
L.K. Darusman
Biopharmaca Research Center, Bogor Agricultural University, Jl Taman Kencana No. 3, Bogor, 16151, Indonesia
I. Batubara
Biopharmaca Research Center, Bogor Agricultural University, Jl Taman Kencana No. 3, Bogor, 16151, Indonesia
T. Mitsunaga
Faculty of Applied Biological Science, Gifu University, 1-1 Yanagido, Gifu, 501-1193, Japan
M. Rahminiwati
Biopharmaca Research Center, Bogor Agricultural University, Jl Taman Kencana No. 3, Bogor, 16151, Indonesia
E. Djauhari
Biopharmaca Research Center, Bogor Agricultural University, Jl Taman Kencana No. 3, Bogor, 16151, Indonesia
K. Yamauchi
Faculty of Applied Biological Science, Gifu University, 1-1 Yanagido, Gifu, 501-1193, Japan
Research Journal of Medicinal Plants
Year: 2012 | Volume: 6 | Issue: 8 | Page No.: 615-620







ABSTRACT
Intsia palembanica is the most potential plant having tyrosinase inhibitor over 30 Indonesian plants that have been searched. It’s active compound namely (-)-robidanol, (+)-epirobidanol and 4’-dehydroxyrobidanol had been identified and showed the tyrosinase inhibitory activity. The study aims to obtain the kinetic enzyme inhibition type of the three active compounds. About 3 g of I. palembanica methanol extract was fractionated using n-hexane soluble part, ethyl-acetate soluble part and aqueous soluble part. Compounds were isolated for the most active part (ethyl-acetate soluble part) by silica gel column chromatography. Purification of fractions was performed using preparative High Performance Liquid Chromatography (HPLC). Kinetic inhibition against tyrosinase on monophenolase and diphenolase were analyzed by Lineweaver-Burk plot. The results showed that three active compounds showed different kinetic inhibition type on monophenolase and diphenolase. The kinetic characteristics of (-)-robidanol, the most active inhibitor, on monophenolase were Km increased and Vmax did not changed while on diphenolase were Vmax decreased and Km did not change. In conclusion, (-)-robidanol is a competitive inhibitor for monophenolase and a non-competitive inhibitor for diphenolase. The kinetic data is useful for further research on mechanism of action of the whitening agent that will be formulated.
PDF Abstract XML References Citation
Received: February 14, 2012;
Accepted: May 30, 2012;
Published: December 05, 2012
How to cite this article
L.K. Darusman, I. Batubara, T. Mitsunaga, M. Rahminiwati, E. Djauhari and K. Yamauchi, 2012. Tyrosinase Kinetic Inhibition of Active Compounds from Intsia palembanica. Research Journal of Medicinal Plants, 6: 615-620.
URL: https://scialert.net/abstract/?doi=rjmp.2012.615.620
URL: https://scialert.net/abstract/?doi=rjmp.2012.615.620
INTRODUCTION
In the search for bioactive compounds with tyrosinase inhibitory activity from Indonesian medicinal plants, previously the research focused on Intsia palembanica (Batubara et al., 2010, 2011). Three active compounds isolated from I. palembanica had tyrosinase inhibitory activities, namely (-)-robidanol, (+)-epirobidanol and 4’-dehydroxyrobidanol (Fig. 1).
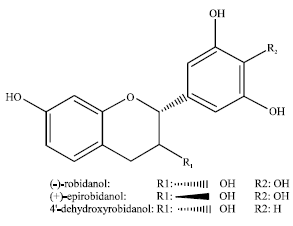 | |
| Fig. 1: | Structure of three flavanols from I. palembanica |
Since the active compounds reported have enzyme inhibitory activity, it is interesting to know the biochemical kinetic properties of an enzyme in the presence of active compounds. Biochemical kinetic properties like Km and Vmax are the biochemical properties most determined on an enzyme in the presence of some compounds or extracts, such as on adenosine deaminase in the presence of garlic and red clover (Avci et al., 2005), catalase in the presence of Brassica oleracea capitata (Gholamhoseinian et al., 2006), lysozyme in the presence of glyoxime and dichloroglyoxime (Ranjbar et al., 2006), α-glucosidase in the presence of Ficus deltoidea (Adam et al., 2010), alanine dehydrogenase in the presence of Thielaviopsis paradoxa (Al-Onazi et al., 2011) and on NADP+-linked-isocitrate dehydrogenase in the presence of Phaseolus mungo L. (Srivastava et al., 2011). The kinetic parameters of tyrosinase such as Km and Vmax have also been reported such as the Km and Vmax values of tyrosinase isolated from potato and edible fungi (Yang and Wu, 2006).
