
Ujomonigho E. Omoregie
Department of Plant Biology and Biotechnology, University of Benin, Nigeria
Joseph K. Mensah
Department of Botany, Ambrose Alli University, Ekpoma, Nigeria
Beckley Ikhajiagbe
Department of Plant Biology and Biotechnology, University of Benin, Nigeria
Research Journal of Mutagenesis
Year: 2014 | Volume: 4 | Issue: 1 | Page No.: 14-22







ABSTRACT
The present study investigated the effects of sodium azide pretreatment of grains on the germination of five varieties of rice. Germination parameters of FARO 44, FARO 52, FARO 57, NERICA L-34 and NERICA L-47 rice varieties treated with different concentrations of sodium azide was studied in petri dish experiments. Lethal dose (LD50) was estimated at 0.032% sodium azide. The concentrations of sodium azide solution (0.004, 0.008, 0.016 and 0.032% sodium azide at pH 3) were prepared on volume/weight basis. Control was distilled water at pH 7. Viable grains were pretreated in sodium azide solutions for 6 h. Germination was monitored from the second to fourteenth Day after Initiation (DAI). There were significant differences (p = 0.05) in percentage germination, length of radicles, length of plumules and dry weights of sprouting grains. Germination percentage on the fourth day after initiation was 0% in FARO 57 and NERICA L-47 pre-treated in 0.032% sodium azide. At 14 DAI, germination percentage in all varieties including the control ranged between 94 and 100%. The longest and shortest radicles were both obtained in NERICA L-47 (86.25, 2.50 mm) in 0.032 and 0%, respectively. The shortest plumules (1.00 mm) were obtained in both NERICA L-34 and NERICA L-47, pre-treated in 0.032% sodium azide with and longest (50.75 mm) in 0% sodium azide pre-treated NERICA L-47 at 6 DAI. The experiment showed that sodium azide significantly delayed but did not inhibit germination.
PDF Abstract XML References Citation
Received: July 22, 2013;
Accepted: October 04, 2013;
Published: April 15, 2014
How to cite this article
Ujomonigho E. Omoregie, Joseph K. Mensah and Beckley Ikhajiagbe, 2014. Germination Response of Five Rice Varieties Treated with Sodium Azide. Research Journal of Mutagenesis, 4: 14-22.
DOI: 10.3923/rjmutag.2014.14.22
URL: https://scialert.net/abstract/?doi=rjmutag.2014.14.22
DOI: 10.3923/rjmutag.2014.14.22
URL: https://scialert.net/abstract/?doi=rjmutag.2014.14.22
INTRODUCTION
Mutations in any population result in genetic variability and this is the basis for breeding for new varieties within a population (Kleinhofs et al., 1978). They either occur naturally or can be induced by physical, biological and chemical agents.
Induced chemical mutations can either be advantageous or disadvantageous in living organisms. Reactive oxygen species produced by these chemicals are chromosome damaging in plants (Yuan and Zhang, 1993). They change amino acids by inducing base pair substitutions, especially GC→AT. These changes, compared to deletions or frame shift mutations interfere with the function of proteins, but do not inactivate them (Van der Veen, 1966). A wide range of morphological and yield parameters have been influenced by these chemical mutagens. By so doing, these mutants help to facilitate the characterization and cloning of those genes that are used in designing crops with improved desired traits (Ahloowalia and Maluszynski, 2001). An example of a chemical mutagen is sodium azide (Al-Qurainy and Khan, 2009).
It has been reported that sodium azide has been used to improve yield traits in groundnut (Mensah and Obadoni, 2007), enhance amylase content in rice (Suzuki et al., 2008) and to impact resistance against Striga in maize (Kiruki et al., 2006). Mutagenicity is mainly mediated by the hydrazoic acid produced by sodium azide (Owais and Kleinhofs, 1988). Hydrazoic acids causes point mutation in the organism’s genome upon entry into the nucleus (Al-Qurainy and Khan, 2009).
