
Dwi Sutiningsih
Department of Epidemiology and Tropical Disease, Faculty of Public Health, University of Diponegoro, Semarang, Indonesia
LiveDNA: 62.16151
Mustofa
Department of Pharmacology, Faculty of Medicine, University of Gadjah Mada, Yogyakarta, Indonesia
Tri Baskoro Tunggul Satoto
Department of Parasitology, Faculty of Medicine, University of Gadjah Mada, Yogyakarta, Indonesia
Edhi Martono
Departement of Plant Pest and Diseases, Faculty of Agriculture, University of Gadjah Mada, Yoyakarta, Indonesia
Research Journal of Medicinal Plants
Year: 2017 | Volume: 11 | Issue: 2 | Page No.: 77-85







ABSTRACT
Background and Objectives: Aedes aegypti Linnaeus is a mosquito species that plays an important role as a vector of diseases in tropical and subtropical regions. Aedes aegypti (L.) resistance against temephos and pyrethroid has been widely reported in several countries including Indonesia. This study aimed to examine the neurotoxic mechanism of bruceine A biolarvicide and to determine the inhibitory activity of the enzyme acetylcholinesterase and gene Voltage-Gated Sodium Channel (VGSC) on A. aegypti (L.) larvae. Materials and Methods: Bruceine A was obtained by extraction and isolation of Makassar fruit’s seeds. Test of inhibition of acetylcholinesterase activity was based on Ellman method. Voltage-Gated Sodium Channel gene inhibitor was tested using PCR. The statistical analysis of inhibition of the enzyme acetylcholinesterase was performed by using Kruskal Wallis test followed by Mann Whitney. The data from observation on behavior response and VGSC gene were descriptively analyzed. Results: Bruceine A could inhibit the action of the enzyme acetylcholinesterase as much as 45.083±0.003%, with symptoms of a neurotoxic that is excitation, convulsions, tremors and paralysis. The result of fragment amplification of VGSC gene and electrophoresis on agarose gel 2% generated in a specific band with the size of the fragment bands of 336 bp on samples of larvae that were still sensitive to bruceine A and cypermethrin. Conclusion: Bruceine A isolated from the seeds of Brucea javanica (L.) Merr. has been demonstrated as biolarvicide against larvae of A. aegypti (L.), through its neurotoxic properties. The mechanism of neurotoxicity is mediated through inhibition of enzyme acetylcholinesterase and VGSC gene.
PDF Abstract XML References Citation
Received: January 03, 2017;
Accepted: March 10, 2017;
Published: March 15, 2017
How to cite this article
Dwi Sutiningsih, Mustofa, Tri Baskoro Tunggul Satoto and Edhi Martono, 2017. Neurotoxic Mechanism of Bruceine A Biolarvicide Against Aedes aegypti Linnaeus Larvae. Research Journal of Medicinal Plants, 11: 77-85.
URL: https://scialert.net/abstract/?doi=rjmp.2017.77.85
URL: https://scialert.net/abstract/?doi=rjmp.2017.77.85
INTRODUCTION
Aedes aegypti Linnaeus is a mosquito species that plays an important role as a vector of diseases in tropical and subtropical regions1,2. Aedes aegypti (L.) is the main vector of dengue fever, Dengue Hemorrhagic Fever (DHF) and Zika3,4. For extraordinary events (outbreaks) and DHF transmission season, the use of insecticides can not be avoided. Chemical control that is often done is by using temephos (Abate)5. According to Komariah et al.5, the continuous use of temephos for a long time and in high frequency causes a susceptibility decrease of target mosquitoes. There are also negative effects on non-target organisms such as natural enemies (predators) and it causes environmental pollution6. Aedes aegypti (L.) resistance against temephos has been widely reported in several countries including Indonesia7-9. It is necessary to keep alert for cross-resistance of A. aegypti (L.) against temephos and pyrethroid because in order to handle the increasing cases of DHF in the rainy season, it also carries out mosquito control efforts by fumigation using cypermethrin (pyrethroid)10. This condition requires the need for the development of vector control methods that are more environmental friendly and less costly, among others, is by the use of insecticides/larvicides deriving from plants.
Bruceine A is a quasinoid derived from Makassar fruit (Brucea javanica L. Merr.). Bruceine A quasinoid can be isolated from the seeds11,12, fruit13-15 and stem of B. javanica (L.) Merr.16. Bruceine A chemical structure is ([15]-3-methyl-2-bruseolid butanoil) with the molecular formula C26H34O11 and mass 522.54 g mol–1 in the form of amorphous powder with bitter taste13. Bruceine A has insecticidal activity, antifeedant and growth inhibition against tobacco budworm (Heliothis virescens F) and Spodoptera frugiperda17. As well as antifeedant activity against fourth instar larvae of Epilachna varivestis Mulsant and fifth instar larvae of Spodoptera eridania Crawler18. Bruceine A has larvicidal activity on the larvae of A. aegypti (L.) with a value of 50 and 90 lethal concentration (LC50 and LC90), respectively by 0.453 and 4.962 ppm. Therefore, it needs to be studied more in depth how is the mechanism of action of biolarvicide bruceine A against the larvae of A. aegypti (L.). This study aimed to examine the neurotoxic mechanism of biolarvicide bruceine A and to determine the inhibitory activity of the enzyme acetylcholinesterase and gene Voltage Gated Sodium Channel (VGSC) on larvae of A. aegypti (L.).
