
K. Gill
Department of Biophysics, All India Institute of Medical Sciences, New Delhi, India
A.K. Singh
Department of Biophysics, All India Institute of Medical Sciences, New Delhi, India
S. Kumar
Department of Biophysics, All India Institute of Medical Sciences, New Delhi, India
B. Mishra
Department of Biophysics, All India Institute of Medical Sciences, New Delhi, India
V. Kapoor
Department of Biotechnology, All India Institute of Medical Sciences, New Delhi, India
S.N. Das
Department of Biotechnology, All India Institute of Medical Sciences, New Delhi, India
R.K. Somvanshi
Department of Biophysics, All India Institute of Medical Sciences, New Delhi, India
S. Dey
Department of Biophysics, All India Institute of Medical Sciences, New Delhi, India
Research Journal of Medicinal Plants
Year: 2012 | Volume: 6 | Issue: 2 | Page No.: 160-170







ABSTRACT
Ginger has been used in traditional Indian and Chinese medicine and is effective for a wide range of ailments including diarrhea, respiratory disorders, inflammatory diseases, arthritis etc. Recent studies have shown the role of ginger extract in the modulation of biochemical pathways involved in chronic inflammation and have thus provided evidences for its anti-inflammatory property. The aim of the study was to identify and purify a novel protein from ginger rhizomes (Zingiber officinales), of Zingiberaceae family possessing anti-fungal, anti-inflammatory and anti-proliferative properties as elicited using antibiotic diffusion method, surface plasmon resonance, spectrophotometric analysis and flow cytometry, respectively. The purified protein G-24 having molecular mass of 24 kD exhibited a potent anti-fungal activity against the mycelial growth in Fusarium exysporum and Candida albicans. It had shown 60% inhibition of human oral cancer cell line (KB cells) at 10 μM concentration. It inhibited inflammatory enzymes; lipooxygenase (LOX) and cyclooxygenase-2 (COX-2) with KD values of 2.04 μM and 2.74 nM, respectively. This confirmed the anti-inflammatory property of G-24.Thus, concluded that the G-24 protein possessed multiple functions viz. antifungal, anti-inflammation and antiproliferation.
PDF Abstract XML References Citation
Received: July 13, 2011;
Accepted: September 27, 2011;
Published: November 02, 2011
How to cite this article
K. Gill, A.K. Singh, S. Kumar, B. Mishra, V. Kapoor, S.N. Das, R.K. Somvanshi and S. Dey, 2012. Isolation and Characterization of a Potent Protein from Ginger Rhizomes Having Multiple Medicinal Properties. Research Journal of Medicinal Plants, 6: 160-170.
URL: https://scialert.net/abstract/?doi=rjmp.2012.160.170
URL: https://scialert.net/abstract/?doi=rjmp.2012.160.170
INTRODUCTION
Plants have an ample of defense mechanisms that are induced on pathogenic infection in order to protect themselves (Ghosh and Ulaganathan, 2004). In case of fungal infections, these mechanisms include the synthesis of many low molecular mass inhibitory compounds such as phenols, tannins, melanins or phytoalexins. The accumulation of proteins capable of acting directly on the fungi also inhibits its growth. Such proteins that help in plant protection are known as Pathogenesis-related (PR) proteins (Huynh et al., 1996).
The PR proteins are divided into several divisions on the basis of their structure, serological relatedness, enzymatic reactions and biological activities (Selitrennikoff, 2001; Edreva, 2005). Earlier, Wang and Ng (2005) had isolated an antifungal protein of molecular weight 32 kD from ginger.
The natural bioactive agents in plants are of continual inspiration for antimicrobial agents and clinical use due to low toxicity, no chemical modification, broad and good pharmacokinetics (Nenaah and Ahmed, 2011). The Ginger inhibits the enzymes like cyclooxygenase-1 and cyclooxygenase-2 in a manner that repress the synthesis of prostaglandins (Kiuchi et al., 1982; Ali et al., 2008). Besides, it was also seen that the ginger inhibits 5-lipoxygenase thereby repressing leukotriene biosynthesis. Thus, these properties set ginger apart from the anti-inflammatory steroidal drugs like NSAIDs (Non Steroidal Anti Inflammatory Drugs). Moreover, it was also seen that the compounds having the dual potential to inhibit both cyclooxygenase and lipoxygenase are better therapeutic agents than NSAIDs as have fewer side effects (Goldstein, 2004).