The direct effect of sodium azide on germination is the first indication of its activity. This is very important step for breeding studies since not all organisms respond to sodium azide treatment (Gichner and Veleminsky, 1977; Kamra and Gollapudi, 1979). With higher doses, the mutagenic efficiency is higher thus improving the chances of more cells to be exposed to mutation, however at a lethal dose there is no germination. That dose has to be determined for the particular species before any induced mutation breeding work can progress. At lower pH values, greater quantities of hydrazonic acid are metabolized from sodium azide which is the compound that penetrates the cell membrane. The effects of hydrazoic acid is higher at pH 3 that at pH 6 (Kleinhofs et al., 1974; Niknejad, 1975). The present study therefore attempts to investigate the effect sodium azide concentrations in rice germination.
The world all over, rice has been known to be the second most-consumed cereal grain (Crawford and Shen, 1998). With the ever-changing climate, there seem to exist a vast range of genetic variability of crops of economic in order to adapt to extreme weather conditions. One way of creating these variabilities is through chemical mutagens like sodium azide.
The rice varieties selected for this study were FARO 44, FARO 52, FARO 57, NERICA L-34 and NERICA L-47. The grains were obtained from the National Cereals Research Institute, Badeggi, Bida, Nigeria. They are among the rain-fed lowland mega rice cultivars in Nigeria (Akinwale et al., 2012). The Emergency Rice Initiative Project promoted these popular rice varieties in Nigeria (Maji, 2009). The varieties are high yielding, early maturing, shallow-swamp varieties that have the potential to be bred to withstand salinity and iron toxicity challenges in the shallow swamp and mangrove environments of which rice is a good candidate, converting such areas into arable swamps. These varieties were conventionally bred for the traits they express. Conventional breeding is however an indeed slow process.
Induced mutations promise a quicker alternative in creating genetic variability in plants to enable them adapt to the rapidly changing environmental conditions.
The aim of this study was to observe the potency of sodium azide as a mutagenic agent in these high-yielding rice varieties.
MATERIALS AND METHODS
Preparation of sodium azide (NaN3) solutions: The first part of the experiment was used to test for the Lethal dose 50 (LD50); the dose at which at least 50% of grains will germinate. Seeds were treated with 1.000, 0.2500, 0.0625 and 0.0320% NaN3 concentrations, prepared by weighing 1.000, 0.2500, 0.0625 and 0.0320 g, respectively and dissolving each in 100 mL of distilled water pre-regulated to pH 3 with ortho-phosphoric acid.
After LD50 was determined, reduced concentrations of 0.032, 0.016, 0.008 and 0.004% w/v were prepared by measuring appropriate volumes from stock solution and making up to 100 mL with ortho-phosphoric acid-regulated distilled water (pH 3). The 0% treatment was acid-regulated distilled water at pH 3 with no NaN3. Control was only distilled water at neutral pH.
Grain treatments: Uniform grains, free from insect attack were selected by hand-picking and transferred into labelled Petri-dishes. Grains were pre-soaked in distilled water (pH 7) for 14 h and then transferred to NaN3 (pH 3) solutions for 6 h, with continuous stirring. At the end of the exposure to NaN3 treatment, grains were properly rinsed in water.
Percentage germination: Fifty seeds were germinated in 9 cm sterile Petri dishes lined with one sterile Whatman No. 1 filter paper with 5 mL of distilled water (pH 7). Numbers of grains from which radicles emerged were counted daily up until 14 DAI. Germination in most seeds is initiated from the moment they make contact with sufficient water (Bradford, 1990). Germination tests were conducted under conditions of a 12 h light/dark cycle at 25°C. About 10 drops of distilled water were added into the Petri dishes every day to maintain moist condition to support germination. Percentage germination was then calculated by:
![]()
Vegetative parameters: The lengths of radicles and plumules of four selected germinating grains were measured daily on up till 6 DAI with a measuring tape. One hundred grains of sprouted grains were first weighed fresh and then the grains were dried in a hot-air oven at 60°C for 72 h and the dry weights taken.
