
Judith A. Makwali
Department of Biological Sciences, School of Science, University of Eldoret, P.O. Box 1125-30100, Eldoret, Kenya
Frederick M.E. Wanjala
Department of Biological Sciences, School of Science, University of Eldoret, P.O. Box 1125-30100, Eldoret, Kenya
Johnstone Ingonga
Center for Biotechnology Research and Development (CBRD), Kenya Medical Research Institute (KEMRI), P.O. Box 54840-00200, Nairobi, Kenya
Christopher O. Anjili
Center for Biotechnology Research and Development (CBRD), Kenya Medical Research Institute (KEMRI), P.O. Box 54840-00200, Nairobi, Kenya
Research Journal of Medicinal Plants
Year: 2015 | Volume: 9 | Issue: 3 | Page No.: 90-104







ABSTRACT
Natural products and herbicides are potential sources of new and selective agents for the treatment of important tropical diseases caused by parasites. Leishmaniasis is a major health problem worldwide and affects millions of people especially in developing nations. There is no immunoprophylaxis (vaccination) available for Leishmania infections and conventional treatments are unsatisfactory. Therefore, antileishmanial drugs are urgently needed. In this study, the antileishmanial activity of two herbicides (acriflavine and trifluralin) and two plant biocides (triterpenoid saponin and plumbagin) extracts against Leishmania major parasites was evaluated in vitro. Different doses of acriflavine, trifluralin, saponin and plumbagin extracts were tested against L. major promastigotes and amastigotes. Pentostam and pentamidine were used as the reference drugs under the same experimental conditions. Anti-parasitic effect was observed for saponin and acriflavine for amastigote and promastigote suppression with a 50% inhibitory concentration (IC50) values of 80 and 20 μg mL–1, respectively compared to the standard drugs, pentostam, which had an IC50 value of 130 μg mL–1 for amastigotes and pentamidine which had an IC50 value of 24 μg mL–1 for promastigotes. The optimal efficacy of the compounds tested was 100%, albeit the concentrations at optimal efficacy were significantly different (p<0.05) among the test compounds. Significant differences were also discerned in 90% cytotoxic concentration (CC90) and 50% Cytotoxic Concentration (CC50) after 24 h exposure (p<0.05). Highest CC90 was recorded in acriflavine while saponin had the lowest CC90. The in vitro activity of the test compounds (trifluralin, plumbagin, acriflavine and saponin) against intracellular L. major amastigotes (IC50 = 150, 400, 200 and 80 μg mL–1, respectively) in the absence of obvious cytotoxicity on murine macrophage cells (CC50 >3,400 μg mL–1) demonstrates the potential of these biocides and herbicides in the treatment of cutaneous leishmaniasis.
PDF Abstract XML References Citation
Received: May 12, 2015;
Accepted: June 16, 2015;
Published: June 26, 2015
How to cite this article
Judith A. Makwali, Frederick M.E. Wanjala, Johnstone Ingonga and Christopher O. Anjili, 2015. In vitro Studies on the Antileishmanial Activity of Herbicides and Plant Extracts Against Leishmania major Parasites. Research Journal of Medicinal Plants, 9: 90-104.
URL: https://scialert.net/abstract/?doi=rjmp.2015.90.104
URL: https://scialert.net/abstract/?doi=rjmp.2015.90.104
INTRODUCTION
Leishmaniasis is vector-borne disease caused by a protozoan endo-parasite of the Leishmania genus and is transmitted by the bite of phlebotomine sandflies (Adebayo et al., 2013). Leishmaniasis has been reported to be one of the world’s most neglected diseases with an estimated 12 million people and an annual incidence of two million new cases (about 75% cases of cutaneous leishmaniasis and 25% cases of visceral leishmaniasis) (Gomes et al., 2012). The World Health Organization (WHO) also reports that, about 350 million people are considered at risk of contracting leishmaniasis especially people living in the developing countries (WHO., 2010). Cutaneous Leishmaniasis (CL) is the most common form of leishmaniasis and has an annual incidence of 1-1.5 million cases (90% of these are found in the Old World (Garnier et al., 2007). In CL, the disease is normally localized to the site of infection within dermal macrophages. Typically, papules develop at the site of infection; enlarge to a nodule and progress to ulcerated lesions, which last less than a year (Murray et al., 2005). Treatment for CL aims to accelerate healing, minimize scarring and prevent the development of more complex manifestations. The leishmaniasis present major public health problems throughout tropical and subtropical countries, with a broad spectrum of clinical manifestations, ranging from self-healing in cutaneous forms to fatal disease in the visceral infections. In the New World, cutaneous leishmaniasis is a zoonosis that involves different species of Leishmania and a great variety of sylvatic vectors and mammal reservoirs.
The therapeutic arsenal routinely employed to treat patients with leishmaniasis is limited and unsatisfactory. For cutaneous leishmaniasis, pentavalent antimonials (sodium stibogluconate or meglumine antimoniate) at 20 mg kg-1 day-1 for 20-28 consecutive days are the first line therapeutic scheme recommended by WHO. These compounds are highly toxic, poorly tolerated and their effectiveness highly variable (Zauli-Nascimento et al., 2010). In addition, leishmaniasis are developing resistance to the present drugs of choice which are expensive and toxic; therefore, there is an urgent need for alternative chemotherapeutic regimens (Oryan, 2015). The known activities of trifluralin and acriflavine and considering that they both target the kinetoplastid parasites makes them appropriate molecules for investigation, both in in vitro and in in vivo systems. One of the rational approaches to Leishmania chemotherapy is to test compounds that are known to have antiparasitic effects within the kinetoplastid parasites.