MATERIALS AND METHODS
Materials: The study was conducted from May-June, 2016. Bruceine A extraction and isolation was carried out in Pharmaceutical Biology laboratory, Faculty of Pharmacy, University of Gadjah Mada. The test of inhibitor activity of acetylcholinesterase enzyme and behavior response of A. aegypti (L.) larvae was carried out in Parasitology Laboratory, Faculty of Medicine, University of Gadjah Mada, while the test of VGSC gene inhibition with PCR was performed in Biochemistry Laboratory, Veterinary Faculty, University of Gadjah Mada.
Makassar fruit (B. javanica L. Merr.) was bought from a wholesaler of medicinal plants (Aneka Herbal Yogyakarta), being identified in the Laboratory of Pharmaceutical Biology, Faculty of Pharmacy, University of Gadjah Mada to obtain more information on the properties of the simplicial. Other substances and solvents used in extraction and isolation of bruceine A are hexane (commercial, E. Merck), methanol (commercial, E. Merck), dichloromethane (commercial, E. Merck), chloroform (commercial, E. Merck), ethyl acetate (commercial, E. Merck) and silica gel (E. Merck). The instar III larvae of A. aegypti (L.) to the beginning of instar IV were acquired and developed at the Laboratory of Parasitology, Faculty of Medicine, University of Gadjah Mada.
Bruceine A isolation from the seed of Brucea javanica (L.) Merr.: Bruceine A isolation from B. javanica (L.) Merr. seed based on method of Mangungsong11. Brucea javanica (L.) Merr., dried seeds (5 kg) was made it into powder and being shaken with a solution of hexane (15 L), then filtered and separated from the hexane solution (commercial). Subsequently extracted with methanol 15 L (commercial). The extraction of the methanol was through evaporation to obtain a thick extract and then distilled water with the same amount was added to form a suspension. The suspension was partitioned with hexane (3 L) until we obtained the hexane fraction and methanol-water. Hexane fraction was separated and stored while the methanol-water solution was extracted with dichloromethane 1 L (commercial) repeatedly (3 times) to obtain the organic layer of dichloromethane. The result in the organic layer was taken and evaporated to obtain a concentrate and then it was made solution in methanol (100-250 mL) at a temperature of 60̊C and then stored at room temperature. The methanol solution was allowed to stand at room temperature to form crystalline compounds that expected as bruceine A. Further separation was by filtration. The rest of the filtrate/residue was separated by thin layer of chromatography. The filtrate/residue was evaporated and further was separated if necessary.
Behavioral response test: Behavioral response was measured using the method described by Sharma et al.19 with a little modification. The third instar larvae of A. aegypti (L.) to the beginning of fourth instar were put in a plastic cup filled with bruceine A with various concentrations, each of 25 larvae, which consisted of three replications with positive control is temephos 1% while the negative control is used only as much as 100 mL of distilled water. Behavioral response of A. aegypti (L.) larvae was monitored every 10 and 30 min, 1, 5 and 24 h after treatment. Data on behavioral response was observed by physical response, posture and behavior. Data was recorded and photographed using Nikon D5300 camera. Similar observations were performed in the control group and then compared to the treatment group.
Inhibition of the enzyme acetylcholinesterase activity test: Inhibition of the enzyme acetylcholinesterase activity test was based on the method of Ellman et al.20 with slight modifications. Into a test tube containing 1.95 mL of 0.1 M potassium phosphate buffer pH 7.5, was added 200 mL homogenate larvae of A. aegypti (L.) 150 mL DTNB 0.0011 M in phosphate buffer and 100 mL bruceine A solution and alkylaryl polyglycol ether emulator of 400 mg L–1 0.1%. The mixture was given buffer solution as control. After it was perfectly whipped and left for 10 min, then into each test tube it was added 100 mL acetylcholine iodide 0.0105 M in phosphate buffer. The test was performed with six concentrations of bruceine A. The blank mixture that contained the same component with the test mixture, except for the homogenates, enzyme source was replaced with a phosphate buffer. The reaction was left for 30 min, then light absorption solution in each tube was measured at a wavelength of 405 nm using a spectrophotometer (Shimadzu UV-1800, Japan and Europe).