Experimental design and statistics: The experimental design was a completely randomized design with four replicates. Means and standard errors were calculated from data obtained. Data were analyzed following two-way analysis of variance using GENSTAT (8th edition) statistical software package. Where significant F-values were obtained, differences between means were separated using Student Newman Keuls test (Alika, 2006).
RESULTS
Germination at 2 DAI was fastest in the control experiments than in the NaN3-treated rice varieties. Germination percentage declined as sodium azide treatment increased. Germination was slowest in 0.032% NaN3-treated NERICA L-47, starting at 5 DAI. At 14 DAI however, there were no significant differences in the percentage germination obtained for all the varieties irrespective of sodium azide pre-treatment. Percentage germination ranged between 94 and 100% across treatments in all varieties (Fig. 1).
At 2 DAI, the NERICA controls had radicle lengths of 4.75 mm each. There was no radicle formation at 2 DAI in the FARO 52, FARO 57 and the NERICA varieties that were pre-treated in 0.016 and 0.032% NaN3. Radicles were shorter as sodium azide treatment increased across varieties (Table 1). Radicle length in FARO 44 control and 0.032% NaN3 at 5 DAI were 56.00 and 2.75 mm, respectively. NERICA L-34 was 52.50 and 1.00 mm in control and 0.032% NaN3, respectively at 5 DAI.
At 6 DAI, plumule length was least (1.00 mm) in the NERICA varieties pre-treated in 0.032% NaN3 solution, followed by 0.032% NaN3 pre-treated FARO 57 (3.75 mm). Generally, high concentrations of NaN3 in solutions significantly reduced plumule length (Table 2). At 5 DAI, FARO 44 had 24.50 mm plumule length in its control and 2.50 mm plumule length in 0.032% NaN3. Also NERICA L-47 was 29.25 mm in its control with no plumule formation at 0.032% NaN3.
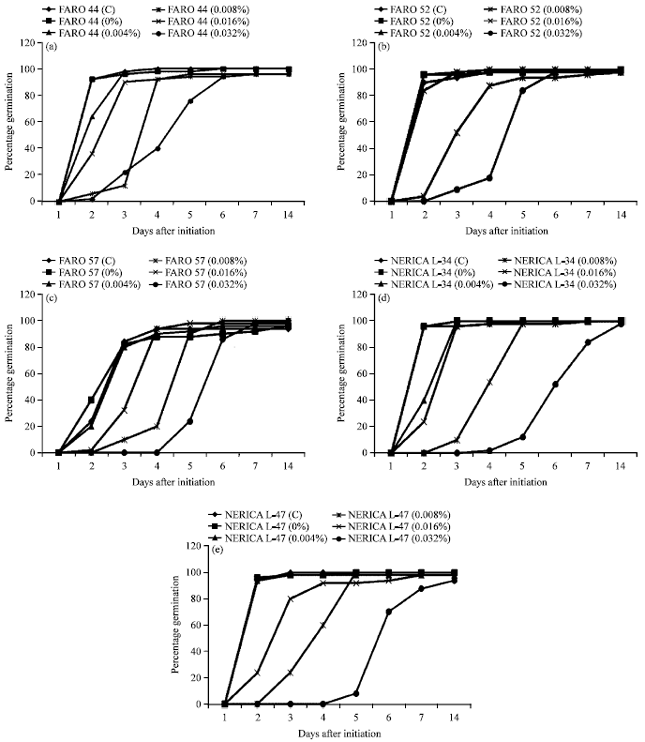 | |
| Fig. 1(a-e): | Percentage germination in the five rice varieties studied, (a) Percentage germination in FARO 44 at different sodium azide concentrations, (b) Percentage germination in FARO 52 at different sodium azide concentrations, (c) Percentage germination in FARO 57 at different sodium azide concentrations, (d) Percentage germination in NERICA L-34 at different sodium azide concentrations and (e) Percentage germination in NERICA L-47 at different sodium azide concentrations |
| Table 1: | Daily changes in lengths of radicles (mm) of the five rice varieties treated with sodium azide solutions |