This study reports the identification of a novel 24 kD antifungal protein (G-24) from ginger having an anti-inflammatory property against the inflammatory enzymes; lipoxygenase and cyclooxygenase-2. It also exhibits anti cancerous property.
MATERIALS AND METHODS
Plant material: This study was conducted in 2009. The ginger rhizomes (1 kg) were purchased from a local market and were further processed at the All India Institute of Medical sciences, New Delhi, India for analysis.
Preparation of plant extract and purification of G-24: The purification involved an anion exchange chromatography using DEAE (Diethylaminoethyl) cellulose followed by an affinity chromatography. The ginger rhizomes were thoroughly washed and homogenized in distilled water. The crude extract was centrifuged at 10,000xg for 30 min. The resultant supernatant in 10 mM Tris-HCl buffer (pH 6.7) was loaded onto DEAE-cellulose column (2.5x50 cm) (Sigma Aldrich, USA) pre-equilibrated with 10 mM Tris-HCl buffer (pH 6.5). The elution was carried out with 10 mM Tris-HCl buffer containing a concentration gradient of 0.0-0.8 M NaCl (pH 6.5). The anti-fungal activity was checked in the unbound as well as in all the eluted fractions and was observed only in the unbound fraction. Hence, the unbound fraction was loaded onto an Affi-gel blue gel column (GE Healthcare Biosciences, Uppasala, Sweden) equilibrated with 10 mM Tris-HCl buffer (pH 6.5). The elution was carried out using a concentration gradient formed with 10 mM Tris-HCl buffer containing 0.0-0.8 M NaCl (pH 6.5).
Molecular mass determination: The sodium dodecyl sulphate-polyacrylamide gel electrophoresis (SDS-PAGE) was carried out using Laemmli system of buffers. The protein spot was excised from the gel and then subjected to in-gel reduction, alkylation and trypsin digestion. The mass spectrometry/mass spectrometry (MS/MS) (Bruker Daltonics, USA) was used to determine the mass and the sequence of the isolated protein. Each of the peptide formed after digestion was used for basic local alignment search tool (BLAST) search. It confirmed that the protein identified by Mascot search programme was the only relevant match in the non-redundant protein database for a particular peptide sequence. The statistically significant hits were recorded together with the number of peptides and percentage coverage of the protein (Kinter and Sherman, 2000).
N-terminal sequence analysis: The N-terminal sequence analysis of the purified protein was done by Edman degradation on a Procise Protein Sequencer (Applied Biosystems). The database was searched for other antifungal proteins with similar sequences using BLAST (http://www.ncbi.nlm.nih.gov/BLAST).
Assay for antifungal activity: The assay for antifungal activity toward Candida albicans (ATCC SC 5314) and Fusarium exosporium (Clinical isolates) was carried out in 90x10 mm Petri plates containing 20 mL of potato dextrose agar. These fungal species have been shown to be sensitive to a variety of antifungal proteins. After the mycelial colony had developed, wells were punctured at the three corners of the plates. An aliquot (100 μL of 0.12 μg) of G-24 protein solution was added to the wells. The positive and negative controls were 2.50 μg of Amphotericin B and the assay buffer without protein, respectively. The plates were incubated at 37°C for 72 h until mycelial growth had enveloped the wells, containing the assay buffer and the zones of inhibition formed around the wells containing the control and G-24 protein. To determine the IC50 value of the G-24 protein, four doses (0.1, 1.0, 2.0 and 20 μM) of it were added separately to three aliquots each containing 4 mL of potato dextrose agar at 45°C, mixed rapidly and poured into three separate small Petri dishes. After the agar had cooled down, a small amount of mycelia of each fungal colony was added. Buffer without G-24 protein served as a control. After incubation at 37°C for 72 h, the IC50 was determined by the following formula (Wang and Ng, 2003).
