, R.A. Ribeiro, M.L. Vale and M.H.L.P. Souza, 2009. Tumor bearing decreases systemic acute inflammation in rats-role of mast cell degranulation. Inflamm. Res., 58: 235-240.
CrossRefPubMedDirect Link - Balendiran, G.K., R. Dabur and D. Fraser, 2004. The role of glutathione in cancer. Cell Biochem. Funct., 22: 343-352.
PubMedDirect Link - Bench, G., M.H. Corzett, R. Martinelli and R. Balhorn, 1999. Cadmium concentrations in the testes, sperm and spermatids of mice subjected to long-term cadmium chloride exposure. Cytometry, 35: 30-36.
CrossRefPubMedDirect Link - Brozovic, G., N. Orsolic, F. Knezevic, A.H. Knezevic and V. Benkovic et al., 2011. The in vivo genotoxicity of cisplatin, isoflurane and halothane evaluated by alkaline comet assay in Swiss albino mice. J. Applied Genet., 52: 355-361.
CrossRefPubMedDirect Link - Buege, J.A. and S.D. Aust, 1978. Microsomal Lipid, Peroxidation. In: Methods in Enzymology, Vol. 52, Flesicher, S. and L. Packer (Eds.)., Academic Press, New York, pp: 302-310.
CrossRefDirect Link - Byun, S.S., S.W. Kim, H. Choi, C. Lee and E. Lee, 2005. Augmentation of cisplatin sensitivity in cisplatin-resistant human bladder cancer cells by modulating glutathione concentrations and glutathione-related enzyme activities. BJU Int., 95: 1086-1090.
CrossRefPubMedDirect Link - Cabrera, G., 2000. Effect of five dietary antimutagens on the genotoxicity of six mutagens in the microscreen prophage-induction assay. Environ. Mol. Mutagen., 36: 206-220.
CrossRefPubMedDirect Link - Chen, G., K.J. Hutter and W.J. Zeller, 1995. Positive correlation between cellular glutathione and acquired cisplatin resistance in human ovarian cancer cells. Cell Biol. Toxicol., 11: 273-281.
CrossRefPubMedDirect Link - Cullen, K.J., K.A. Newkirk, L.M. Schumaker, N. Aldosari, J.D. Rone and B.R. Haddad, 2003. Glutathione S-transferase π amplification is associated with cisplatin resistance in head and neck squamous cell carcinoma cell lines and primary tumors. Cancer Res., 63: 8097-8102.
Direct Link - Dos Santos, G.C., L.M. Mendonca, G.A. Antonucci, A.C. dos Santos, L.M.G. Antunes and M.L.P. Bianchi, 2012. Protective effect of bixin on cisplatin-induced genotoxicity in PC12 cells. Food Chem. Toxicol., 50: 335-340.
CrossRefDirect Link - Durak, I., O. Akyol, E. Basesme, O. Canbolat and M. Kavutcu, 1994. Reduced erythrocyte defense mechanisms against free radical toxicity in patients with chronic renal failure. Nephron, 66: 76-80.
CrossRefDirect Link - Edelweiss, M.I., A. Trachtenberg, E.X. Pinheiro, J. da-Silva, M. Riegel, H.M. Lizardo-Daudt and M.S. Mattevi, 1995. Clastogenic effect of cisplatin on Wistar rat bone marrow cells. Braz. J. Med. Biol. Res., 28: 679-683.
PubMedDirect Link - El-Refaiy, A.I. and F.I. Eissa, 2012. Protective effects of ascorbic acid and zinc against cadmium-induced histopathological, histochemical and cytogenetic changes in rats. Comunicata Scientiae, 3: 162-180.
Direct Link - Elsendoorn, T.J., N.I. Weijl, S. Mithoe, A.H. Zwinderman and F. Van Dam et al., 2001. Chemotherapy-induced chromosomal damage in peripheral blood lymphocytes of cancer patients supplemented with antioxidants or placebo. Mutat. Res./Genet. Toxicol. Environ. Mutagen., 498: 145-158.