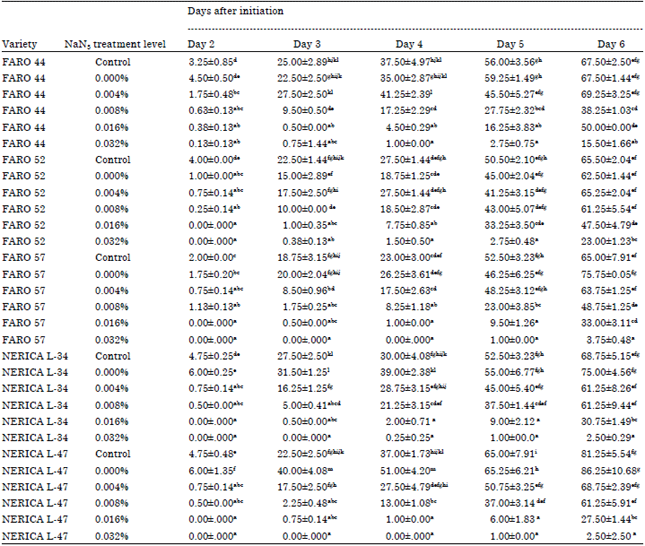 | |
| Mean±Standard error. Mean of four replicates. Different alphabets in columns indicate significant differences at 5% level of probability | |
Fresh weights of sprouting grains ranged from 3.51 g in the 0.032% NaN3-treated NERICA L-47, to 6.82 g in the untreated NERICA L-47 variety. Dry weights ranged from 2.02-2.63 g (Table 3).
DISCUSSION
Percentage germination was significantly delayed as sodium azide concentration increased in all varieties (Fig. 1). Germination was delayed more in FARO 57 and the NERICA varieties at the highest sodium azide treatment than in both FARO 44 and 52. Cheng and Gao (1988) treated barley seeds with sodium azide and recorded significant decreases in percentage germination. Pearson et al. (1974, 1975) observed delay in the initiation of plant growth with treatments of sodium azide, under the same conditions. Mostafa (2011) reported slight decrease in germination of sun flower seeds with high treatments of sodium azide. Khan et al. (2004, 2005) also reported decreases in germination in chick pea and Mung bean. Sinha and Lai (2007) also noticed decrease in lentil germination as a result of sodium azide treatment. These effects are as a result of delay or inhibitions in physiological and biological processes for germination which include enzyme activity, thereby impeding ATP synthesis in the affected cells, slowing down germination rate and germination percentage.
| Table 2: | Daily changes in lengths of plumules (mm) of the five rice varieties treated with sodium azide solutions |
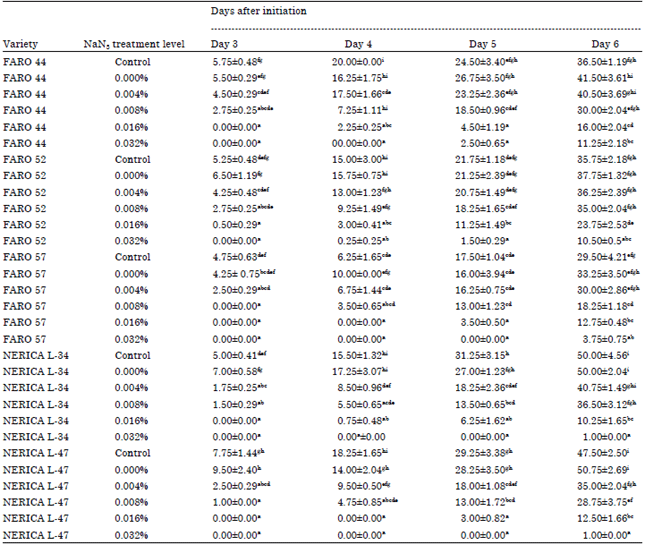 | |
| Mean±Standard error. Mean of four replicates. Different alphabets in columns indicate significant differences at 5% level of probability | |
However, some seeds develop tolerance to the inhibitory effects of sodium azide on germination on additional days to the physiological effects with respect to germination (Al-Qurainy and Khan, 2009). The reversible inhibitory effects of sodium azide on most divalent ion-containing enzymes account mostly for its toxicity (Kleinhofs et al., 1978). Kleinhofs et al. (1978) recorded that 0.003 M sodium azide increased mutations in pea. However, higher sodium azide doses caused disturbance in genetic and physiological activities leading to death of cells. In this study, lethal doses for the rice varieties were concentrations above 0.032% sodium azide.