Information about kinetics inhibition of tyrosinase inhibitor worth to determined, because there were many compounds natural and synthetic have activity of tyrosinase inhibitor but only some of them have the information about kinetics behavior (Chang, 2009). Enzyme inhibitors are classified into four types, namely competitive, uncompetitive, mixed (competitive/uncompetitive) and non-competitive inhibitors (Chang, 2009). Since the IC50 values, the usual expression for inhibitor strength, of the three flavanols isolated from I. palembanica were strong (Batubara et al., 2011), it is interesting to find the kinetic inhibition of the compounds. The aim of this research was to classify inhibition strength of the three isolated flavanols from I. palembanica, by its kinetic inhibition on Lineweaver Burk plot.
MATERIALS AND METHODS
Plant materials, extraction, isolation and identification of active compounds: The research was performed from October 2010 till April 2011. I. palembanica samples were collected from Samarinda, East Kalimantan, Indonesia. All the extraction, isolation and identification processes were performed as in our previous report (Batubara et al., 2010, 2011). Briefly, 3 g of I. palembanica methanol extract was fractionated using n hexane, ethylacetate and water to result to n-hexane soluble part, ethylacetate soluble part and aqueous soluble part. n-hexane soluble part (13 mg) had no tyrosinase inhibition activity, while ethylacetate soluble part (1.4 g) and aqueous soluble part (1.6 g) had tyrosinase activity (IC50 value for diphenolase were 3,97 and 4,34 ppm, respectively). Ethyl acetate soluble part which more active compared to aqueous soluble part was separated by silica gel column chromatography with hexane, ethyl acetate and methanol as the developing solvents and resulted to 26 fractions. Fraction 4-8 purified using preparative HPLC with reversed phase column Inertsil ODS-3 (GL Sciences 10 mm idx200 mm) monitored at 280 nm. The solvent system used was as follows: a gradient program for 45 minutes from 5% to 100% of methanol in TFA 0.05% (in water) at flow rate of 3 mL min-1. This separation step gave (-)-robidanol (15.8 mg), (+)-epirobidanol (47.4 mg) and 4'-dehydroxyrobidanol (10.5 mg) (Fig. 1). The structures of the isolated compounds were determined by comparison of their spectroscopic data with those reported in the literature. 1H- and 13C- NMR were recorded with a JEOL ECP 600 MHz spectrometer with TMS as the internal reference and chemical shifts expressed in δ (ppm). All spectroscopy data was reported on our previous report (Batubara et al., 2011).
The kinetic inhibition test against mushroom tyrosinase: This assay was performed similar to the inhibition assay which we published previously (Batubara et al., 2011). The concentration of pure compound used in this experiment was the same with IC50 (monophenolase: (-)-robidanol: 8.7 μM, (+)-epirobidanol: 20.2 μM, 4’-dehydroxyrobidanol: 15.2 μM; diphenolase: (-)-robidanol: 26.6 μM, (+)-epirobidanol: 178.5 μM, 4’-dehydroxyrobidanol: 50.0 μM). Concentrations of substrate were 0, 0.2, 0.4, 0.6, 0.8, 1.2, 1.6 and 2.0 mM for L-tyrosine and 0, 1, 2, 4, 6, 8, 10 and 12 mM for L-DOPA. The Lineweaver and Burk plot of 1/Absorbance versus 1/[substrate] was used to determine the kinetic parameters of tyrosinase biocatalysis with or without the pure compound. The Michaelis constants Km and V max were calculated from the plot.
RESULTS AND DISCUSSION
(-)-Robidanol, (+)-epirobidanol and 4’-dehydroxyrobidanol are compounds in group of catechin like compounds, flavanol. Unlike catechin, the three isolated compounds are more active on monophenolase and diphenolase activities of tyrosinase (Batubara et al., 2011). The kinetic type of catechin as tyrosinase inhibitor is a competitive type on monophenolase (Kim and Uyama, 2005) and mixed type inhibitor on diphenolase (Nirmal and Benjakul, 2012).