CrossRefDirect Link - Florea, A.M. and D. Busselberg, 2011. Cisplatin as an anti-tumor drug: Cellular mechanisms of activity, drug resistance and induced side effects. Cancers, 3: 1351-1371.
CrossRefDirect Link - Fox, M.R.S., 1975. Protective effects of ascorbic acid against toxicity of heavy metals. Ann. N.Y. Acad. Sci., 258: 144-150.
CrossRefDirect Link - Frezza, M., S. Hindo, D. Chen, A. Davenport, S. Schmitt, D. Tomco and Q.P. Dou, 2010. Novel metals and metal complexes as platforms for cancer therapy. Curr. Pharmaceut. Des., 16: 1813-1825.
CrossRefDirect Link - Garcia-Sagredo, J.M., 2008. Fifty years of cytogenetics: A parallel view of the evolution of cytogenetics and genotoxicology. Biochim. Biophys. Acta (BBA)-Gene Regul. Mechan., 1779: 363-375.
CrossRefDirect Link - Gentile, J.M., S. Rahimi, J. Zwiesler, G.J. Gentile and L.R. Ferguson, 1998. Effect of selected antimutagens on the genotoxicity of antitumor agents. Mutat. Res./Fundam. Mol. Mech. Mutagen., 402: 289-298.
CrossRefDirect Link - Giri, S., A. Giri, G.D. Sharma and S.B. Prasad, 2002. Mutagenic effects of carbosulfan, a carbamate pesticide. Mutat. Res./Genet. Toxicol. Environ. Mutagen., 519: 75-82.
CrossRefDirect Link - Griffiths, H.R. and J. Lunec, 2001. Ascorbic acid in the 21st century-more than a simple antioxidant. Environ. Toxicol. Pharmacol., 10: 173-182.
CrossRefDirect Link - Hamada, S.I., M. Kamada, H. Furumoto, T. Hirao and T. Aono, 1994. Expression of glutathione S-transferase-π in human ovarian cancer as an indicator of resistance to chemotherapy. Gynecol. Oncol., 52: 313-319.
CrossRefDirect Link - Hrubisko, M., A.T. McGown and B.W. Fox, 1993. The role of metallothionein, glutathione, glutathione S-transferases and DNA repair in resistance to platinum drugs in a series of L1210 cell lines made resistant to anticancer platinum agents. Biochem. Pharmacol., 45: 253-256.
CrossRefDirect Link - Hunt, C.R., J.E. Sim, S.J. Sullivan, T. Featherstone and W. Golden et al., 1998. Genomic instability and catalase gene amplification induced by chronic exposure to oxidative stress. Cancer Res., 58: 3986-3992.
Direct Link - Husain, K., C. Morris, C. Whitworth, G.L. Trammell, L.P. Rybak and S.M. Somani, 1998. Protection by ebselen against cisplatin-induced nephrotoxicity: Antioxidant system. Mol. Cell. Biochem., 178: 127-133.
CrossRefDirect Link - Jain, A., J. Martensson, T. Mehta, A.N. Krauss, P.A. Auld and A. Meister, 1992. Ascorbic acid prevents oxidative stress in glutathione-deficient mice: effects on lung type 2 cell lamellar bodies, lung surfactant and skeletal muscle. Proc. Natl. Acad. Sci., 89: 5093-5097.
Direct Link - Kalita, S., A.K. Verma and S.B. Prasad, 2014. Chlorambucil and ascorbic acid-mediated anticancer activity and hematological toxicity in Dalton's ascites lymphoma-bearing mice. Indian J. Exp. Biol., 52: 112-124.
Direct Link - Kartalou, M. and J.M. Essigmann, 2001. Mechanisms of resistance to cisplatin. Mutat. Res./Fundam. Mol. Mech. Mutagen., 478: 23-43.
CrossRefDirect Link - Kliesch, U. and I.D. Adler, 1987. Micronucleus test in bone marrow of mice treated with 1-nitropropane, 2-nitropropane and cisplatin. Mutat. Res. Lett., 192: 181-184.
