
Amir Rezazadeh
Department of Horticulture, Gorgan University of Agricultural Sciences and Natural Resources, Gorgan, Iran
Azim Ghasemnezhad
Department of Horticulture, Gorgan University of Agricultural Sciences and Natural Resources, Gorgan, Iran
Mojtaba Barani
Department of Soil Science, Gorgan University of Agricultural Sciences and Natural Resources, Gorgan, Iran
Telmah Telmadarrehei
Department of Plant Protection, College of Agronomic Sciences, Sari Agricultural Sciences and Natural Resources University, Sari, Iran
Research Journal of Medicinal Plants
Year: 2012 | Volume: 6 | Issue: 3 | Page No.: 245-252







ABSTRACT
Due to high content of phenolic compounds that exhibit pharmacological activities such as antioxidant, the leaves of artichoke have important drug values. In present experiment, the effect of salt stress on phenolic compounds and antioxidant activity of artichoke leaves grown up in the field and pot conditions were studied. The salinity of the soil varied from 1.5 to 20.6 dS m-1 and 1.3 to 29 dS m-1 in pot and field experiments, respectively. Results showed that the highest amount of these compounds was recorded at 6.9 and 6.45 dS m-1 in pot and field experiments, respectively. As salinity increased up to 6 in both experiment, a reduction in total phenol and flavonoid was observed. Results showed that the antioxidant activity of leaf, positively increased by the salinity levels; in both trials the maximum antioxidant activity (IC50 = 0.01 mg mL-1) was observed at highest level of salinity. Similar to phenolic and flavonoid compounds the contents of chlorogenic and caffeic acids were at their highest levels in moderate salinity. Caffeic and chlorogenic acids accumulation was limited by high level of salinity without reduction in antioxidant activity. It can be concluded that at under the moderate and high salinity conditions, health-promoting compounds (polyphenol contents) in artichoke leaves are enhanced.
PDF Abstract XML References Citation
Received: October 27, 2011;
Accepted: November 23, 2011;
Published: January 10, 2012
How to cite this article
Amir Rezazadeh, Azim Ghasemnezhad, Mojtaba Barani and Telmah Telmadarrehei, 2012. Effect of Salinity on Phenolic Composition and Antioxidant Activity of Artichoke (Cynara scolymus L.) Leaves. Research Journal of Medicinal Plants, 6: 245-252.
URL: https://scialert.net/abstract/?doi=rjmp.2012.245.252
URL: https://scialert.net/abstract/?doi=rjmp.2012.245.252
INTRODUCTION
Artichoke (Cynara scolymus L.) is an herbaceous perennial plant belonging to the Compositae family (Asteraceae) that grows naturally in severe habitat conditions with high temperature, high salinity and drought in summer (Benlloch-Gonzalez et al., 2005). The heads of the artichoke are edible and the leaves have many pharmaceutical properties (Gebhardt, 1997). The action of phenolic compounds as antioxidant activity is mainly due to caffeoylquinic acid derivatives such as Chlorogenic acid and flavonoids such as luteolin glycosides effects (Gebhardt and Fausel, 1997; Brown and Evans, 1998).
In hard climatic conditions, plants subjected to stress, produce Reactive Oxygen Species (ROS) that cause cellular damage and are known to be involved in several plant disorders (Abdi and Ali, 1999). It is believed that increase in secondary metabolites synthesis in response to stressful conditions protect the cellular structures against oxidation (Chanwitheesuk et al., 2005). It has been proven that, in the hard environmental conditions the amount of antioxidant and antimicrobial components are increased in the plant tissues (Maisuthisakul et al., 2007). In plant, environmental stresses (biotic and abiotic) such as salinity lead to accumulation of polyphenol constituents (Dixon and Palva, 1995). Many studies confirmed the effect of salinity on antioxidant accumulation of the plant tissues. For example the polyphenols content and antioxidant activity of leaves of the halophyte Cakile maritime were increased by salinity (Ksouri et al., 2007). Leaf phenolic content of Artichoke was significantly increased at 25-50 mM NaCl (Hanen et al., 2008). Parida et al. (2004) reported the accumulation of polyphenols in moderate salinity in mangrove. In paper fruits, the total phenolic content increased with salinity level in red matured fruits (Navarro et al., 2006). Despite the induction of polyphenol contents in the tissues, stresses restrict biomass production of plant (De Abreu and Mazzafera, 2005). Antioxidants play an important role in the defense mechanism and hence, augment in response to these stresses (Dixon and Palva, 1995). In fact, plants with high levels of antioxidants, either organic or induced, have higher resistance to this oxidative damage (Dhindsa and Matowe, 1981).