Test of inhibitor of gene VGSC: The VGSC inhibitor test was done using PCR method. This test consists of six concentrations of bruceine A with positive control using cypermethrin and the negative control using 100 mL of distilled water. The genomic DNA of the mosquito larvae was extracted using protocol SYNC DNA kit (Geneaid). A total of 50 mg of larvae tissue to a mortar and were added liquid nitrogen and grind the tissue thoroughly using a pestle. Refill the mortar occasionally with liquid nitrogen to keep the sample frozen. The tissue powder were transferred to 1.5 mL microcentrifuge tube and added 200 μL of GST buffer and 20 μL of proteinase K then vortexed thoroughly. The mixture was incubated for 1-3 h at 65̊C or until the sample lysate becomes clear. During incubation, invert the tube occasionally and transfer the required volume of elution buffer (200 μL per sample) to a new 1.5 mL microcentrifuge tube and heat to 60̊C. If insoluble material remains following incubation, then being centrifuged for 2 min at 14-16,000 rpm and then carefully transferred the supernatant to a new 1.5 mL microcentrifuge tube. It was added 200 μL of GST buffer and shaken vigorously for 10 sec. Meanwhile, add 200 μL of absolute ethanol to the sample lysate and mix immediately by shaking vigorously for 10 sec. If precipitate appears, break it up as much as possible with a pipette. The all of the mixture were transferred to the GS column and being centrifuged at 14-16,000 rpm for 1 min and then transferred the GS column to a new 2 mL collection tube. It was added 400 μL of W1 buffer to the GS column and centrifuged at 14-16,000 rpm for 30 sec then was discarded the flow-through. The GS column was added 600 μL of wash buffer and being centrifuged at 14-16,000 rpm for 30 sec and then was discarded the flow-through. The GC column was placed back in 2 mL collection tube and then being centrifuged again for 3 min at 14-16,000 rpm to dry the column matrix. The dried GS column was transferred to a clean 1.5 mL microcentrifuge tube and was added 100 μL of pre-heated elution buffer, TE buffer or water into the center of the column matrix. Let stand for at least 3 min to allow elution buffer, TE buffer or water to be completely was absorbed. It was centrifuged again at 14-16,000 rpm for 30 sec to elute purified DNA. The VGSC gene fragment was amplified by PCR using primers: DSAAF : 5`’ CGGGAGGTAAGTTATTGTGA 3`’ dan DSAAR : 5`’ ATGGAAATGCCTTACCCTA 3`’ (KM. 677279). Visualization of PCR results was done by using method of electrophoresis on agarose gel 2% and evaluated under UV light 300 nm.
Data analysis: The percentage of inhibition of the enzyme acetylcholinesterase was calculated using Ellman formula20 and expressed as Mean±Standard Error of Mean (SEM), while the percentage of sink, almost die and dead larvae were expressed as Mean±Standard Deviation (SD). The statistical analysis was performed by using Kruskal Wallis test followed by Mann Whitney. Statistically significant differences were indicated at p<0.05. The data from the result of amplification of VGSC gene were analyzed descriptively21.
RESULTS
Behavioral responses of A. aegypti (L.) larvae: Observation of behavioral responses of larvae A. aegypti (L.) after treatment with bruceine A was done directly by visual observation at changes in the physical response (movement/behavior) of larvae A. aegypti (L.) compared with controls19,22.
 | |
| Fig. 1(a-d): | Digital photomicrographs of early IV instar larvae of A. aegypti (L.) (a) Control larvae, (b) Larvae treated with 1 ppm temephos, (c) Larvae treated with 4 ppm bruceine A and (d) Larvae treated with 16 ppm bruceine A |
| The circled larvae show the aggressive anal gills biting behaviour forming ring-shaped structure | |
| Table 1: | Behavioral response of A. aegypti (L.) larvae after treatment with bruceine A, temephos (1 ppm) and control over the time period of observation |
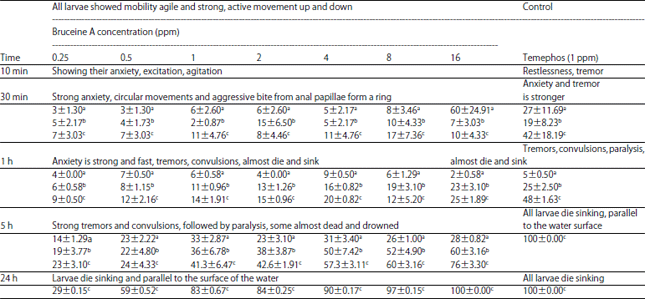 | |
| a: Larvae sink±SD, b: Larvae almost die/moribund±SD, c: Dead larvae±SD, SD: Standard deviation, ppm: Part per million | |
Observation of behavioral responses of larvae of A. aegypti (L.) after treatment with bruceine A showed different behavior patterns compared to controls (Table 1, Fig. 1). Immediately after being exposed to various concentrations of bruceine A, all the larvae of A. aegypti (L.) still showed a natural vigorous movement.