CrossRefDirect Link - Krishnaja, A.P. and N.K. Sharma, 2003. Ascorbic acid potentiates mitomycin C-induced micronuclei and sister chromatid exchanges in human peripheral blood lymphocytes in vitro. Teratogen. Carcinogen. Mutagen., 23: 99-112.
CrossRefDirect Link - Longchar, A. and S.B. Prasad, 2015. Biochemical changes associated with ascorbic acid-cisplatin combination therapeutic efficacy and protective effect on cisplatin-induced toxicity in tumor-bearing mice. Toxicol. Rep., 2: 489-503.
CrossRefDirect Link - Meijer, C., N.H. Mulder, G.A. Hospers, D.R. Uges and E.G. de Vries, 1990. The role of glutathione in resistance to cisplatin in a human small cell lung cancer cell line. Br. J. Cancer, 62: 72-77.
Direct Link - Misra, S. and R.C. Choudhury, 2006. Vitamin C modulation of cisplatin-induced cytogenotoxicity in bone marrow, spermatogonia and its transmission in the male germline of Swiss mice. J. Chemother., 18: 182-187.
CrossRefDirect Link - Mitazaki, S., M. Hashimoto, Y. Matsuhashi, S. Honma and M. Suto et al., 2013. Interleukin-6 modulates oxidative stress produced during the development of cisplatin nephrotoxicity. Life Sci., 92: 694-700.
CrossRefDirect Link - Mohammadnejad, D., A. Abedelahi, J. Soleimani-Rad, A. Mohammadi-Roshandeh, M. Rashtbar and A. Azami, 2012. Degenerative effect of cisplatin on testicular germinal epithelium. Adv. Pharmaceut. Bull., 2: 173-177.
CrossRefDirect Link - Nagyova, A. and E. Ginter, 1995. The influence of ascorbic acid on the hepatic cytochrome P-450 and glutathione in guinea-pigs exposed to 2,4-dichlorophenol. Physiol. Res., 44: 301-305.
PubMedDirect Link - Nagyova, A. and E. Ginter, 1994. Interactions between hepatic ascorbic acid, cytochrome P-450 and lipids in female guinea pigs with different ascorbic acid intake. Physiol. Res., 43: 307-312.
PubMedDirect Link - Nefic, H., 2008. The genotoxicity of vitamin C in vitro. Bosnian J. Basic Med. Sci., 8: 141-146.
PubMedDirect Link - Negahdar, M., M. Djalali, H. Abtahi, M.R. Sadeghi, T. Aghvami, E. Javadi and H. Layegh, 2005. Blood superoxide dismutase and catalase activities in women affected with breast cancer. Iran. J. Publ. Health, 34: 39-43.
Direct Link - Ognjanovic, B.I., N.Z. Djordjevic, M.M. Matic, J.M. Obradovic, J.M. Mladenovic, A.S. Stajn and Z.S. Saicic, 2012. Lipid peroxidative damage on cisplatin exposure and alterations in antioxidant defense system in rat kidneys: A possible protective effect of selenium. Int. J. Mol. Sci., 13: 1790-1803.
CrossRefDirect Link - Osanto, S., J.C.P. Thijssen, V.M. Woldering, J.L.S. van Rijn, A.T. Natarajan, A.D. Tates and A.V. Carrano, 1991. Increased frequency of chromosomal damage in peripheral blood lymphocytes up to nine years following curative chemotherapy of patients with testicular carcinoma. Environ. Mol. Mutagen., 17: 71-78.
CrossRefPubMedDirect Link - Park, J.H., K.R. Davis, G. Lee, M. Jung and Y. Jung et al., 2012. Ascorbic acid alleviates toxicity of paclitaxel without interfering with the anticancer efficacy in mice. Nutr. Res., 32: 873-883.
CrossRefPubMedDirect Link - Pereira, C.F. and C.R. de Oliveira, 2000. Oxidative glutamate toxicity involves mitochondrial dysfunction and perturbation of intracellular Ca2+ homeostasis. Neurosci. Res., 37: 227-236.