The inhibition kinetics of (-)-robidanol, (+)-epirobidanol and 4’-dehydroxyrobidanol were analyzed using Lineweaver-Burk plot on their IC50 values. The results for monophenolase and diphenolase activities are presented in Fig. 2a and b, respectively. In monophenolase activity (substrate: L-tyrosine), Km value of (+)-epirobidanol was increased while Vmax decreased. The results showed that (+)-epirobidanol is a mixed inhibitor type. Unlike (+)-epirobidanol, (-)-robidanol and 4’-dehydroxyrobidanol are competitive inhibitors because the Km increased while Vmax did not changed. The inhibition type of (-)-robidanol and 4’-dehydroxyrobidanol are the same with kojic acid on monophenolase activity (Chen et al., 1991), catechin (Kim and Uyama, 2005) and gallocatechin-3-O-gallate (No et al., 1999). Different with the isolated compounds and kojic acid, sesamol from sesame seed had been reported as a non-competitive inhibitor on monophenolase (Kumar et al., 2011).
In diphenolase (substrate L-DOPA), the inhibition type of (+)-epirobidanol and 4’-dehydroxyrobidanol are uncompetitive type because Km value and Vmax decrease, while (-)-robidanol is non-competitive inhibitor (Vmax decreased and Km did not change). The types of inhibition of 3 isolated compounds on diphenolase are different with catechin, a mixed type inhibitor which could bind with enzyme and enzyme-substrate complex of tyrosinase (Nirmal and Benjakul, 2012).
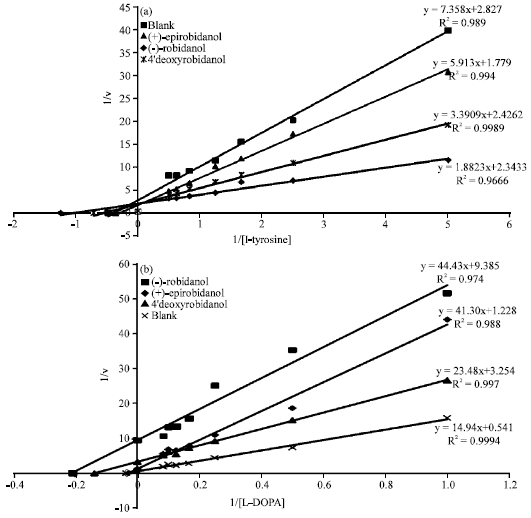 | |
| Fig. 2(a-b): | Lineweaver-Burk plot of active compounds from I. palembanica on (a) monophenolase and (b) diphenolase |
Most information relating to the type of inhibitor for diphenolase are existing especially as a competitive inhibitor, for example, flavonols such as quercetin, galangin, fisetin, 3,7,4’-trihydroxyflavone and morin (Xie et al., 2003), flavones such as norartocarpetin (Ryu et al., 2008), isoflavones such as 6-hydroxydaidzein (Chang et al., 2007) and also sesamol, a phenolic compound formed from mushroom (Kumar et al., 2011). The data of inhibition type of 3 isolated compounds from I. palembanica added the type of inhibitor for diphenolase activity of tyrosinase.
The inhibition kinetics of monophenolase and diphenolase are different because tyrosinase consists of two active sites. The formations on monophenolase and diphenolase reactions were different. On monophenolase reaction, hydrophobic interaction reaction in substrate took place between one Cu atom on tyrosinase while on diphenolase reaction, hydroxyl group of L-DOPA will be deprotonated by peroxide ion bound between the two Cu atoms in tyrosinase (Ismaya et al., 2011).
Competitive inhibitor of tyrosinase means that the inhibitor binds with free enzyme. On the other hand, competitive inhibitors also can prevent substrate to bind with enzyme or inhibitor can be a copper chelator. The enzyme-substrate complex binding only showed for an uncompetitive inhibitor type. While for a mixed type inhibitor can bind to a free enzyme and also to the enzyme-substrate complex with a free enzyme (Chang, 2009).
CONCLUSION
The three active compounds showed different kinetic inhibition type on monophenolase and diphenolase. The most active compound, (-)-robidanol, had a competitive inhibitor type for monophenolase and a non-competitive inhibitor type for diphenolase.
ACKNOWLEDGMENTS
We are thankful to Ms. Mary Grace Saldajeno for grammatical correction of the manuscript. This work was supported by Higher Education Directorate of the National Education Department of Republic of Indonesia (Hibah kompetitif penelitian untuk publikasi international No: 4472H/PP/DP2M/VI/2010) and the Chemistry of Natural Products Laboratory, Gifu University, Japan.