In traditional medicine, many plants have already provided valuable clues for potentially antiparasitic compounds, especially simple quinones, quassinoids and related naphthoquinones (Gonzalez-Coloma et al., 2012). One of the main reasons for which a number of plant metabolites with leishmanicidal activity have not made it to clinical evaluation is their high cytotoxicity. This lack of selectivity is evident in products, such as the phorbol esters, the quassinoids and the acetogenins. On the other hand, there are metabolites, such as gabunine and ursolic acid that lack cytotoxicity, but have only a weak in vivo leishmanicidal activity. Perhaps chemical transformation of the latter metabolites could improve their antiprotozoal activity, while, at the same time, maintaining their lack of cytotoxicity. In the results reported here, plumbagin extract used showed leishmanicidal activity and lacked any obvious cytotoxicity. Existing information confirms that plumbagin has many antimicrobial, dermatological and cytological activities against protozoan parasites. Saponins vary widely in the kind and intensity of biological activity. Some of the more important reported activities include haemolytic, local irritant, inflammatory, cytotoxic and antimicrobial activity.
The objective of the present study was to determine the antileishmanial activity of trifluralin, acriflavine, saponin and plumbagin extract against L. major parasites.
MATERIALS AND METHODS
Compounds: Plant extract solutions and the reference drugs were prepared in 100% Dimethyl Sulfoxide (DMSO) at 20 mg mL-1. Pentamidine isethionate, sodium stibogluconate (Pentostam), acriflavine, trifluralin and saponin extract were purchased from SIGMA. Roots of Plumbago capensis were collected from Karura forest, Nairobi, Kenya. Plant extractions were done in KEMRI at center for Traditional Medicine and Drugs Research. The purity of plumbagin extract was determined by High-Performance Liquid Chromatography (HPLC; purity/90%) and Nuclear Magnetic Resonance (NMR) spectroscopy. In most experiments, pentostam was used as a reference drug.
Parasites: Metacyclic promastigotes of L. major strain IDUB/KE/83 = NLB-144, which was originally isolated in 1983 from a female P. duboscqi collected near Marigat, Baringo District, Rift Valley Province, Kenya, was used (Beach et al., 1984). This strain has been maintained by cryopreservation and in vitro culture and periodic passage in BALB/c mice at Kenya Medical Research Institute (KEMRI).
Mouse peritoneal macrophages: Five week old female BALB/c mice were purchased from Kenya Agricultural Research Institute (KARI) and kept in our animal facility at KEMRI and were used for the in vitro studies of L. major at eight weeks of age. The experiments using mice were done in compliance with animal Care and Use Committee (ACUC) guidelines of Kenya Medical Research Institute (KEMRI). The mice were used as a source of peritoneal macrophages which were harvested and cultured according to the protocol of Lu and Varley (2013). Briefly, thioglycollate-elicited peritoneal macrophages from two female BALB/c mice were collected in 0.8% saline and allowed to adhere onto cover slips placed in 24-well culture plates for 30 min at 37°C in a 4% CO2 atmosphere. Non-adherent cells were then removed and the adhered macrophages were washed twice with 0.9% saline and cultured for 24 h in culture medium (RPMI; Gibco-RBL, Gaithersburg, Md.) supplemented with 10% FBS. The adherent cultured macrophages were then used for infection assays.
Assay of in vitro antiproliferative activity: Inhibition of cell growth was tested in vitro by cultivating promastigotes (1×106) or amastigotes (5×106) in the presence of increasing concentrations of saponin extract, plumbagin extract, trifluralin and acriflavine in 24-well culture dishes (Corning Life Sciences, Corning, N.Y.) for 2, 12 and 24 h. Cell viability was assessed at 24 h by measuring the cleavage of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT; Sigma-Aldrich) with metabolically active cells as described by Mahmoudvand et al. (2015). MTT cleavage was measured by using a multiwell scanning spectrophotometer (Labsystems; Multiskan EX) with a reference wavelength of 690 nm and a test wavelength of 595 nm. Assays were performed in triplicate and results are expressed as the mean percent reduction of parasite numbers compared to untreated control wells calculated for at least three independent experiments. The 50% inhibitory concentration (IC50) was determined from sigmoidal regression of the concentration-response curves.
The inhibition of amastigote intracellular growth was assayed by analyses of the number of infected cells in macrophage monolayers. Murine macrophages were plated in round glass coverslips inside the wells of a 24-well culture dish at a concentration of 5×105 cells per coverslip in RPMI 1640 supplemented with 10% fetal bovine serum, 2 mM L-glutamine and 50 mg of gentamicin mL-1. After 2 h of incubation at 37°C in an atmosphere of 5% CO2, L. major stationary-phase promastigotes were added to the wells (2.5×106 per well) and the cultures were incubated at 33°C in a 5% CO2 atmosphere. After 3 h, free promastigotes were removed by extensive washing with RPMI medium without fetal calf serum and infected cultures were treated with the different drug concentrations for 24 h. The monolayers were washed, fixed and stained with the Instant Prov kit (Newprov, Pinhais, Brazil) and the percentage of infected macrophages was assessed by light microscopy observation by counting 100 cells in triplicate coverslips.