Golestan province in North of Iran has especial environmental conditions from highly wet acidic soil forest to saline desert. The salinity varies from 1 to more than 50 dS m-1 (Siadat et al., 1997). The main aim of the present study was investigation the effect of natural salinity on the stability of artichoke plant (biomass production) and the quality of leaves (antioxidant compounds) to evaluate the potential of artichoke cultivation in this area. To compare the obtained results with control data, a pot experiment also was performed.
MATERIALS AND METHODS
Plant material and cultivation conditions
Field trial: This research was carried out at Golestan province, in the North of Iran during two successive seasons in 2009-2010. According to the soil salt level map of Golestan province (Iran) six sites with desirable range of salinity were selected and a complete randomized block design with two varieties and three replications was designed. Electrical Conductivity (EC) of the soil samples were evaluated in laboratory. For electrical conductivity more than 50 samples were collected and among them 6 locations with different EC (1.3, 3.5, 6.45, 11.5, 21 and 29 dS m-1) were selected as experimental sites. To check the EC variations, new soil samples were collected and the EC was calculated every 2 months. Seeds of artichoke (green globe and local variety) were sterilized, scarified and then planted into pot (of 8 cm diameter) and were kept in greenhouse. One month later the seedling was transplanted in the main place in the field. Irrigation was done when necessary. Four months after planting leaves were harvested and transferred to laboratory for chemical analysis.
Greenhouse trial: This research was carried out at Gorgan University of Agricultural Sciences and Natural Resources, Gorgan, Iran during two successive seasons in 2010-2011. A complete randomized design with 6 levels of salinity, two variety and 4 replications was performed in greenhouse trial. Seeds of two varieties of artichoke (green globe and local variety), sterilized and scarified in sulfuric acid (H2SO4) for 15 min. Then the seeds were planted into round plastic pots (of 35 cm diameter and 40 cm height) filled with mix of 50% sand and 50% clay (EC = 1.5 dS, m-1 pH = 7.6) and were kept in cold greenhouse (with daily temperature of 14-26°C) and irrigated with tap water (EC = 0.3 dS m-1). To perform the salinity, 35 days after planting, irrigation with saline water with different salinity (0, 50, 100, 150, 200 mM NaCl) was started and continued until harvest (120 days). To check the electrical conductivity of saturated soil extracts, soil was sampled every 2 month. Mean EC values were 1.5, 6.9, 11.5, 16.4 and 20.6 dS m-1.
Finally, In both experiments the parameters like total phenol and flavonoid compounds, antioxidant activity and the content of 5-O-cafeoyl quinic acid (chlorogenic acid) and Caffeic acid were measured.
Chemicals: Caffeic acid and chlorogenic acid were prepared from Roth (Karlsruhe, Germany); 1, 1-diphenyl-2-picryl hydrazyl radical (DPPH), Gallic acid, quercetin, acetic acid and acetonitril were purchased from Merck (Germany). Folin Ciocalteau reagent was obtained from Sigma Chemicals Co., USA. Dionised water was used. All other solvents were HPLC grade from Merck (Darmstadt, Germany).
Extract preparation: The leaf samples were collected from the experimental plants of both field and greenhouse trials. Well drained (at room temperature) samples were finely powdered and each sample (5 g) was extracted by percolation method using pure methanol (50 mL) for 24 h at room temperature. The obtained extract concentrated to a crude solid extract using a rotary vacuum evaporator at 40°C and then freeze-dried to have a complete solvent removal extract.
Determination of total phenol: Total phenol content was determined by Folin Ciocalteau reagent (McDonald et al., 2001). The extract of samples (0.5 mL of different dilutions) was mixed with Folin Ciocalteau reagent (5 mL, 1:10 diluted with distilled water) and aqueous Na2CO3 (4 mL, 1 M). The mixture was allowed to stand for 15 min and the phenols were determined by colorimetry at 765 nm. Total phenol value was expressed as gallic acid equivalents per gram (mg GAE g-1 DW). All samples were analyzed in three replications. The standard curve was prepared by 0, 50, 100, 150, 200 and 250 mg mL-1 solutions of Gallic acid in methanol: water (50:50, v/v) (R2 = 0.99).