REFERENCES
- Adam, Z., S. Khamis, A. Ismail and M. Hamid, 2010. Inhibitory properties of Ficus deltoidea on α-glucosidase activity. Res. J. Med. Plant, 4: 61-75.
CrossRef - Al-Onazi, M., S. Al-Dahain, A. El-Ansary and N. Marraiki, 2011. Isolation and characterization of Thielavinopsis paradoxa L-alanine dehydrogenase. Asian J. Applied Sci., 4: 702-711.
CrossRef - Avci, A., M. Kacmaz, M. Kavutcu, E. Gocmen and I. Durak, 2005. Effects of an antioxidant extract on adenosine deaminase activities in cancerous human liver tissues. Int. J. Cancer Res., 1: 53-56.
CrossRefDirect Link - Batubara, I., L.K. Darusman, T. Mitsunaga, M. Rahminiwati and E. Djauhari, 2010. Potency of Indonesian medicinal plants as tyrosinase inhibitor and antioxidant agent. J. Biol. Sci., 10: 138-144.
CrossRef - Chang, T.S., H.Y. Ding, S.S.K. Tai and C.Y. Wu, 2007. Mushroom tyrosinase inhibitory effects of isoflavones isolated from soygerm koji fermented with Aspergillus oryzae BCRC 32288. Food Chem., 105: 1430-1438.
CrossRefDirect Link - Chen, J.S., C.I. Wei and M.R. Marshall, 1991. Inhibitory mechanism of kojic acid on polyphenol oxidase. J. Agric. Food Chem., 39: 1897-1901.
CrossRefDirect Link - Gholamhoseinian, A., N. Ghaemi and F. Rahimi, 2006. Partial purification and properties of catalase from Brassia oleracea capitata. Asian J. Plant Sci., 5: 827-831.
CrossRefDirect Link - Kim, Y.J. and H. Uyama, 2005. Tyrosinase inhibitors from natural and synthetic sources: Structure, inhibition mechanism and perspective for the future. Cell. Mol. Life Sci., 62: 1707-1723.
Direct Link - Kumar, C.M., U.V. Sathisha, S. Dharmesh, A.G.A. Rao and S.A. Singh, 2011. Interaction of sesamol (3,4-methylenedioxyphenol) with tyrosinase and its effect on melanin synthesis. Biochimie, 93: 562-569.
CrossRef - No, J.K., D.Y. Soung, Y.J. Kim, K.H. Shim and Y.S. Jun et al., 1999. Inhibition of tyrosinase by green tree components. Life Sci., 65: PL241-PL246.
CrossRefDirect Link - Ranjbar, B., S. Afshar, A. Kakanejadifard, K. Khajeh, H. Naderi-Manesh, L. Hassani and N. Alizadeh, 2006. Effects of glyoxime and dichloroglyoxime on lysozyme: Kinetic and structural studies. Asian J. Biochem., 1: 153-161.
CrossRefDirect Link - Ryu, Y.B., T.J. Ha, M.J. Curtis-Long, H.W. Ryu, S.W. Gal and K.H. Park, 2008. Inhibitory effects on mushroom tyrosinase by flavones from the stem barks of Morus lhou (S.) Koidz. J. Enzyme Inhib. Med. Chem., 23: 922-930.
PubMedDirect Link - Srivastava, P.K., G.K. Srivastava, I. Mani, S. Yadav and A. Anand, 2011. Role of metabolites and significance of sh groups in the action of NADP+-linked isocitrate dehydrogenase of urdbean seeds (Phaseolus mungo L.). Asian J. Biochem., 6: 181-190.
CrossRefDirect Link - Yang, Z. and F. Wu, 2006. Catalytic properties of tyrosinase from potato and edible fungi. Biotechnology, 5: 344-348.
CrossRefDirect Link - Batubara, I., L.K. Darusman, T. Mitsunaga, H. Aoki, M. Rahminiwati, E. Djauhari and K. Yamauchi, 2011. Flavonoid from Intsia palembanica as skin whitening agent. J. Biol. Sci., 11: 475-480.
CrossRefDirect Link - Chang, T.S., 2009. An updated review of Tyrosinase inhibitors. Int. J. Mol. Sci., 10: 2440-2475.
CrossRefDirect Link