Inhibition of infection in macrophage cells and nitric oxide production: Mouse peritoneal macrophages were obtained as described above. The parasites and/or the macrophages were either not treated or treated with 7.5, 31.25, 62.5, 125, 250 μg of test compound per ml 20 min prior to the macrophage-parasite interactions. The adherent cultured macrophages and the free parasites were washed once and resuspended in fresh culture medium. Dead parasites were removed from the medium by centrifugation (1,000×g, 5 min) and intact living L. major promastigotes in the stationary growth phase were then added to the macrophage culture plate wells. The parasite-macrophage interaction studies were performed at 37°C for 90 min by using parasites and/or macrophages pretreated with the test compounds or macrophages that had already been infected with the parasites for 24 h and then treated with the test compounds. A ratio of 10 promastigotes to 1 macrophage was used for both infection assays. After the interaction assays were done, the coverslips were fixed and Giemsa stained and the percentage of infected macrophages was determined by counting 600 cells in triplicate coverslips. The association indices were determined by multiplying the percentage of infected macrophages by the mean number of parasites per infected cell. Association indices were the number of parasites that actually infected the macrophages. The supernatants from control and L. major-infected macrophages were analyzed for their nitrite contents by the Griess reaction. The absorbance at 550 nm was measured and the concentration of nitrite was calculated by using a linear regression of a standard curve, as described by Ignacio et al. (2001). The mice were first killed to obtain mouse peritoneal macrophages for both infection with Leishmania and nitric oxide measurements, all KEMRI animal house guidelines and institutional policies for the treatment of mice were adhered to.
Cytotoxicity assay: Cytotoxicity was evaluated by cultivating 5×105 murine macrophages or vero cells in 24-well plates for 24 h in the presence of increasing concentrations of trifluralin, plumbagin, acriflavine and saponin. Cell viability was assessed by the MTT assay as described elsewhere (Mahmoudvand et al., 2015) and results are expressed as the percent reduction in cell viability compared to untreated control cultures. The 50% cytotoxic concentration was determined as described above for the IC50 values. The Selectivity Index (SI) was calculated using the following equation:
![]()
Ethical clearance: All procedures were approved by the ethics committees for animal care and research: KEMRI Animal Care and Use Committee (ACUC), Scientific Steering Committee (SSC) and Ethical Review Committee (ERC). The guidelines were strictly adhered to during the research.
Statistical analysis: All experiments were performed in triplicate. The mean and standard deviation of at least three experiments were determined. Statistical analysis of the differences between mean values obtained for the experimental groups was done by Student’s t test. p-values of 0.05 or less were considered significant. To determine the efficacy range of the chemical dosages used, logistic model which can be used to identify non-linear response to the ranges of concentrations, was fitted to the data. The logit model:
![]()
is a general logistic model which takes the form:
![]()
in dose response treatments (Agresti, 1990). The model of efficacy ranges for the chemicals was fitted using GENSTAT (GenStat Release 4.24DE). Model fit was based on residual likelihood ratio chi-square statistic. Data fit that fully described the model was ascertained using scaled deviance. Otherwise all the model fits that did not fully describe the data were based on residual likelihood ratio chi-square statistic based on the Wald statistics. Mean comparisons of significant differences among efficacious dose concentrations were tested using Tukey’s HSD (Bohbot et al., 2004). Significance was accepted at p<0.05.
RESULTS
Efficacy of test compounds on amastigote and promastigote of Leishmania major: Efficacy of plant extracts, acridine and dinitroaniline herbicides on the growth inhibition of amastigote forms is shown in Fig. 1. The amastigote growth inhibition was affected by the various test compounds (p<0.05) after 24 h exposure.
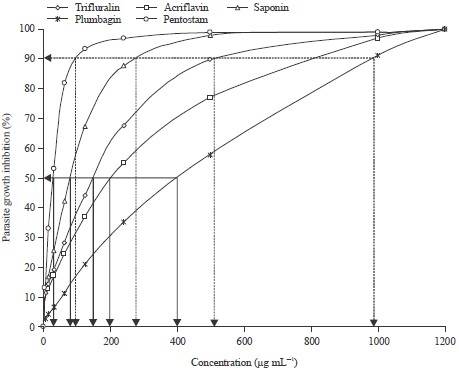 | |
| Fig. 1: | Amastigote growth inhibition (IC50 and IC90 of the test compounds) |
The % growth inhibition estimated for the Amastigote form of parasite using saponin, fully fitted the logistic regression model describing a dose response treatment (R2 = 0.9425). Subjected to 24 h treatment with saponin extract, the model equation was log (r/1-r) = 0.0000+0.6470C-0.0013C2+0.0000C3 and the parameter significance (p-value) obtained for each coefficient were: b0 (P = 0.0354), b1 (P = 0.0000) b2 (P = 0.0000) and b3 (P = 0.0000). The % growth inhibition estimated for the amastigote form of parasite using plumbagin, fully fitted the logistic regression model describing a dose response treatment (R2 = 0.9882). Subjected to 24 h treatment with plumbagin extract, the model equation was log (r/1-r) = 0.0000+0.0968C-0.0001C2+0.0000C3 and the parameter significance (p-value) obtained for each coefficient were: b0 (P = 0.0017), b1 (P = 0.0001), b2 (P = 0.0070) and b3 (P = 0.0004). The % growth inhibition estimated for the amastigote form of parasite using trifluralin, fully fitted the logistic regression model describing a dose response treatment (R2 = 0.9814). Subjected to 24 h treatment with trifluralin, the model equation was log (r/1-r) = 0.0000+0.3287C-0.0004C2+0.0000C3 and the parameter significance (p-value) obtained for each coefficient were: b0 (P = 0 .0000), b1 (P = 0.0000), b2 (P = 0.0001) and b3 (P = 0.0051). The % growth inhibition estimated for the amastigote form of parasite using acriflavine, fully fitted the logistic regression model describing a dose response treatment (R2 = 0.9491). Subjected to 24 h treatment with acriflavine, the model equation was log (r/1-r) = 0.0000+0.2682C-0.0004C2+0.0000C3 and the parameter significance (p-value) obtained for each coefficient were: b0 (P = 0.0007), b1 (P = 0.0000), b2 (P = 0.0064) and b3 (P = 0.0240).