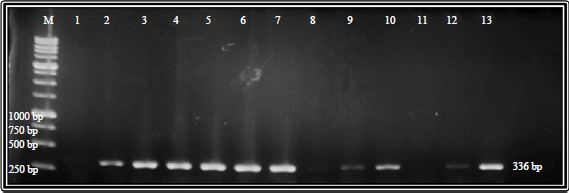 | |
| Fig. 2: | Fragment bands of 336 bp on samples of A. aegypti (L.) larvae after treatment with bruceine A, cypermethrin and control |
M: Marker, Lane 1: Control (with out treatment), Lane 2: 0.5 μL Cypermethrin, Lane 3: 1 μL Cypermethrin, Lane 4: 2 μL Cypermethrin, Lane 5: 4 μL Cypermethrin, Lane 6: 8 μL Cypermethrin, Lane 7: 16 μL Cypermethrin, Lane 8: 0.5 ppm Bruceine A, Lane 9: 1 ppm Bruceine A, Lane 10: 2 ppm Bruceine A, Lane 11: 4 ppm Bruceine A, Lane 12: 8 ppm Bruceine A and Lane 13: 16 ppm Bruceine A | |
| Table 2: | Inhibition of acetylcholinesterase enzyme on the larvae of A. aegypti (L.) after treatment with bruceine A at various concentrations, temephos (1 ppm), malathion (1 ppm) and control (without treatment) |
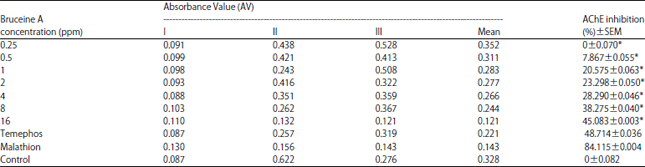 | |
| *p<0.05 when compared with negative control, Kruskal Wallis, Mann Whitney, AChE: Acetylcholinesterase enzyme, ppm: Part per million, SEM: Standard error of mean | |
However, after 10 min of exposure with bruceine A, larvae of A. aegypti (L.) indicated anxiety, excitation and agitation (Table 1). Most larvae showed heavy anxiety response and very aggressive behavior as expressed by biting their anal gills using their mouth so that it looked like a circular motion to form a ring structure (Fig. 1b-d). After 1 h, most of the larvae that were exposed to the highest concentration of bruceine A (16 ppm) and temephos (1 ppm), showed tremors movement and convulsions followed by paralysis and eventually death. On the other hand, the larvae of A. aegypti (L.) treated with bruceine A low concentration, they appeared more aggressive. Five hours after exposure bruceine A at various concentrations, A. aegypti (L.) larval mortality was recorded as much as 23-76%, whereas in the treatment with 1 ppm temephos, the mortality of larvae reached 100%. Larval mortality of 100% was reached on treatment with bruceine A at concentration of 16 ppm after 24 h of observation (Table 1).
Activity of the enzyme acetylcholinesterase inhibition: The results of measurement of the enzyme acetylcholinesterase inhibitory activity on the larvae of A. aegypti (L.) after treatment with bruceine A at various concentrations, temephos (1 ppm), malathion (1 ppm) and control are shown in Table 2. At the highest concentration of bruceine A (16 ppm), the activity of the enzyme acetylcholinesterase inhibition was 45.083±0.003%, whereas with 1 ppm temephos and 1 ppm malathion the inhibition was 48.714±0.036 and 84.115±0.004%, respectively. Kruskal Wallis analysis followed Mann Whitney indicated that there were significant differences in inhibitory activity of the enzyme acetylcholinesterase of larvae A. aegypti (L.) at various concentrations of bruceine A (p<0.05).
Inhibition of Voltage-Gated Sodium Channels (VGSC) gene: The results of amplification of VGSC gene was the fragment band with the size of 336 bp on samples of larvae that were still sensitive to bruceine A and cypermethrin. The PCR results of VGSC gene on the larvae of A. aegypti (L.) is presented in Fig. 2. It shows that only samples of A. aegypti (L.) larvae with treatment of bruceine A at concentration of 0.5 ppm (sample No. 8) and 4 ppm (sample No. 11) there were no specific visible band.