The percentage of inhibition of fungal growth is calculated as:
Anti inflammatory assay
Inhibition Assay for LOX and COX-2: The activity of purified Lipoxygenase was determined by using the conjugated diene method (Somvanshi et al., 2008) of biochemical assay. The enzyme activity was measured on T60 UV- visible spectrophotometer (Wibtoft. Leics, United Kingdom) by estimating the rate of formation of Hydroperoxy lipid product at an absorbance of 234 nm. For the inhibition studies, 1:1 molar ratios of the soybean lipoxygenase (LOX) which was earlier purified from the lab was incubated with G-24 for 30 min and the decline in activity was determined. The percentage inhibition was calculated (Somvanshi et al., 2008).
In case of COX-2 activity, the peroxidase assay was performed to estimate the formation of prostaglandin at 610 nm. For the inhibition assay, the recombinant COX-2 was incubated with G-24 in 1:1 molar ratio in the assay buffer for 45 min at 25°C. The decline in activity was determine and the percentage inhibition of G-24 was then calculated (Somvanshi et al., 2007).
Surface plasmon resonance (SPR) studies-binding studies of G-24 with LOX and COX-2: The binding properties of G-24 were investigated by Surface Plasmon Resonance (SPR) (Nylander et al., 1982). All SPR measurements were performed at 25°C using the BIAcore-2000 (Pharmacia Biosensor AB, Uppsala, Sweden) which is a biosensor-based system for the real-time specific interaction analysis.
The sensor chip CM5, surfactant P20, the amine coupling containing N-hydroxysuccinimide (NHS), N-ethyl-N’-3 diethylaminopropyl carbodiimide (EDC) and ethanolamine hydrochloride (Pharmacia Biosensor AB, Uppsala, Sweden) were used. The LOX was immobilized on the sensor chip CM5 at a flow rate of 10 μL min-1 at 25°C using amine coupling method (Somvanshi et al., 2008). The CM5 sensor chip was immobilized with LOX protein (Somvanshi et al., 2008). The three different concentrations of G-24 i.e., 0.01, 0.03 and 0.05 μM were passed at a flow rate of 10 μL min-1 at 25°C over the immobilized LOX. The dissociation of protein was performed by 10 mM HBS-EP buffer (pH 7.4).
The COX-2 was immobilized over the Ni-NTA chip (Somvanshi et al., 2007). The interaction of COX-2 with G-24 was performed by passing three different concentrations of G-24 (0.01, 0.03 and 0.05 μM) over the immobilized COX-2 at a flow rate of 10 μL min-1 for 4 min and the change in RU was observed. The sensor chip with the immobilized COX-2 was already available in the laboratory. The rate constants KA and KD were obtained by fitting the primary sensogram data using the BIA evaluation 3.0 software (Myszka, 2000; Karlsson and Falt, 1997).
Protease and protease inhibitory activity assay: The assay was done using 50 μL each of the purified G-24 (test sample) and the trypsin solution (positive control). The test sample as well as the positive control was incubated with 350 μL of freshly prepared casein (Sigma Aldrich, USA) solution for 30 min followed by the addition of 1 mL of 4% (w/v) trichloroacetic acid. The resultant mixture was incubated at room temperature for 30 min and centrifuged at 10,000 rpm for 15 min. The absorbance of the casein fragments produced in the supernatant by the proteolytic action was observed at 280 nm against water as a blank.
For the protease inhibition assay, G-24 protein was incubated with trypsin in 50 mM Tris-HCl buffer (pH 8) at 25°C for 30 min and inhibition was then carried out using a similar protocol as for the protease activity assay for the enzyme. The absorbance of supernatant was recorded at 280 nm.
Anti-proliferative assay by flow cytometry: Human oral squamous cell carcinoma cell line (KB) was obtained from National Centre for Cell Sciences (Pune, India) and maintained in Minimal Essential Media (MEM) (Sigma Aldrich, USA) supplemented with 10% foetal bovine serum and antibiotics (streptomycin, penicillin and fungizone). The tumour cells were used in subsequent experiments. Briefly, KB cells were plated in 24-well plates (50,000 cells/well) in duplicates at 37°C in CO2 incubator and after 24 h the tumour cells were pulsed with optimum doses (200 μL) of G-24 as well as cisplatin (positive control). The untreated control wells received equal volume of Phosphate Buffered Saline (PBS) (pH 7.4). The cells were harvested after 48 h, labeled with 50 μL mL-1 of Propidium Iodide (PI) and about 10,000 events were acquired in flow cytometer (BD LSR II, Becton-Dickinson). The percentage of dead cells (PI-labeled) was determined using BD FACSDivaTM software.