Determination of total flavonoid: Colorimetric aluminum chloride method was used for flavonoid determination (Chang et al., 2002). Briefly, 0.5 mL solution of each plant extracts in methanol were separately mixed with 1.5 mL of methanol, 0.1 mL of 10% aluminum chloride, 0.1 mL of 1 M potassium acetate and 2 mL of distilled water and left at room temperature for 30 min; the absorbance of the reaction mixture was measured at 415 nm with Camspec M501 Single Beam Scanning UV/Vis Spectrophotometer. Total flavonoid contents were calculated as quercetin from a calibration curve. The calibration curve was prepared by preparing quercetin solutions at concentrations of 12.5 to 100 mg mL-1 in methanol.
DPPH radical scavenging assay: DPPH was used for determination of free radical-scavenging activity of the extracts (Koleva et al., 2002). Different concentrations of each leaf extract were added at an equal volume to methanolic solution of DPPH (100 μM). The mixture was shaken vigorously and left standing. After 15 min at room temperature, the absorbance was recorded at 517 nm as a lower IC50 value corresponds to a higher antioxidant activity. The experiment was repeated for three times. IC50 values denote the concentration of sample which is required to scavenge 50% of DPPH free radicals.
Determination of chlorogenic and caffeic acids by HPLC: The contents of chlorogenic and caffeic acid were determined using HPLC method. Standards were dissolved in methanol, the extract solutions was filtered through 0.25 μm filter (Whatmen type). The HPLC analysis was carried out on a Merck Hitachi apparatus model LaChrom L-7100 connected to a computer analytical program HSM and an RP C18 μ bondapak column (250 x 4.6 mm, 5 μm) was set at 40°C. Mobile phase included filtered H3PO4 (Phosphoric acid 0.5%), acetonitril and dionised water. The flow rate was kept at 1 mL per min. UV detector at 280 nm was used for detection. Identification of individual polyphenols was carried out using their retention times (chlorogenic acid: 9 min; caffeic acid: 14 min).
Statistical analysis: In present experiment a randomized complete block design with three replicates per treatment was used. The statistical analysis was done using the SPSS software version 17.0. Least Significant Differences (LSD) at α = 5% were computed for measurements with F-value <0.05.
RESULTS AND DISCUSSION
Total phenol and flavonoid contents: The leaf phenolic content was dependent on soil salinity. The accumulation of these compounds was significantly increased after treatment with moderate salinity both in field and pot trial. In the field trial, total phenol concentration was augmented from 50.1 to 70.2 mg GAE g-1 in 1.3 and 6.45 dS m-1, respectively. In contrast, at higher salinity, the accumulation of these compounds declined (Table 1). In greenhouse, by increasing of salt concentration from 1.5 up to 6.9 dS m-1, phenolic contents were significantly increased by 20% (60.3 to 72.3 mg GAE g-1). However, at higher salinity polyphenol concentration was diminished (Table 2). As a results the amount of polyphenols in leaf were maximal at 6.45 and 6.9 dS m-1, respectively in field and greenhouse trials. An increase in phenolic compounds in artichoke leaf with moderate salinity has been reported by Hanen et al. (2008). Also increase in phenolic compounds response to salinity has also been reported in extract of different tissues of some other plants (Agastian et al., 2000; Muthukumarasamy et al., 2000). In contrast, the total phenolic contents of 5- and 7-day-old radish sprouts were significantly diminished by moderate salinity compare with the control (Yuan et al., 2010). It is supporting the theory that leaf polyphenols protect artichoke against the oxidative stress generated by salinity and secondary metabolites may play a role in artichoke salinity tolerance.
Salinity had a same effect on flavonoid accumulation as phenolic compounds. Total flavonoid contents in salt-stressed artichoke plants were significantly increased. Flavonoid content ranged from 19.6 to 49.7 mg quercetin equivalent per g of extract powder (Table 1 and 2). As salinity increased from low to moderate, an increase in the content of flavonoid was observed in both experiments; 40 and 20% in field and pot experiments, respectively. In contrast to, at higher salinity levels a significant reduction of flavonoid was observed.