DISCUSSION
Behavioral responses of A. aegypti (L.) larvae: The first step in the measurements of toxic effects was the observation of the physical response/behavior of A. aegypti (L.) larvae after treatment with the test materials. The behavioral response of A. aegypti (L.) larvae at instar III and early instar IV after the treatment with bruceine A at various concentrations showed a different pattern of behavior compared to the control group (no treatment) and the treatment group with temephos 1 ppm (Table 1, Fig. 1). It was as expected from the effects of toxic compounds of bruceine A on the behavior of A. aegypti (L.) larvae. Ten minutes after exposure of bruceine A, larvae of A. aegypti (L.) indicated anxiety, excitation and agitation that were signs of being poisoned in larvae of A. aegypti (L.). Sanjaya and Safaria23 stated that the toxic compounds that enter the body of larvae can cause four-stage responses, including excitation, convulsions, paralysis and death. The high movement of the larvae after contact with toxic substance of bruceine A which was characterized by symptoms of anxiety (excitation). The rapid movement of larvae was the larval form of the stress which indicates that the toxic material of bruceine A has entered into the body of larvae. Further stages characterized by the movements of larvae that were slower due to the increasing number of toxic compounds that enter the body of larvae during the time of observation. The slow movement of the larvae when larvae were given touch will show that the larval stage has experienced paralysis and eventually will die. Some of the larvae showed heavy anxiety response and very aggressive with biting their anal gills using their mouth so that it looks like a circular motion to a form ring like shape structure (Fig. 1c, d). The results are consistent with study of Sutiningsih and Nurjazuli24, who proved that early instar IV of A. aegypti (L.) larvae shows neurotoxic symptoms after treatment with brusatol. Brusatol biolarvicide is proven significantly to have larvicidal effect on A. aegypti (L.) with the LC50 of 0.669 ppm and LC90 that is at 8.331 ppm. Sharma et al.20 already proved that the extract of leaves and stems of Achyranthes aspera on the larvae of A. aegypti (L.) at early fourth instar causes behavioral agitation, excitation and the aggressive movement of biting anal gills. These symptoms were similar to symptoms caused by a neurotoxin. So it can be concluded that extracts of A. aspera can act as cytolysin that affect larval neuromuscular system. Circular movement of the larvae, accompanied by aggressive biting behavior of anal gills prove their neurotoxic symptoms of bruceine A on the larvae of A. aegypti (L.). Anal gills on mosquitoes have a role in regulating electrolyte balance needed for survival25. Bruceine A causes electrolyte imbalances in the anal area, causing the larvae bite anal gills with great force. The results of this research were similar with the results of previous studies indicating that bioactive compounds of plants can act as a neurotoxin despite aggressive move and uncoordinated along with other toxic symptoms observed at interval time that was relatively different26-29. Similar behavioral observation has also been reported by Warikoo and Kumar30 on the larvae of A. aegypti (L.) after treatment with root extract of Argemone mexicana. The results of microscopic observation on larvae of A. aegypti (L.) after treatment with the extracts of A. mexicana at concentration of 91.331 and 156.684 ppm for 24 h showed a structural deformity of the anal gills30. This damage causes the malfunctioning of osmotic and ionic regulation that was suspected as the cause of larval death of A. aegypti (L.)28.
Inhibition of the activity of the enzyme acetylcholinesterase: Neurotoxic symptoms in the larvae of A. aegypti (L.) after treatment with bruceine A was also proven by the inhibition of the activity of the enzyme acetylcholinesterase. Bruceine A as a toxic substance can inhibit the action of the enzyme acetylcholinesterase, which function is to hydrolyze the acetylcholine. At the highest concentration of bruceine A that was 16 ppm, the inhibitory activity of the enzyme acetylcholinesterase was 45.083±0.003%, while with temephos 1 ppm and malathion 1 ppm, the inhibitory activity of the enzyme acetylcholinesterase were higher than bruceine A, ie. amounted to 48.714±0.036 and 84.115±0.004%, respectively (Table 2). Inhibitory activity of the enzyme acetylcholinesterase increased if the concentration of bruceine A was higher and was statistically significant (p<0.05). The toxic compounds of bruceine A enter the larval body through the pores of the skin/cuticle, digestive tract and siphon. The toxic compounds cause the disruption of the digestive system, the respiratory system and the nervous system in the larvae31. Under normal circumstances the function of acetylcholine is to conduct nerve impulses. Acetylcholine undergoes hydrolysis by the enzyme acetylcholinesterase into choline and acetic acid32. The impact of enzyme acetylcholinesterase inhibition would cause disruption and damage of the nervous system causing the muscles remain contracted, finally exhausted and paralyzed leading to death33. The larval death is caused by a paralysis on the respiratory muscles so the larvae can not breathe and eventually die.