RESULTS
Purification of protein: The G-24 protein was purified by two chromatographic steps. The unbound fraction from the first step of purification i.e., DEAE cellulose column gave a positive antifungal test (data not shown). This unbound fraction was then loaded onto affinity column and the three adsorbed fractions; peak 1, 2 and 3 were obtained. The antifungal activity was found in peak 1 fraction which was eluted with 0.1 M NaCl in Tris buffer (pH 6.5) and not in the peak 2 and 3 eluted with 0.6 M and 0.8 M NaCl in Tris buffer (pH 6.5) as shown in Fig. 1A. The Fig. 1B showed the molecular masses of proteins present in crude sample, unadsorbed fraction from DEAE-Cellulose and 0.1 M NaCl fraction of affinity gel column. The 0.1 M NaCl fraction from affinity gel column was also subjected to silver staining (Fig. 1C).
The protein concentrations obtained in unbound fraction from anion exchange and the final fraction from affinity gel chromatography eluted with 0.1 M NaCl were 47 and 10 mg mL-1, respectively, as quantified by Bradford method.
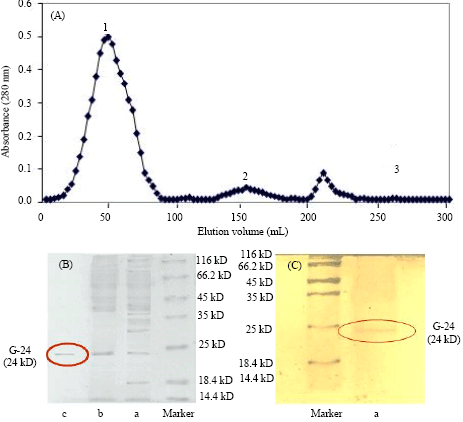 | |
| Fig. 1 (A-C): | (A) Affinity chromatography profile for affi-blue gel column. The unabsorbed fraction from the DEAE-cellulose column was loaded on to the affi-blue gel column pre-equilibrated with the 10 mM tris buffer (pH 6.5) and a concentration gradient of 0.1-0.8 M NaCI in 10 mM tris buffer (pH 6.5) was used for elution. The protein G-24 was eluted in the 0.1 M NaCI in 10 mM tris buffer (ph 6.5) fraction as shown in peak 1. (B) SDS-PAGE of G-24 using coomassie brilliant blue stain: (A) crude extract, (B) unbound fraction from DEAE-Cellulose, anion exchange, (C) 0.1 M NaCI eluted fraction of affi-gel blue gel, affinity chromatography. (C) SDS-PAGE of G-24 using sliver stain: (A) Purified protein G-24 eluted in 0.1 M NaCI eluted fraction of affi-gel, affinity chromatography |
Molecular weight determination: The final fraction (peak 1 of Fig. 1A) from affinity gel showing a single band of MW 24 kD (Fig.1 B, C) reveals ≥95% purity of the protein. The mascot search program from matrix science shows the candidate protein as the purified protein and the score of 78 based on probability analysis.
N-terminal sequence: The N-terminal sequence of the purified antifungal ginger protein, G24 was GVLPDSVVTT as shown in Table 1 and exhibits some similarity with the partial sequence of an arietin (Chickpea antifungal peptide). It was different from N-terminal sequence of the protein first isolated from ginger rhizome (Wang and Ng, 2005).
Antifungal activity assay: The antifungal activity of the purified protein was observed towards Fusarium exosporium and Candida albicans (Fig. 2). The mycelium growth had advanced till it reached the peripheral discs containing the test sample and both the controls.
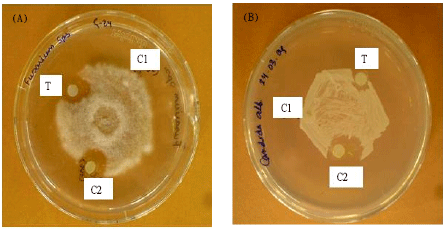 | |
| Fig. 2 (A-B): | Antifiungal activity of G-24 towards; (A) Fusarium exosporium and (B) Candida albicens. 100 μL (0.12 μg) G-24 in 10 mM Tris-HCI pH 6.5 buffer containing 0.1 M NaCI as test sample (T), 10 mM Tris-HCI pH 6.5 buffer containing 0.1 M NaCI (CI) was used as a negative control and μg mL-1 amphotericin B (C2) was used as a positive control |
| Table 1: | N-terminal sequence of ginger antifungal protein (results of BLAST search) |
 | |
The crescents of inhibition zone around the test sample containing G-24 and the positive control with Amphotericin B were observed, confirming the antifungal activity of G-24. The IC50 values for the antifungal activity against the two aforementioned fungal species were 4.6 and 8.0 μM, respectively.