At 14 DAI, there was no significant difference in the germination percentages irrespective of sodium azide treatments. It is very possible that on additional days the seeds, in this case, may have developed some level of tolerance to the inhibitory effects of sodium azide (Al-Qurainy and Khan, 2009).
Length of radicles and plumules decreased as sodium azide concentration increased this however could be as result of delays in germination (Table 1 and 2).
| Table 3: | Fresh and dry weights of sprouting grains of the five rice varieties (g) at fourteen days after initiation |
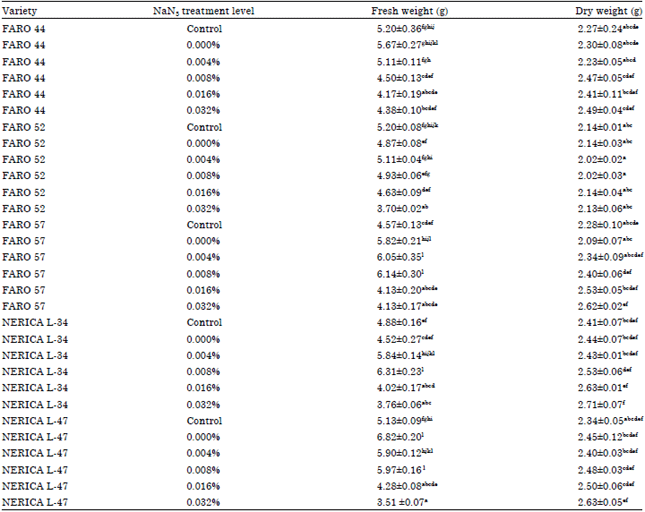 | |
| Mean±standard error. Mean of four replicates. Different alphabets in columns indicate significant differences at 5% level of probability | |
Lal et al. (2009) previously reported marked decrease in seedling height at high concentrations of mutagens. Singh and Yadav (1987) also established that reduction in seedling height correlated with increasing concentrations of the mutagens.
Dry weights of sprouting grains were significantly higher in sodium azide-treated plants than their controls, except in FARO 52 (Table 3). Seed germination and seedling emergence have been described as the beginning of the life cycle of plants and is critical for the establishment of plant populations (Khan and Gulzar, 2003). After overcoming the initial sodium azide inhibition to germination, sodium azide-treated plants were observed to have accumulated more biomass than untreated plants, thereby improving their chances of survival and establishment.
CONCLUSION
This study reveals that sodium azide is a potent chemical mutagen in the rice varieties used for the experiment which corroborates its effects on other plants also studied by Researchers. It has also provided baseline information on sodium azide lethal dose for the varieties studied. The research can therefore proceed to study its effects on yield parameters and also identify and select positive mutations for further breeding experiments.
REFERENCES
- Ahloowalia, B.S. and M. Maluszynski, 2001. Induced mutations-A new paradigm in plant breeding. Euphytica, 118: 167-173.
CrossRefDirect Link - Al-Qurainy, F. and S. Khan, 2009. Mutagenic effects of sodium azide and its application in crop improvement. World Applied Sci. J., 6: 1589-1601.
Direct Link - Bradford, K.J., 1990. A water relations analysis of seed germination rates. Plant Physiol., 94: 840-849.
CrossRef - Cheng, X. and M. Gao, 1988. Biological and genetic effects of combined treatments of sodium azide, gamma rays and EMS in barley. Environ. Exp. Bot., 28: 281-288.