Promastigotes of L. major were cultured with 0, 200, 400, 600, 800 and 1000 μg mL-1 of the test compounds for 24 hours. In the absence of the test compounds L. major promastigotes proliferated over time. In contrast, promastigotes treated with the test compounds gradually decreased in number in a time-dependent fashion. After 24 h of incubation with the test compounds, L. major promastigotes showed a significant reduction in number when compared with the controls. These results demonstrate the inhibition of the growth of L. major promastigotes by the test compounds. Efficacy of plant extracts, acridine and dinitroaniline herbicides on the growth inhibition of promastigote form is shown in Fig. 2. The promastigote growth inhibition was evidently affected by the various test compounds (p<0.05) after 24 h exposure. The % growth inhibition estimated for the promastigote form of parasite using saponin, plumbagin, trifluralin, acriflavine and pentamidine fully fitted the logistic regression model describing a dose response treatment (R2 = 0.9777, 0.9919, 0.9892, 0.9460 and 0.9900), respectively.
Cytotoxicity assay: Results of the efficacy of plant extracts and herbicides on the viability of mammalian cell are shown in Fig. 3. Mammalian cell viability was significantly affected by the various test compounds (p<0.05) after 24 h exposure. The % cell viability estimates of mammalian cells treated with Saponin, fully fitted the logistic regression model describing a dose response treatment (R2 = 0.9756). Subjected to 24 h treatment with saponin extracts, the model equation was log (r/1-r) = 100-14.2924C-1.0440C2+0.1445C3 and the parameter significance (p-value) obtained for each coefficient were: b0 (P = 0.1351), b1 (P = 0.0001) b2 (P = 0.1857) and b3 (P = 0.0091). The % cell viability estimates of mammalian cells treated with plumbagin, fully fitted the logistic regression model describing a dose response treatment (R2 = 0.9815). After treatment for 24 h, with plumbagin extract, the model equation was log (r/1-r) = 100-16.7896C+0.2176C2+0.0413C3 and the parameter significance (p-value) obtained for each coefficient were: b0 (P = 0.0860), b1 (P = 0.0000) b2 (P = 0.7307) and b3 (P = 0.3286). The % cell viability estimates of mammalian cells treated with trifluralin, fully fitted the logistic regression model describing a dose response treatment (R2 = 0.9961).
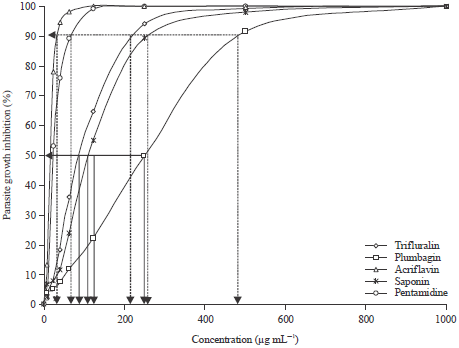 | |
| Fig. 2: | Promastigote growth inhibition (IC50 and IC90 of the test compounds) |
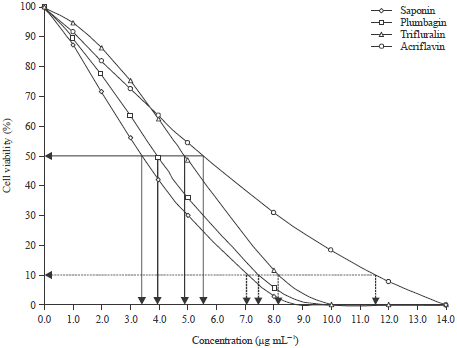 | |
| Fig. 3: | Mammalian cell mortality (CC50 and CC90 of the test compounds) |
After treatment for 24 h with trifluralin, the model equation was log(r/1-r) = 100-2.2533C+2.4044C2+0.1639C3 and the parameter significance (p-value) obtained for each coefficient were: b0 (P = 0.6994), b1 (P = 0.0717) b2 (P = 0.0000) and b3 (P = 0.0000). The % cell viability estimates of mammalian cells treated with acriflavine, fully fitted the logistic regression model describing a dose response treatment (R2 = 0.9755). After treatment for 24 h, with acriflavine, the model equation was log (r/1-r) = 100-10.3497C+0.1760C2+0.0075C3 and the parameter significance (p-value) obtained for each coefficient were: b0 (P = 0.0364), b1 (P = 0.0199) b2 (P = 0.8418) and b3 (P = 0.8877).
| Table 1: | Antileishmanial activity of the test compounds |
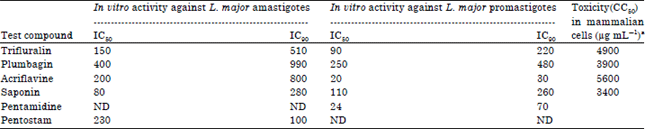 | |
| aNo cytotoxicity on mammalian cells (CC50, >3,400 μg mL-1), ND: Not done | |
| Table 2: | Comparison of values of CC50 for mammalian cells macrophages and IC50 for promastigote form of L. major and their respective Selectivity Indices (SI) |
 | |
| SI = CC50 Vero cells/IC50 promastigotes | |
Antileishmanial activity of test compounds: Table 1 describes the antileishmanial activity of the test compounds. The optimal efficacy of all the test compounds was 100%, albeit the concentrations at optimal efficacy were significantly different among the test compounds. Significant differences were also discerned in CC90 and CC50 after 24 h exposure (p<0.05). Highest CC90 was recorded in acriflavine while saponin had the lowest CC90. The highest CC50 was acriflavine, while the lowest was found to be Saponin.