CrossRefDirect Link - Poirier, M.C., E. Reed, C.L. Litterst, D. Katz and S. Gupta-Burt, 1992. Persistence of platinum-ammine-DNA adducts in gonads and kidneys of rats and multiple tissues from cancer patients. Cancer Res., 52: 149-153.
Direct Link - Ramadan, L.A., O.H. El-Habit, H. Arafa and M.M. Sayed-Ahmad, 2001. Effect of cremophorel on cisplatin-induced organ toxicity in normal rat. J. Egypt. Nat. Cancer Inst., 13: 139-145.
Direct Link - Seetharama Rao, K.P. and K. Narayana, 2005. In vivo chromosome damaging effects of an inosine monophosphate dehydrogenase inhibitor: Ribavirin in mice. Indian J. Pharmacol., 37: 90-95.
CrossRefDirect Link - Rosselli, F., L. Zaccaro, M. Venturi and A.M. Rossi, 1990. Persistence of drug-induced chromosome aberrations in peripheral blood lymophocytes of the rat. Mutat. Res./Fundam. Mol. Mechan. Mutagen., 232: 107-114.
CrossRefPubMedDirect Link - Roy, N.K. and T.G. Rossman, 1992. Mutagenesis and comutagenesis by lead compounds. Mutat. Res./Genet. Toxicol,, 298: 97-103.
CrossRefDirect Link - Sainath, S.B., T. Sowbhagyamma, P. Madhu, K.P. Reddy, B.P. Girish and P.S. Reddy, 2011. Protective Effect of Speman on Cisplatin-Induced Testicular and Epididymal Toxicity in Mice. Int. J. Green Pharm., 5: 286-291.
Direct Link - Smith, I.K., T.L. Vierheller and C.A. Thorne, 1988. Assay of glutathione reductase in crude tissue homogenates using 5,5'-dithiobis(2-nitrobenzoic acid). Anal. Biochem., 175: 408-413.
CrossRefPubMedDirect Link - Sorenson, C.M. and A. Eastman, 1988. Mechanism of cis-diamminedichloroplatinum(II)-induced cytotoxicity: Role of G2 arrest and DNA double-strand breaks. Cancer Res., 48: 4484-4488.
Direct Link - Sweet, W.L. and J.S. Blanchard, 1991. Human erythrocyte glutathione reductase: Chemical mechanism and structure of the transition state for hydride transfer. Biochemistry, 30: 8702-8709.
CrossRefDirect Link - Terradas, M., M. Martin, L. Tusell and A. Genesca, 2010. Genetic activities in micronuclei: Is the DNA entrapped in micronuclei lost for the cell? Mutat. Res., 705: 60-67.
CrossRefDirect Link - Townsend, D.M. and K.D. Tew, 2003. The role of glutathione-S-transferase in anti-cancer drug resistance. Oncogene, 22: 7369-7375.
CrossRefDirect Link - Trbojevic, I.S., B.I. Ognjanovic, N.Z. Ordevic, S.D. Markovic, A.S. Stajn, J.P. Gavric and Z.S. Saicic, 2010. Effects of cisplatin on lipid peroxidation and the glutathione redox status in the liver of male rats: The protective role of selenium. Arch. Boil. Sci. Belgrade, 62: 75-82.
CrossRefDirect Link - Ushijima, C., Y. Kumasako, P.E. Kihaile, K. Hirotsuru and T. Utsunomiya, 2000. Analysis of chromosomal abnormalities in human spermatozoa using multi-colour fluorescence in-situ hybridization. Hum. Reprod., 15: 1107-1111.
CrossRefDirect Link - Yedjou, C., L. Thuisseu, C. Tchounwou, M. Gomes, C. Howard and P. Tchounwou, 2009. Ascorbic acid potentiation of arsenic trioxide anticancer activity against acute promyelocytic leukemia. Arch. Drug Inform., 2: 59 -65.
CrossRefDirect Link - Ayinde, O.C., S. Ogunnowo and R.A. Ogedegbe, 2012. Influence of vitamin C and vitamin E on testicular zinc content and testicular toxicity in lead exposed albino rats. BMC Pharmacol. Toxicol., 13: 17-24.
CrossRefDirect Link