Salt stress restricts plant growth more than photosynthesis; as a result, plant diverts the synthesis of carbohydrates to produce secondary metabolites (Coley, 1986). In contrast, at high salt concentration, uptake of phosphor and potassium that are principal substance of secondary metabolites such as polyphenols, decline (Waring and Pitman, 1985). Also, due to disturbance of enzymatic activities in high salinity, photosynthesis is declined; therefore, growth and production of polyphenols will be decreased (Wong et al., 2006). It can explain the observed reduction of polyphenol compounds at high salt concentration in our experiments.
| Table 1: | Effect of salinity on the polyphenol accumulation of Cynara scolymus leaves in field experiment |
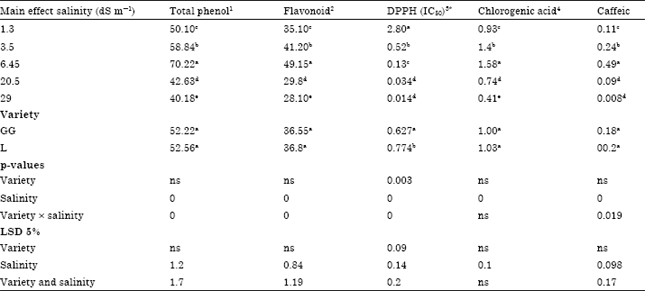 | |
| 1 mg gallic acid equivalent per g of extract powder, 2 mg quercetin equivalent per g of extract powder, 3mg mL-1, 4mg g-1 of extract powder, lower IC50 means higher antioxidant activity, GG: Green Globe, L: Local, Means in the same column followed by different letters differ significantly at p<0.05 | |
| Table 2: | Effect of salt treatments on polyphenol compounds of Cynara scolymus in pot experiment |
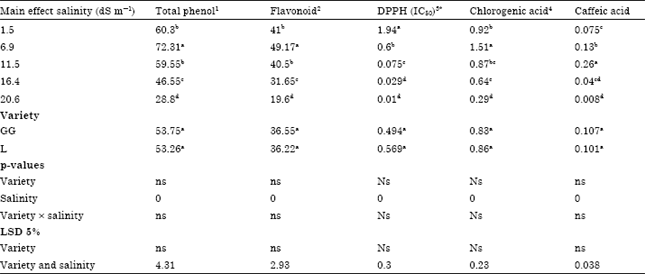 | |
| 1 mg gallic acid equivalent per g of extract powder, 2 mg quercetin equivalent per g of extract powder, 3mg mL-1, 4mg g-1 of extract powder, lower IC50 means higher antioxidant activity, GG: Green Globe, L: Local, Means in the same column followed by different letters differ significantly at p<0.05 | |
Other factors such as interactions among different molecules, water solubility and etc could explain the lack of correlation between potency and amount of polyphenols.
It can be concluded that moderate salinity induced the normal saline tolerance pathway via increasing total phenolic and flavonoid compounds. As salinity increased up to normal tolerance of plant, system is substituted by other mechanism as obviously can be seen in antioxidant activity of extract.
DPPH radical-scavenging activity: In present experiment antioxidant activity of leaf extract was dependent on the salt concentration in the medium. It has been found that the radical scavenging activity of the extract enhanced over the whole range of salt concentration (Table 1, 2). When the interaction effect of used treatments was investigated, a significant difference was observed (Table 1, 2). In this respect, the highest value was obtained in local variety in pot experiment at 200 mM NaCl (20.6 dS m-1). The antioxidant activity was maximal (IC50 = 0.014 and IC50 = 0.01 mg mL-1 in field and pot experiments, respectively) at the highest salinity level. It has been reported a close link between the antioxidant capacity and salinity tolerance in several crops, such as pea (Hernandez et al., 2000), cotton (Gosset et al., 1994), rice (Dionisio-Sese and Tobita, 1998) and foxtail millet (Sreenivasulu et al., 2000). No significant correlation was observed between antioxidant activity and total phenolic content (data did not showed). Phenolic contents increased in moderate salinity (6.45 and 6.9 dS m-1) and declined at high salt levels. Opposite to the reduction of phenolic and flavonoid compounds at high salinity, there was a dramatic increase in antioxidant activity related to higher salinity level both in field and greenhouse trial (Table 1, 2). Djeridane et al. (2006) explained the unclear relationship between the antioxidant activity and total phenol in several ways: the total phenolic fraction does not incorporate all the antioxidants, and synergistic interactions between the antioxidants in the mixture make the antioxidant activity not only dependent on the concentration but also on the structure and the nature of the antioxidants. In addition, the overall antioxidant effect is a complex process that depends on several modes of polyphenol interaction which may participate to differing degrees due to their deferent ratios among subclasses or single molecules.
Based on the obtained results of present study, it can be concluded that there is a wide variation of antioxidant compounds in artichoke under different levels of salinity and/or the function of compounds change under different levels of stress (e.g., salt stress).