The inhibitory activity of acetylcholinesterase on the larvae of A. aegypti (L.) with 1 ppm malathion treatment was higher (84.115±0.004%) as compared to treatment with 1 ppm temephos (48.714±0.036%). It is likely because the sensitivity of each stadium/instar of the larvae towards the insecticides is different34. In addition, the toxicity of temephos against insects is closely related to how much the molecule is oxidized, the more it is oxidized, the more it loses its toxicity35. Temephos is easily degraded when exposed to sunlight, so the ability to kill the larvae depends on the degradation36. Temephos and malathion are both insecticides of the organophosphate class which inhibit acetylcholinesterase, an enzyme located in the central and peripheral nervous systems. Organophosphate disables acetylcholinesterase by phosphorylating the serine hydroxyl group that is located on the active side of acetylcholinesterase, which will form the phosphorylated cholinesterase compounds. Cholinesterase enzyme can not function any more, because of the reduction of the levels of the active sites of these enzyme37. Reduced cholinesterase enzyme resulted in a decreased ability to hydrolyze the acetylcholine, resulting in longer acetylcholine receptor activation, which will intensify and prolong the effects of cholinergic nerve stimulation before and after ganglion (pre and postganglionic). The decrease in plasma cholinesterase activity will return to normal within 3 weeks while red blood cells will take 2 weeks. Inhibition of the enzyme acetylcholinesterase by organophosphate pesticides can lead to the accumulation of both acetylcholine muscarinic and nicotinic receptors in the central and peripheral nervous system. The excess of acetylcholine originally cause excitation, paralysis and death33,38.
Inhibitory of Voltage Gated Sodium Channel(VGSC) gene: The analysis results of the fragments from Voltage-Gated Sodium Channel (VGSC) genes of A. aegypti (L.) larvae showed a difference between bruceine A and control treatment (Fig. 2). The samples of larvae bruceine A at concentration of 2, 8 and 16 ppm produced a specific band with the size of the fragment bands of 336 (bp), as well as with cypermethrin. This indicates that the mechanism of action of bruceine A in killing larvae is presumably through the inhibitor of VGSC gene, like the cypermethrin insecticides. In contrast, the control group (no treatment) showed no specific band similar to those in cypermethrin. At the highest concentration bruceine A of 16 ppm, the fragment band of VGSC gene was thicker than the lower concentration. This is most likely because of the large volume of different concentrations of DNA during DNA isolation, the solubility of the compound is not complete and contamination during DNA extraction or PCR running. Molecular increase in VGSC gene expression caused by an increase in amplification of genes that encoding VGSC39,40.
Voltage Gated Sodium Channel (VGSC) is a transmembrane protein found in nerve cells and muscle cells that plays a role in action potential of cell. The VGSC is part of a proteinsuperfamily41. The constituent sub unit of voltage-gated sodium channel is a polypeptide chain consisting of more than 1800 amino acids. The VGSC consists of four homologous domains(I-IV), each consisting of one to six hydrophobic segments (S1-S6) and a P-loop between S5 and S640. The principle of VGSC is to continue the action potential to become signaling on other nerve cells. The action potential starts when nerve fibers are stimulated, the Voltage Gated Sodium Channel (VGSC) will open and sodium ions that are positively charged moves into the cell to change the resting potential (polarization) into action potential (depolarization) forming a membrane potential, a few milliseconds later the channel closes and the signal transmitted to the next nerve cells42,43. As a contact poison, bruceine A that applied directly can penetrate the skin/cuticle or sensory glands and other organs related to the cuticle. Bruceine A is suspected to be dissolved to the fat layer of cuticle and enter the body of the insect. As a result, the channel which should closed will remain open, so that many sodium will be in the cells causing the discharge of potential action that occurs continuously in the nervous system of the insect causing hyperexcitability and seizures insects and eventually die44.
Cypermethrin is a class of synthetic pyrethroid, which is characterized as axonic /neurotoxin. Cypermethrin would be bound to a nerve protein known as VGSC. Cypermethrin tied to this gate and prevent normal closure which resulted in sustained nerve stimulation. This causes tremors and in coordination of movements in insect that is poisoned45. The use of cypermethrin is highly popular because of its effectiveness and relatively inexpensive. Chemical structure of cypermethrin resembles pyrethrum, with high and stable biological toxicity compared to other natural toxins46. The applications of insecticide cypermethrin pyrethroid on insect populations which is sensitive can bind Voltage Gated Sodium Channel (VGSC) protein, which regulates the nerve impulses. In insects that are resistant to insecticides from pyrethroid group and DDT, the important resistance mechanism is the presence of a change or mutation in the VGSC gene.
CONCLUSION
Bruceine A isolated from the seeds of Makassar fruit (B. javanica L. Merr.) can act as a neurotoxin in the larvae of A. aegypti (L.). The mechanism of action of bruceine A biolarvicide is through inhibitor of the enzyme acetylcholinesterase and VGSC gene, which showed by their behavioral responses that impact the larval neuromuscular systems that are hyperexcitation, convulsions, paralysis and aggressive biting movement from anal gills which forms a ring structure.