Anti-inflammatory activity analysis by spectrophotometer: The spectrophotometric data obtained showed nearly 70% activity loss for LOX after incubation with G-24 in the presence of the substrate linoleic acid and 80% activity loss for COX-2 in the presence of the substrate arachidonic acid and the co-factor haematin revealing that G-24 exhibits strong anti-inflammatory property. Figure 3a had shown the rise of absorbance with time, i.e., the activity of LOX in presence of substrate and in Fig. 3b, the absorbance became stable much earlier with the increase in time in presence of G-24 protein. Similarly, in Fig. 3c the rise in absorbance for the activity of COX-2 was due to the oxidation of TMPD in presence of substrate and in Fig. 3d the rate of oxidation decreased in the presence of G-24. This result indicated the inhibition of inflammatory enzymes by G-24.
Binding studies of LOX and COX-2 by SPR analysis: The anti-inflammatory property was further confirmed by the binding studies of G-24 with LOX and the recombinant COX-2 protein. The sensogram (Fig. 3e and f) showed binding of G-24 at varying concentrations with LOX and COX-2, respectively. The change in RU with varying concentrations of G-24 indicated the change in mass on LOX and COX-2 immobilized on the chip with time and the dissociation constants were found be 2.04 μM and 2.74 nM, respectively.
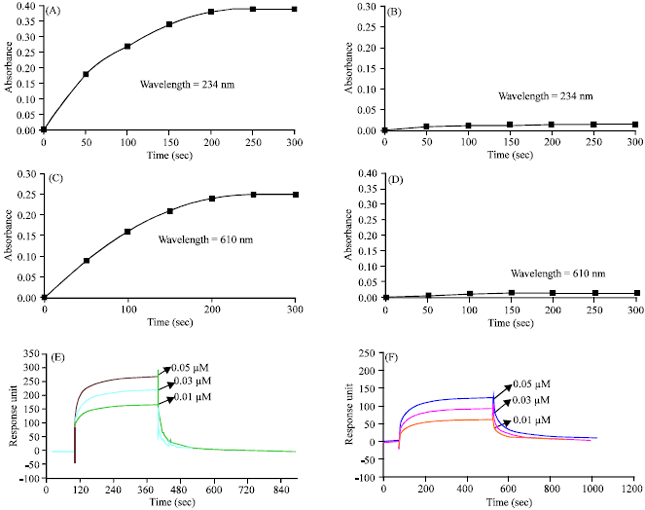 | |
| Fig. 3 (A-F): | Anti-inflammatory studies. (A) Activity profile of LOX, (B) Activity profile of LOX incubated with G-24 in 1:1 molar ratio for 45 min at room temperature, (C) Activity profile of COX-2, (D) Activity profile of COX-2 incubated with G-24 in 1:1 molar ratio for 45 min at room temperature. Sensogram showing binding of different concentrations of G-24 (I1 = 0.01 μM I2 = 0.03 μM I3 = 0.05 μM) on the immobilized (E) Lox and (F) COX-2 on the CM 5 sensor chip |
Protease and protease inhibitory activity assay: The purified G-24 protein was tested against trypsin as a positive control and was found that it did not exhibit protease and protease inhibitory activity (Data not shown).
Anti-proliferative activity assay: The present purified protein was also showing potent cytotoxic activity on human oral squamous carcinoma cell (KB) line. The cytotoxic assay was done by using PI labeling and flow cytometry. The G-24 induced cytotoxicity in KB cells in a dose dependent manner at 10-40 μM concentrations in 48 h culture. The frequency of cytotoxicity at 10, 20, 30 and 40 μM concentrations was 12, 41, 60.7 and 33%, respectively. Thus, the optimum cytotoxic dose of the protein was 30 μM (Fig. 4).