Chlorogenic acid and caffeic acid: Both in field and pot experiments, a significant increase was observed in the chlorogenic acid content at the moderate level of salinity (1.58 and 1.51 mg g-1 DW at 6.45 and 6.9 dS m-1 field and pot experiments, respectively). Similar variation was observed in caffeic acid accumulation (Table 1, 2). In field experiment as salinity increased from 1.3 to 6.45 dS m-1, the content of caffeic acid reached to a maximum amount of 0.49 mg g-1 DW. In contrast, in pot experiment the observed increase in caffeic acid content was limited at the salinity of 11.5 dS m-1. Up to the moderate level of salinity both in pot and field experiment, a reduction was observed in the content of these two phenolic compounds. Therefore, significant increase in the accumulation of caffeic acid and chlorogenic acid in response to salinity suggesting that these phenolic compounds are perhaps stress-induced in Artichoke. This could help to reduce oxidative pressure, since caffeic acid shows high radical scavenging activity and was reported to increase both under drought (Grace, 2005) and high irradiance (Bidel et al., 2007) stresses. This observation supports the hypothesis that, due to their polyhydroxyl nature, chlorogenic acid and caffeic acid contributes significantly to the antioxidant activity of the artichoke leaves. As same conclusion was reported by Sgherri et al. (2007).
CONCLUSION
In conclusions, the results demonstrate that phenolic compounds constitute a major part of antioxidants in artichoke. Furthermore, environmental stresses could significantly enhance the antioxidant capacity of leave extract. As a result, salt stressed plants may be interesting potential sources of polyphenols for economical use.
In summary, although C. scolymus can be easily be propagated under high salinity condition (as was alive under 29 dS m-1 under field conditions) but the highest antioxidant activity as well as the maximum biomass production could be achieved under moderate salinity levels.
REFERENCES
- Abdi, S. and A. Ali, 1999. Role of ROS modified human DNA in the path-ogenesis and etiology of cancer. Cancer Lett., 142: 1-9.
CrossRefDirect Link - Agastian, P., S.J. Kingsley and M. Vivekanandan, 2000. Effect of salinity on photosynthesis and biochemical characteristics in mulberry genotypes. Photosynthetica, 38: 287-290.
CrossRefDirect Link - Benlloch-Gonzalez, M., J.M. Fournier, J. Ramos and M. Benlloch, 2005. Strategies underlying salt tolerance in halophytes are present in Cynara cardunculus. Plant Sci., 168: 653-659.
Direct Link - Bidel, L.P.R., S. Meyer, Y. Goulas, Y. Cadot and Z.G. Cerovic, 2007. Responses of epidermal phenolic compounds to light acclimation: In vivo qualitative and quantitative assessment using chlorophyll. Fluorescence excitation spectra in leaves of three woody species. J. Photochem. Photobiol., 88: 163-179.
CrossRef - Brown, J.E. and C.A. Rice-Evans, 1998. Luteolin rich artichoke extract protects low density lipoprotein from oxidation in vitro. Free Radic. Res., 29: 247-255.
CrossRef - Chang, C.C., M.H. Yang, H.M. Wen and J.C. Chern, 2002. Estimation of total flavonoid content in propolis by two complementary colometric methods. J. Food Drug Anal., Vol. 10.
CrossRefDirect Link - Chanwitheesuk, A., A. Teerawutgulrag and N. Rakariyatham, 2005. Screening of antioxidant activity and antioxidant compounds of some edible plants of Thailand. Food Chem., 92: 491-497.
CrossRefDirect Link - Coley, P.D., 1986. Costs and benefits of defense by tannins in a neotropical tree. Oecologia, 70: 238-241.
CrossRefDirect Link - De Abreu, I.N. and P. Mazzafera, 2005. Effect of water and temperature stress on the content of active constituents of Hypericum brasiliense Choisy. Plant Physiol. Biochem., 43: 241-248.
CrossRefDirect Link - Dionisio-Sese, M.L. and S. Tobita, 1998. Antioxidant responses of rice seedlings to salinity stress. Plant Sci., 135: 1-9.
CrossRefDirect Link - Dixon, R.A. and N.L. Paiva, 1995. Stress induced phenylpropanoid metabolism. Plant Cell, 7: 1085-1097.
PubMed - Djeridane, A., M. Yousfi, B. Nadjemi, D. Boutassouna, P. Stocker and N. Vidal, 2006. Antioxidant activity of some Algerian medicinal plants extracts containing phenolic compounds. Food Chem., 97: 654-660.