SIGNIFICANCE STATEMENTS
The result of this study reinforces the potential of bruceine A as a natural larvicide in the control of disease vectors, especially A. aegypti (L.). This study will help the researchers to discover the critical areas of target site of biolarvicide bruceine A, which has not been explored yet. Therefore, a new theory about the mechanism of neurotoxic of biolarvicide bruceine A and possibly other actions, can be achieved.
ACKNOWLEDGMENTS
The authors would like to thank the Aris Haryanto, as head of the Department of Biochemistry, Faculty of Veterinary Medicine, University of Gadjah Mada, Indonesia and Suhendra Pakpahan Department of Biochemistry, Faculty of Veterinary Medicine, University of Gadjah Mada, Indonesia who have helped in the result analysis of PCR.
REFERENCES
- Morrison, A.C., E. Zielinski-Gutierrez, T.W. Scott and R. Rosenberg, 2008. Defining challenges and proposing solutions for control of the virus vector Aedes aegypti. PLoS Med., Vol. 5.
CrossRefDirect Link - Ponlawat, A., J.G. Scott and L.C. Harrington, 2005. Insecticide susceptibility of Aedes aegypti and Aedes albopictus across Thailand. J. Med. Entomol., 42: 821-825.
PubMedDirect Link - Petersen, L.R., D.J. Jamieson, A.M. Powers and M.A. Honein, 2016. Zika virus. N. Engl. J. Med., 374: 1552-1563.
CrossRefDirect Link - Kroeger, A., A. Lenhart, M. Ochoa, E. Villegas, M. Levy, N. Alexander and P.J. McCall, 2006. Effective control of dengue vectors with curtains and water container covers treated with insecticide in Mexico and Venezuela: Cluster randomised trials. Br. Med. J., 332: 1247-1252.
CrossRefDirect Link - Boewono, D.T. and Widiarti, 2007. Susceptibility of dengue haemorrhagic fever vector (Aedes aegypti) against organophosphate insecticides (malathion and temephos) in some districts of Yogyakarta and Central Java Provinces. Bull. Health Res., 35: 49-56.
Direct Link - Biber, P.A., J.R. Duenas, F.L. Almeida, C.N. Gardenal and W.R. Almiron, 2006. Laboratory evaluation of susceptibility of natural subpopulations of Aedes aegypti larvae to temephos. J. Am. Mosq. Control Assoc., 22: 408-411.
CrossRefDirect Link - Rodriguez, M.M., J. Bisset, M. Ruiz and A. Soca, 2002. Cross-resistance to pyrethroid and organophosphorus insecticides induced by selection with temephos in Aedes aegypti (Diptera: Culicidae) from Cuba. J. Med. Entomol., 39: 882-888.
CrossRefDirect Link - NoorShahida, A., T.W. Wong and C.Y. Choo, 2009. Hypoglycemic effect of quassinoids from Brucea javanica (L.) Merr (Simaroubaceae) seeds. J. Ethnopharmacol., 124: 586-591.
CrossRefDirect Link - Subeki, H. Matsuura, K. Takahashi, K. Nabeta, M. Yamasaki, Y. Maede and K. Katakura, 2007. Screening of Indonesian medicinal plant extracts for antibabesial activity and isolation of new quassinoids from Brucea javanica. J. Nat. Prod., 70: 1654-1657.
CrossRefDirect Link - Bawm, S., H. Matsuura, A. Elkhateeb, K. Nabeta and Subeki et al., 2008. In vitro antitrypanosomal activities of quassinoid compounds from the fruits of a medicinal plant, Brucea javanica. Vet. Parasitol., 158: 288-294.
CrossRefPubMedDirect Link - Liu, J.Q., C.F. Wang, X.Y. Li, J.C. Chen, Y. Li and M.H. Qiu, 2011. One new pregnane glycoside from the seeds of cultivated Brucea javanica. Arch. Pharm. Res., 34: 1297-1300.
CrossRefDirect Link - Liu, Z.L., S.H. Goh and S.H. Ho, 2007. Screening of Chinese medicinal herbs for bioactivity against Sitophilus zeamais Motschulsky and Tribolium castaneum (Herbst). J. Stored Prod. Res., 43: 290-296.
CrossRefDirect Link - Klocke, J.A., M. Arisawa, S.S. Handa, A.D. Kinghorn, G.A. Cordell and N.R. Farnsworth, 1985. Growth inhibitory, insecticidal and antifeedant effects of some antileukemic and cytotoxic quassinoids on two species of agricultural pests. Cell. Mol. Life Sci., 41: 379-382.
CrossRefDirect Link - Leskinen, V., J. Polonsky and S. Bhatnagar, 1984. Antifeedant activity of quassinoids. J. Chem. Ecol., 10: 1497-1507.
CrossRefDirect Link - Sharma, A., S. Kumar and P. Tripathi, 2015. Impact of Achyranthes aspera leaf and stem extracts on the survival, morphology and behaviour of an Indian strain of dengue vector, Aedes aegypti L. (Diptera: Culicidae). J. Mosquito Res., Vol. 5.