CrossRef - Crawford, G.W. and C. Shen, 1998. The origins of rice agriculture: Recent progress in East Asia. Antiquity, 72: 858-866.
CrossRefDirect Link - Gichner, T. and J. Veleminsky, 1977. The very low mutagenic activity of sodium azide in Arabidopsis thaliana. Biol. Plant., 19: 153-155.
CrossRefDirect Link - Kamra, O.P. and B. Gollapudi, 1979. Mutagenic effects of sodium azide in Drosophila melanogaster. Mutation Res./Genet. Toxicol., 25: 381-384.
CrossRefPubMedDirect Link - Khan, M.A. and S. Gulzar, 2003. Germination responses of Sporobolus ioclados: A saline desert grass. J. Arid Environ., 53: 387-394.
CrossRefDirect Link - Khan, S., M.R. Wani and K. Parveen, 2004. Induced genetic variability for quantitative traits in Vigna radiate (L.) Wilczek. Pak. J. Bot., 36: 845-850.
Direct Link - Kleinhofs, A., C. Sander, R.A. Nilian and C.F. Konzak, 1974. Azide Mutagenicity-Mechanism and Nature of Mutants Produced. In: Polyploidy and Induced Mutations in Plant Breeding: Proceedings of a Meeting on Mutation and Polyploidy and of a Research Co-Ordination Meeting on Improvement of Mutation Breeding Techniques, Held in Bari, October 2-10,1972, FAO and IAEA (Eds.). IAEA, Vienna, pp: 195-199.
- Kleinhofs, A., W.M. Owais and R.A. Nilan, 1978. Azide. Mutat. Res./Rev. Genet. Toxicol., 55: 165-195.
CrossRefDirect Link - Lal, G.M., B. Toms and S. Smith, 2009. Induced chlorophyll mutations in black gram. Asian J. Agric. Sci., 1: 1-3.
Direct Link - Mensah, J.K. and B. Obadoni, 2007. Effects of sodium azide on yield parameters of groundnut (Arachis hypogaea L.). Afr. J. Biotechnol., 6: 668-671.
Direct Link - Mostafa, G.G., 2011. Effect of sodium azide on the growth and variability induction in Helianthus annuus L. Int. J. Plant Breed. Genet., 5: 76-85.
CrossRefDirect Link - Niknejad, M., 1975. The Effect of Duration and Condition of Post-Treatment Storage on the Physiological Damage and Mutagenic Frequency of Barley Treated with Different Concentrations of Sodium Azide. In: Barley Genetics III, Gaul, H. (Ed.). Verlag Karl Thiemig, Munich, Germany, ISBN-13: 9783521061088, pp: 132-145.
- Owais, W.M. and A. Kleinhofs, 1988. Metabolic activation of the mutagen azide in biological systems. Fundam. Mol. Mechanisms Mutagenesis, 197: 313-323.
CrossRefPubMedDirect Link - Pearson, O.W., C. Sander and R.A. Nilan, 1975. The effect of sodium azide on cell processes in the embryonic barley shoot. Radiation Bot., 15: 315-322.
CrossRefDirect Link - Sinha, A. and J.P. Lal, 2007. Effect of mutagens on M1 parameters and qualitative changes induced in M2 generation in lentil. Legume Res. Int. J., 30: 180-185.
Direct Link - Suzuki, Y., Y. Sano, K. Ise, U. Matsukura, N. Aoki and H. Sato, 2008. A rice mutant with enhanced amylose content in endosperm without affecting amylopectin structure. Breed. Sci., 58: 209-215.
CrossRef - Van der Veen, J.H., 1966. Sterility, embryonic lethals and chlorophyll mutations. Arabidopsis Inform. Service, 33: 26-26.
Direct Link - Yuan, H.Y. and Z.L. Zhang, 1993. Effect of free radicals and temperature on sister chromatid exchange in Hordeum vulgare L. Acta Bot. Sin., 35: 20-26.
Direct Link