Mammalian cells macrophages treated with 500, 1000 and 1400 μg mL-1 of the test compounds showed different cell viabilities. Macrophages treated with 500 μg mL-1 of plumbagin, saponin, acriflavine and trifluralin, showed 39, 34, 55 and 50% viable cells, respectively. There were no viable cells at 1000 and 1400 μg mL-1 dose concentration for all the test compounds except for acriflavine which showed 20% viable cells at 1000 μg mL-1 dose concentration (Fig. 3). The toxicity to mammalian cells and the antiprotozoal activity were compared (Table 2) using the Selectivity Index (SI) ratio: CC50 mammalian cells/ IC50 Leishmania amastigotes, where CC50 is the 50% of the cytotoxic concentration. A value greater than 1 is considered to be more selective against L. major amastigotes and a value lower than 1 is considered to be more selective to the cells. In this case, the S.I were, 54.4, 15.6, 280 and 30 for trifluralin, plumbagin, acriflavine and saponin, respectively, demonstrating that all the test compounds were less toxic to mammalian cell than to the protozoans (Table 2). Therefore, all the test compounds were more selective against the parasites than the macrophages.
Inhibition of infection in macrophage cells: Survival response of L. major in macrophage treatments is provided in Fig. 4. Survival under test compounds followed logistic dose-response curve. At 7.5 μg mL-1, survival under trifluralin, acriflavine and Saponin were not significantly (p>0.05) different from each other. The survival under the three treatment compounds were however, significantly higher than that of pentostam and plumbagin. Survival of L. major at a concentration 31.25-125 μg mL-1 were all significantly different; those treated using trifluralin and pentostam had the highest and lowest survival respectively. No L. major survived at 250 μg mL-1 in the test compounds.
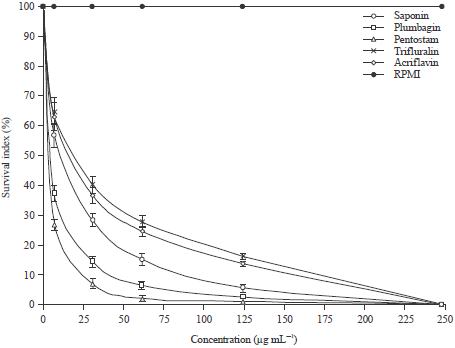 | |
| Fig. 4: | Survival of L. major in macrophage at different dose concentration of the test compounds |
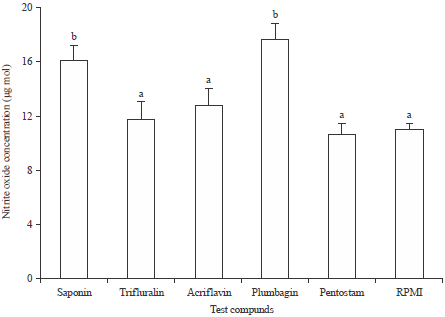 | |
| Fig. 5: | Concentration of nitric oxide produced by macrophages induced by the test compounds (Each bar represents the mean ± standard error of at least three independent experiments which were performed in triplicate. Mean values marked with different letters (a or b) are significantly different at α = 0.05) |
Nitric Oxide (NO) production: To obtain the requisite concentration of nitrite, a graph of absorbance against concentration was plotted (Fig. 6). The curve fit for the standard was highly significant at 99% indicating proper preparations of standards. Results showing production of nitrite oxide among the test compounds is shown in Fig. 5. There were significant differences in the nitrite oxide produced among the test compounds (F = 8.788, df = 5, P = 0.005).
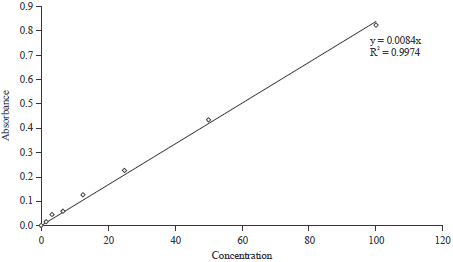 | |
| Fig. 6: | Nitric oxide production |
Plumbagin produced the highest nitrite oxide followed by saponin. There were however no significant differences (p>0.05) observed between the RPMI, pentostam, acriflavine and trifluralin. Probably our minimum concentration of 1.56 μg mL-1 was sufficient to liberate upto 100% of nitrite in the tissues.
DISCUSSION
Efforts to find new chemotherapeutics for leishmaniasis have been ongoing for decades and numerous potentially new drug candidates and/or putative drug targets have indeed been proposed by many authors (Martin-Quintal et al., 2009; Makwali et al., 2012; Falodun et al., 2014). In this study, antileishmanial activity of the test compounds in order of potency was as follows in decreasing strength: saponin, acriflavine, trifluralin and plumbagin. These data showed that, in general, these test compounds are efficient against Leishmania species without toxicity to peritoneal macrophages (CC50, >3,400 μg mL-1). The basic treatment for leishmaniasis consists of the administration of pentavalent antimonials that were developed more than 50 years ago; however, serious toxic effects and the emergence of resistance are limiting the drugs’ usefulness (Singh et al., 2012). Although, there are several compounds being studied, it is important to search for new compounds with high potency and low toxicity.