CrossRef - Ellman, G.L., K.D. Courtney, V. Andres Jr. and R.M. Featherstone, 1961. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol., 7: 88-90.
CrossRefDirect Link - Sanjaya, Y. and T. Safaria, 2006. [Toxicity of poison from spider Nephila sp. to Aedes aegypti L. larvae]. J. Biodiversitas, 7: 191-194.
Direct Link - Sutiningsih, D. and Nurjazuli, 2017. Effect of brusatol biolarvicide administration on behavioral response of Aedes aegypti and its toxicity on vero cells. J. Biol. Sci., 17: 127-135.
CrossRefDirect Link - Dharmagadda, V.S.S., S.N. Naik, P.K. Mittal and P. Vasudevan, 2005. Larvicidal activity of Tagetes patula essential oil against three mosquito species. Bioresour. Technol., 96: 1235-1240.
CrossRefDirect Link - Chaithong, U., W. Choochote, K. Kamsuk, A. Jitpakdi and P. Tippawangkosol et al., 2006. Larvicidal effect of pepper plants on Aedes aegypti (L.) (Diptera: Culicidae). J. Vector Ecol., 31: 138-144.
CrossRefDirect Link - Kumar, S., R. Warikoo and N. Wahab, 2010. Larvicidal potential of ethanolic extracts of dried fruits of three species of peppercorns against different instars of an indian strain of dengue fever mosquito, Aedes aegypti L. (Diptera: Culicidae). Parasitol. Res., 107: 901-907.
CrossRefPubMedDirect Link - Warikoo, R. and S. Kumar, 2013. Impact of Argemone mexicana extracts on the cidal, morphological and behavioral response of dengue vector, Aedes aegypti L.(Diptera: Culicidae). Parasitol. Res., 112: 3477-3484.
Direct Link - Boublik, Y., P. Saint-Aguet, A. Lougarre, M. Arnaud, F. Villatte, S. Estrada-Mondaca and D. Fournier, 2002. Acetylcholinesterase engineering for detection of insecticide residues. Protein Eng. Des. Sel., 15: 43-50.
CrossRefDirect Link - Kim, I.H., R. Suzuki, Y. Hitotsuyanagi and K. Takeya, 2003. Three novel quassinoids, javanicolides A and B and javanicoside A, from seeds of Brucea javanica. Tetrahedron, 59: 9985-9989.
CrossRefDirect Link - Bisset, J.A., M.M. Rodriguez, Y. Ricardo, H. Ranson, O. Perez, M. Moya and A. Vazquez, 2011. Temephos resistance and esterase activity in the mosquito Aedes aegypti in Havana, Cuba increased dramatically between 2006 and 2008. Med. Vet. Entomol., 25: 233-239.
CrossRefDirect Link - Weill, M., P. Fort, A. Berthomieu, M.P. Dubois, N. Pasteur and M. Raymond, 2002. A novel acetylcholinesterase gene in mosquitoes codes for the insecticide target and is non-homologous to the ace gene Drosophila. Proc. Biol. Sci., 269: 2007-2016.
CrossRefPubMedDirect Link - Cloyd, R., 2004. Natural indeed: Are natural insecticides safer and better than conventional insecticides? Illinois Pesticide Rev., 17: 1-7.
Direct Link - Brengues, C., N.J. Hawkes, F. Chandre, L. McCarroll and S. Duchon et al., 2003. Pyrethroid and DDT cross-resistance in Aedes aegypti is correlated with novel mutations in the voltage-gated sodium channel gene. Med. Vet. Entomol., 17: 87-94.
CrossRefPubMedDirect Link - Kushwah, R.B.S., C.L. Dykes, N. Kapoor, T. Adak and O.P. Singh, 2015. Pyrethroid-resistance and presence of two knockdown resistance (kdr) mutations, F1534C and a novel mutation T1520I, in Indian Aedes aegypti. PLoS Negl. Trop. Dis, Vol. 9.
CrossRefDirect Link - Catterall, W.A., 2000. From ionic currents to molecular mechanisms: The structure and function of voltage-gated sodium channels. Neuron, 26: 13-25.
CrossRefDirect Link - Zhou, W., I. Chung, Z. Liu and A.L. Goldin, 2004. A voltage-gated calcium-selective channel encoded by a sodium channel-like gene. Neuron, 42: 101-112.
CrossRefDirect Link - Dong, K., 2007. Insect sodium channels and insecticide resistance. Invert. Neurosci., 7: 17-30.
CrossRefDirect Link - Krieger, R.I., 2010. Haye's Handbook of Pesticide Toxicology. 3rd Edn., Academic Press, Cambridge, Massachusetts, ISBN: 978-0-12-374367-1, Pages: 2342.
Direct Link