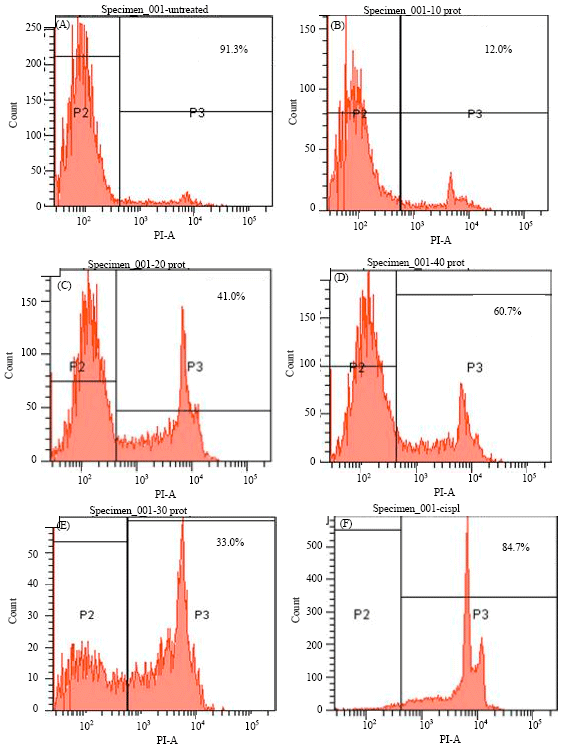 | |
| Fig. 4 (A-F): | Antitumor activity of G-24 on KB cells using PI staining and flow cytometry. (A) Untreated KB cells, (B) KB cells treated with 10 μM protein, (C) 20 μM protein, (D) 30 μM, (E) 40 μM and (F) cisplatin |
DISCUSSION
Many antifungal proteins from the plant sources exhibit anti-inflammatory and anti-proliferative properties. The ginger extract has shown to exhibit antithrombotic activities (Thomson et al., 2002; Srivastava and Mustafa, 1989) as it prevents the in vitro platelet aggregation and Thromboxane (TXB2) production (Thomson et al., 2002; Srivastava, 1984; Flynn et al., 1986). In addition, gingerdione has been shown to inhibit the formation of 5 Hydroxyeicosatetraenoic Acid (5-HETE) and Prostaglandin E2 (PGE2) from arachidonic acid. Gingerol and dehydroparadol favors the inhibition of COX-2 (Thomson et al., 2002; Flynn et al., 1986). Thus, the ginger extract used as a traditional medicine contains bioactive compounds possessing multiple properties viz. antimicrobial, anti-inflammatory and anticancerous (Abdul et al., 2008; Akram et al., 2011; Harliansyah et al., 2007; Khatun et al., 2003; Neogi et al., 2007; Patrick-Iwuanyanwu et al., 2007; Saeid et al., 2010; Tagoe et al., 2011). So far, only one antifungal protein was isolated from ginger by Wang and Ng (2005) and this study identified a protein (G-24) having multiple biological functions. This protein showed strong anti-fungal activity against the fungal species i.e., Fusarium exosporium and Candida albicans with IC50 values of 4.6 μ and 8.0 μM, respectively.
It was found that the G-24 exhibited anti-inflammatory activity by inhibiting LOX and the recombinant COX-2 with high inhibition and binding capacity.
Some protease inhibitors like Cysteine protease inhibitor from pearl millet exhibits antifungal activity (Joshi et al., 1998). Though the purified ginger protein G-24 showed evidence of antifungal activity, it is devoid of protease and protease inhibitory activity.
The presence of certain compounds like shogaols, zingerone and pungent vallinoids, viz. [6]-gingerol and [6]-paradol credit towards the antifungal property of ginger. It has also been reported that an ample of mechanisms are involved in the chemopreventive effects of ginger (Shukla and Singh, 2007). The present purified protein was also showing potent cytotoxic activity on human oral squamous carcinoma cell (KB) line.
CONCLUSION
Hence, it can be concluded that G-24 can be used as a potent medicinal plant product for clinical use as it exhibits various effective properties like antifungal, anti-inflammatory and anti-proliferative.
ACKNOWLEDGMENTS
Authors acknowledge School of Life Sciences, Jawaharlal Nehru University for performing Mass analysis of G-24 protein, National Institute of Immunology, New Delhi, India for N-terminal sequencing.