The in vitro IC50 values for trifluralin, plumbagin, acriflavine and saponin are 90, 250, 20 and 110 μg mL-1, respectively. The selectivity indices of these compounds were 54.4, 15.6, 280 and 30 μg mL-1, respectively, in the mammalian cell model used which are rather high selectivity indices. According to Lenta et al. (2007) selectivity index values higher than 10.0 suggest a better safety of a product for use in mammalian hosts. These results indicate that acridine and dinitroaniline herbicides with the plant extracts are promising lead compounds for development as novel antileishmanial agents. Mokoka et al. (2014) reported that good selectivity considered together with modification of 12-methoxycarnosic acid isolated from Salvia repens could result in the development of new analogues with much more improved antileishmanial activity against axenic L. donovani amastigotes. In the results of this study, saponin displayed marked in vitro anti-amastigote activity with an IC50 value of 80 μg mL-1 against L. major amastigotes at concentrations of 10-1200 μg mL-1 and no toxicity to mammalian macrophages at concentrations <3.4 mg mL-1. In contrast, the antimonial pentostam had an IC50 value of 130 μg mL-1 against L. major amastigotes at concentrations of 10-1200 μg mL-1 but was found to be toxic for macrophages at concentrations >1.0 mg mL-1. Thus, in vitro, saponin exhibited a more powerful anti-amastigote effect and a lower cytotoxicity to mammalian macrophages than pentostam. Saponins are a highly diverse group of glycosides of plant origin. Their main biologic actions are changes in membrane permeability and pore formation (Melzig et al., 2001). Saponins are also hemolytic, probably as a result of their interactions with steroids, especially cholesterol. The amount of cholesterol in the membrane has been shown to be important for this interaction (Gogelein and Huby, 1984). Taking all this information into consideration, the high potency of saponin extract used in this study may be attributed to these factors.
The potent in vitro activity of the test compounds (trifluralin, plumbagin, acriflavine and saponin) against intracellular L. major amastigotes (IC50 = 150, 400, 200 and 80 μg mL-1, respectively) in the absence of obvious cytotoxicity on murine macrophage cells (CC50 >3,400 μg mL-1), which formed the basis for advanced exploration of this promising lead activity should not be used as the sole criterion for lead selection. Selectivity is another, probably more relevant characteristic for defining lead candidates (Maes et al., 2004). These results showed antileishmanial activity for all compounds tested against promastigotes, with most IC50 values being lower than for the standard drug pentostam. However, pentamidine showed good anti-promastigote activity with an IC50 value of 24 μg mL-1. The antileishmanial activities of these compounds used in this study were comparable to those reported by Ribeiro et al. (2015), who showed that isolated compounds of organic fractions derived from Stryphnodendron obovatum stem bark were more potent than the positive control, rutin. These findings exhibited that amastigotes were more susceptible to the test compounds than the promastigotes. This difference in their susceptibility could be related to their structural, biochemical and morphological features as previously demonstrated by Shokri et al. (2012).
Saponin had the lowest IC50 value of any compounds tested against L. major amastigotes, with an IC50 value of 80 μg mL-1. It was at least 5-fold more effective than the standard drug, pentostam. The intracellular amastigotes have also been shown to express different enzymatic patterns, which could improve the parasite defense (Chan-Bacab and Pena-Rodriquez, 2001). Incubation of L. major-infected macrophages with the test compounds demonstrated significant antileishmanial activity at the highest concentration. The relationship between mammalian toxicity and antiparasitic effect, given by the S.I., demonstrated that saponin was at least 30-fold more harmful to intracellular amastigotes than to mammalian cells. Acriflavine had the lowest IC50 value of any compounds tested against L. major promastigotes, with an IC50 value of 20 μg mL-1. It was at least 1.2-fold more effective than the standard drug, pentamidine. However, antileishmanial activity against promastigotes does not guarantee activity against intracellular amastigotes, the clinically more relevant form of the parasite. Acriflavine had an IC50 value of 200 μg mL-1 against amastigotes.
Trifluralin showed good antiparasitic activity against L. major parasites. This therefore, strengthens the possibility of and enhances the impetus for developing trifluralin as a therapeutic agent for the treatment of leishmaniasis. Trifluralin is not genotoxic, as indicated by the standard tests, such as the salmonella/microsome mutagenicity test, the yeast genotoxicity assay, the mouse lymphoma assay, the sister chromatid exchange assay, the in vitro chromosome aberration assay and the in vivo micronucleus test for mammalian genotoxicity (Zaidenberg et al., 2006). Oncogenicity tests in rats and mice and embryotoxicity tests in rats and rabbits revealed neither carcinogenic nor teratogenic potential when taken orally (Marra et al., 2002). In addition, tubulins as new targets affected by trifluralin, are encoded by multiple copies of genes and thus drug resistance may be less likely to develop. Regarding human health (Brooks, 1973) there are no reported cases of long-term health effects in man due to trifluralin or its formulation. In acute toxicity (poisoning) most incidents of trifluralin exposure reported in man have involved dermal and eye irritation and nausea. Dizziness and headache, fever or chills and muscle aches or weakness have also been reported. Despite the eventual toxicity incidents reported, the therapeutic possibilities are evident, as shown by already obtained results.
Macrophages, the target cells in therapy of leishmaniasis, play an important role in the immunological control of intracellular parasites through the production of cytokines and oxygen metabolites (Kalyanaraman, 2004). One of the main mechanisms is the up-regulation of nitric oxide inside the cell which is an effective mediator of amastigote killing (Mauel and Ransijn, 1997). Plumbagin produced the highest nitrite oxide followed with saponin. There were however no significant differences (p>0.05) observed between the pentostam, acriflavine, trifluralin and the controls. These results indicate that these compounds might have a specific antiparasitic activity rather than causing activation of NO production by macrophages per se. These results are consistent to results reported by Ribeiro et al. (2014), that suggested that other mechanisms than NO production may well be involved in the elimination of parasites by Dipteryx alata and Jacaranda cuspidifolia without requiring macrophage activation via iNOS expression. In addition, efficacy against extracellular promastigotes with trifluralin, acriflavine and pentamidine having an IC50 value of 90, 20 and 24 μg mL-1, respectively supports the hypothesis of a specific antiparasitic activity for these compounds.