CrossRefDirect Link - Gebhardt, R., 1997. Antioxidative and protective properties of extracts from leaves of the artichoke (Cynara scolymus L.) against hydroperoxide-induced oxidative stress in cultured rat hepatocytes. Toxicol. Applied Pharmacol., 144: 279-286.
CrossRefPubMedDirect Link - Gebhardt, R. and M. Fausel, 1997. Antioxidant and hepatoprotective effects of artichoke extracts and constituents in cultured rat hepatocytes. Toxicol. In Vitro., 11: 669-672.
CrossRef - Gosset, D.R., E.P. Millhollon and M.C. Lucas, 1994. Antioxidant response to NaCl stress in salt-tolerant and salt-sensitive cultivars of cotton. Crop Sci., 34: 706-714.
CrossRefDirect Link - Hanen, F., R. Ksouri, W. Megdiche, N. Trabelsi, M. Boulaaba and C. Abdelly, 2008. Effect of Salinity on Growth, Leaf Phenolic Content and Antioxidant Scavenging Activity in Cynara cardunculus L. In: Biosaline Agriculture and High Salinity Tolerance, Abdelli, C., M. Ozturk, M. Ashraf and Y.C. Grignon, (Eds.). Birkhauser Verlag, Switzerland, pp: 335-343.
- Hernandez, J.A., A. Jimenez, P. Mullineaux and F. Sevilla, 2000. Tolerance of (Pisum sativum) to long term salt stress is associated with induction of antioxidant defenses. Plant Cell Environ., 23: 853-862.
Direct Link - Koleva, I.I., T.A. van Beek, J.P.H. Linssen, A. de Groot and L.N. Evstatieva, 2002. Screening of plant extracts for antioxidant activity: A comparative study on three testing methods. Phytochem. Anal., 13: 8-17.
CrossRefDirect Link - Ksouri, R., W. Megdiche, A. Debez, H. Falleh, C. Grignon and C. Abdelly, 2007. Salinity effects on polyphenol content and antioxidant activities in leaves of the halophyte Cakile maritima. Plant Physiol. Biochem., 45: 244-249.
CrossRefDirect Link - Maisuthisakul, P., M. Suttajit and R. Pongsawatmanit, 2007. Assessment of phenolic content and free radical-scavenging capacity of some thai indigenous plants. Food Chem., 100: 1409-1418.
CrossRefDirect Link - McDonald, S., P.D. Prenzler, M. Antolovich and K. Robards, 2001. Phenolic content and antioxidant activity of olive extracts. Food Chem., 73: 73-84.
CrossRefDirect Link - Muthukumarasamy, M., S.D. Gupta and R. Pannerselvam, 2000. Enhancement of peroxidase, polyphenol oxidase and superoxide dismutase activities by triadimefon in NaCl stressed Raphanus sativus L. Biol. Plant., 43: 317-320.
Direct Link - Navarro, J.M., P. Flores, C. Garrido and V. Martinez, 2006. Changes in the contents of antioxidant compounds in pepper fruits at different ripening stages, as affected by salinity. Food Chem., 96: 66-73.
CrossRefDirect Link - Parida, A.K., A.B. Das, Y. Sanada and P. Mohanty, 2004. Effects of salinity on biochemical components of the mangrove, Aegiceras corniculatum. Aqua. Bot., 80: 77-87.
CrossRefDirect Link - Sreenivasulu, N., R. Grimm, U. Wobus and W. Weschke, 2000. Differential response of antioxidant compounds to salinity stress in salt-tolerant and salt-sensitive seedlings of foxtail millet (Setaria italica). Physiol. Plant., 109: 435-442.
CrossRefDirect Link - Yuan, G., X. Wang, R. Guo and Q. Wang, 2010. Effect of salt stress on phenolic compounds, glucosinolates, myrosinase and antioxidant activity in radish sprouts. Food Chem., 121: 1014-1019.
CrossRef - Waring, R.H. and G.B. Pitman, 1985. Modifying lodgepole pine stands to change susceptibility to mountain pine beetle attack. Ecology, 66: 889-897.
Direct Link - Wong, C.C., H.B. Li, K.W. Cheng and F. Chen, 2006. A systematic survey of antioxidant activity of 30 Chinese medicinal plants using the ferric reducing antioxidant power assay. Food Chem., 97: 705-711.
CrossRefDirect Link