It has been reported that naphthoquinones extracted from several members of the plant family Bignoniaceae possess activity both in vitro and in vivo against various strains of Leishmania (Iwu et al., 1994). Lapachol, diospyrin, plumbagin and b-lapachone are the most active members of this group (Teixeira et al., 2001). In this study, plumbagin exhibited leishmanicidal activity in vitro. The mechanism by which plumbagin induces lysis of intracellular amastigotes in vitro is not clear. However, a characteristic of the naphthoquinones is that they interfere with the oxygen metabolism of the tumour cell, blocking cell respiration and generating free oxygen radicals (Oswald et al., 1994). Besides free oxygen radicals, there is evidence that Nitric Oxide (NO) production which follows the induction of nitric oxide synthetase by IFN-g, plays an important role for the death of intracellular amastigotes in murine macrophages (Teixeira et al., 2001). NO is considered the most important metabolite involved in Leishmania killing in mice (Assreuy et al., 1994).
Possibly, plumbagin inhibits NO-dependent physiological and/or pathophysiological actions in vivo and since it showed leishmanicidal activity in vitro then it could have acted directly on the parasite and not necessarily through the activation of macrophages as is the case in vivo. This is the first report on plumbagin that shows direct inhibition of L. major amastigotes. It has been estimated that 60% of people living in developing countries depend on traditional medicine for their primary health care. Therefore, new therapies derived from medicinal herbs offer a promising and practical avenue to the development of new drugs for the most needed in Leishmania-endemic countries.
CONCLUSION
This study provides valuable data that the biocides and herbicides used exhibit antileishmanial properties without any toxicity for mammalian and murine cells. These findings therefore, represent an important new asset in the search for novel antileishmanial drugs. Further laboratory and clinical studies of these study compounds are required in order to understand their antileishmanial principles.
ACKNOWLEDGMENTS
The authors thank Mr. Lucas Ogutu of Kenya Medical Research Institute (KEMRI) for rearing the animals used in this study. We wish to appreciate the support given by Moi University as the host University and Center for Biotechnology Research and Development (CBRD), KEMRI for providing the laboratory space to carry out the investigations.
REFERENCES
- Adebayo, L.O., D. Suleman and A.A. Samson, 2013. Natural products in antileishmanial drug discovery: A review. J. Asian Scient. Res., 3: 157-173.
Direct Link - Assreuy, J., F.Q. Cunha, M. Epperlein, A. Noronha-Dutra, C.A. O'Donnell, F.Y. Liew and S. Moncada, 1994. Production of nitric oxide and superoxide by activated macrophages and killing of Leishmania major. Eur. J. Immunol., 24: 672-676.
CrossRefDirect Link - Bohbot, V.D., G. Iaria and M. Petrides, 2004. Hippocampal function and spatial memory: Evidence from functional neuroimaging in healthy participants and performance of patients with medial temporal lobe resections. Neuropsychology, 18: 418-425.
CrossRefDirect Link - Chan-Bacab, M.J. and L.M. Pena-Rodriguez, 2001. Plant natural products with leishmanicidal activity. Nat. Prod. Rep., 18: 674-688.
PubMed - Falodun, A., V. Imieje, O. Erharuyi, A. Joy and P. Langer et al., 2014. Isolation of antileishmanial, antimalarial and antimicrobial metabolites from Jatropha multifida. Asian Pac. J. Trop. Biomed., 4: 374-378.
CrossRefDirect Link - Garnier, T., A. Mantyla, T. Jarvinen, M.J. Lawrence, M.B. Brown and S.L. Croft, 2007. Topical buparvaquone formulations for the treatment of cutaneous leishmaniasis. J. Pharm. Pharmacol., 59: 41-49.
CrossRefDirect Link - Gogelein, H. and A. Huby, 1984. Interaction of saponin and digitonin with black lipid membranes and lipid monolayers. Biochimica Biophysica Acta (BBA)-Biomembr., 773: 32-38.
CrossRefDirect Link - Gomes, L.I., F.M. Gonzaga, E. de Morais-Teixeira, B.S. de Souza-Lima, V.V. Freire and A. Rabello, 2012. Validation of quantitative real-time PCR for the in vitro assessment of antileishmanial drug activity. Exp. Parasitol., 131: 175-179.
CrossRefDirect Link - González-Coloma, A., M. Reina, C. Sáenz, R. Lacret and L. Ruiz-Mesia et al., 2012. Antileishmanial, antitrypanosomal and cytotoxic screening of ethnopharmacologically selected peruvian plants. Parasitol. Res., 110: 1381-1392.
CrossRefDirect Link - Ignacio, S.R.N., J.L.P. Ferreira, M.B. Almeida and C.F. Kubelka, 2001. Nitric oxide production by murine peritoneal macrophages in vitro and in vivo treated with Phyllanthus tenellus extracts. J. Ethnopharmacol., 74: 181-187.
CrossRefDirect Link - Iwu, M.M., J.E. Jackson and B.G. Schuster, 1994. Medicinal plants in the fight against leishmaniasis. Parasitol. Today, 10: 65-68.
CrossRefPubMedDirect Link - Kalyanaraman, B., 2004. Nitrated lipids: A class of cell-signaling molecules. Proc. Natl. Acad. Sci. USA., 101: 11527-11528.
CrossRefDirect Link - Lenta, B.N., C. Vonthron-Senecheau, R.F. Soh, F. Tantangmo and S. Ngouela et al., 2007. In vitro antiprotozoal activities and cytotoxicity of some selected Cameroonian medicinal plants. J. Ethnopharmacol., 111: 8-12.
CrossRefDirect Link - Lu, M. and A.W. Varley, 2013. Harvest and culture of mouse peritoneal macrophages. Bio-Protocol, Vol. 3, No. 22.
Direct Link - Maes, L., N. Germonprez, L. Quirijnen, L. Van Puyvelde, P. Cos and D.V. Berghe, 2004. Comparative activities of the triterpene saponin Maesabalide III and liposomal amphotericin B (AmBisome) against Leishmania donovani in hamsters. Antimicrob. Agents Chemother., 48: 2056-2060.
CrossRefDirect Link - Mahmoudvand, H., F. Ezzatkhah, F. Sharififar, I. Sharifi and E.S. Dezaki, 2015. Antileishmanial and cytotoxic effects of essential oil and methanolic extract of Myrtus communis L. Korean J. Parasitol., 53: 21-27.
CrossRefDirect Link - Makwali, J.A., F.M. Wanjala, J.C. Kaburi, J. Ingonga, W.W. Byrum and C.O. Anjili, 2012. Combination and monotherapy of Leishmania major infection in BALB/c mice using plant extracts and herbicides. J. Vector Borne Dis., 49: 123-130.
PubMedDirect Link - Marra, C.A., A. Zaidenberg, M.J.T. de Alaniz and H. Buschiazzo, 2002. The restoring effect of trifluralin and benznidazole on the abnormal fatty-acid pattern induced by Trypanosoma cruzi in the liver microsomes of infected mice. Ann. Trop. Med. Parasitol., 96: 249-264.
CrossRefDirect Link - Mauel, J. and A. Ransijn, 1997. Leishmania spp.: Mechanisms of toxicity of nitrogen oxidation products. Exp. Parasitol., 87: 98-111.
CrossRefDirect Link - Melzig, M.F., G. Bader and R. Loose, 2001. Investigations of the mechanism of membrane activity of selected triterpenoid saponins. Planta Medica, 67: 43-48.
CrossRefPubMedDirect Link - Mokoka, T.A., X.K. Peter, G. Fouche, N. Moodley and M. Adams et al., 2014. Antileishmanial activity of 12-methoxycarnosic acid from Salvia repens Burch. ex Benth. (Lamiaceae). South Afr. J. Bot., 90: 93-95.
CrossRefDirect Link - Murray, H.W., J.D. Berman, C.R. Davies and N.G. Saravia, 2005. Advances in leishmaniasis. Lancet, 366: 1561-1577.
CrossRefPubMedDirect Link - Oryan, A., 2015. Plant-derived compounds in treatment of leishmaniasis. Iran. J. Vet. Res., 16: 1-19.
Direct Link - Oswald, I.P., T.A. Wynn, A. Sher and S.L. James, 1994. NO as an affector molecule of parasite killing: Modulation of its synthesis by cytokines. Comp. Biochem. Physiol. Part C: Pharmacol. Toxicol. Endocrinol., 108: 11-18.
CrossRefDirect Link - Ribeiro, T.G., A.M. Nascimento, B.O. Henriques, M.A. Chavez-Fumagalli and J.R. Franca et al., 2015. Antileishmanial activity of standardized fractions of Stryphnodendron obovatum (Barbatimao) extract and constituent compounds. J. Ethnopharmacol., 165: 238-242.
CrossRefDirect Link - Ribeiro, T.G., M.A. Chavez-Fumagalli, D.G. Valadares, J.R. Franca and P.S. Lage et al., 2014. Antileishmanial activity and cytotoxicity of Brazilian plants. Exp. Parasitol., 143: 60-68.
CrossRefDirect Link - Shokri, A., I. Sharifi, A. Khamesipour, N. Nakhaee and M.F. Harandi et al., 2012. The effect of verapamil on in vitro susceptibility of promastigote and amastigote stages of Leishmania tropica to meglumine antimoniate. Parasitol. Res., 110: 1113-1117.
CrossRefDirect Link - Singh, N., M. Kumar and R.K. Singh, 2012. Leishmaniasis: Current status of available drugs and new potential drug targets. Asian Pac. J. Trop. Med., 5: 485-497.
CrossRefDirect Link - Teixeira, M.J., Y.M. de Almeida, J.R. Viana, J.G.H. Filha and T.P. Rodrigues et al., 2001. In vitro and in vivo leishmanicidal activity of 2-hydroxy-3-(3-methyl-2-butenyl)-1,4-naphthoquinone (lapachol). Phytother. Res., 15: 44-48.
CrossRefPubMedDirect Link - Zaidenberg, A., T. Luong, D. Lirussi, J. Bleiz and M.B. Del Buono et al., 2006. Treatment of experimental chronic chagas disease with trifluralin. Basic Clin. Pharmacol. Toxicol., 98: 351-356.
CrossRefDirect Link - Zauli-Nascimento, R.C., D.C. Miguel, J.K.U. Yokoyama-Yasunaka, L.I.A. Pereira and M.A.P. de Oliveira et al., 2010. In vitro sensitivity of Leishmania (Viannia) braziliensis and Leishmania (Leishmania) amazonensis Brazilian isolates to meglumine antimoniate and amphotericin B. Trop. Med. Int. Health, 15: 68-76.
CrossRefDirect Link - Martin-Quintal, Z., R. Moo-Puc, F. Gonzalez-Salazar, M.J. Chan-Bacab, L.W. Torres-Tapia and S.R. Peraza-Sanchez, 2009. In vitro activity of Tridax procumbens against promastigotes of Leishmania mexicana. J. Ethnopharmacol., 122: 463-467.
CrossRefDirect Link