REFERENCES
- Abdul, A.B., S.I. Abdel-Wahab, A.S. Al-Zubairi, M.M. El-Hassan and S.M. Murali, 2008. Anticancer and antimicrobial activities of zerumbone from the rhizomes of Zingiber zerumbut. Int. J. Pharmacol., 4: 301-304.
CrossRefDirect Link - Akram, M., M.I. Shah, K. Usmanghan, E. Mohiuddin and A. Sami et al., 2011. Zingiber officinale roscoe (A medicinal plant). Pak. J. Nutr., 10: 399-400.
Direct Link - Ali, B.H., G. Blunden, M.O. Tanira and A. Nemmar, 2008. Some phytochemical, pharmacological and toxicological properties of ginger (Zingiber officinale Roscoe): A review of recent research. Food Chem. Toxicol., 46: 409-420.
CrossRefPubMedDirect Link - Edreva, A., 2005. Pathogenesis-related proteins: Research progress in the last 15 years. Gen. Applied Plant Physiol., 31: 105-124.
Direct Link - Goldstein, J.L., 2004. Challenges in managing NSAID-associated gastro-intestinal tract injury. Digestion, 69: 25-33.
PubMed - Huynh, Q.K., J.R. Borgmeyer, C.E. Smith, L.D. Bell and D.M. Shah, 1996. Isolation and characterization of a 30 kDa protein with antifungal activity from leaves of Engelmannia pinnatifida. Biochem. J., 316: 723-727.
PubMedDirect Link - Joshi, B.N., M.N. Sainani, K.B. Bastawade, V.S. Gupta and P.K. Ranjekar, 1998. Cysteine protease inhibitor from pearl millet: A new class of antifungal protein. Biochem. Biophys. Res. Commun., 246: 382-387.
PubMed - Karlsson, R. and A. Falt, 1997. Experimental design for kinetic analysis of protein-protein interactions with surface plasmon resonance biosensors. J. Immunol. Methods, 200: 121-133.
CrossRefPubMedDirect Link - Khatun, A., S. Nasrin and M.T. Hossain, 2003. Large scale multiplication of ginger (Zingiber Officinale Rosc.) from shoot-tip culture. J. Biological Sci., 3: 59-64.
CrossRefDirect Link - Kiuchi, F., M. Shibuya and U. Sankawa, 1982. Inhibitors of prostaglandin biosynthesis from ginger. Chem. Pharm. Bull., 30: 754-757.
PubMed - Myszka, D.G., 2000. Kinetic, equilibrium and thermodynamics analysis of macromolecular interaction with BIACORE. Methods Enzymol., 323: 325-340.
CrossRefPubMedDirect Link - Neogi, U., R. Saumya and B. Irum, 2007. In vitro combinational effect of bio-active plant extracts on common food borne pathogens. Res. J. Microbiol., 2: 500-503.
CrossRefDirect Link - Nenaah, E.G. and M.E. Ahmed, 2011. Antimicrobial activity of extracts and latex of Calotropis procera (Ait.) and synergistic effect with reference antimicrobials. Res. J. Med. Plant, 5: 706-716.
CrossRefDirect Link - Nylander, C., B. Leidberg and T. Lind, 1982. Gas detection by means of surface plasmon resonance. Sens. Actuators, 3: 79-88.
CrossRef - Patrick-Iwuanyanwu, K.C., M.O. Wegwu and E.O. Ayalogu, 2007. The protective nature of garlic, ginger and vitamin E on CCl4-induced hepatotoxicity in rats. Asian J. Biochem., 2: 409-414.
CrossRefDirect Link - Saeid, J.M., A.B. Mohamed and M.A. Al-Baddy, 2010. Effect of aqueous extract of ginger (Zingiber officinale) on blood biochemistry parameters of broiler. Int. J. Poult. Sci., 9: 944-947.
CrossRefDirect Link - Selitrennikoff, C.P., 2001. Antifungal proteins. Applied Environ. Microbiol., 67: 2883-2894.
CrossRefPubMedDirect Link - Shukla, Y. and M. Singh, 2007. Cancer preventive properties of ginger: A brief review. Food Chem. Toxicol., 45: 683-690.
CrossRefPubMedDirect Link - Somvanshi, R.K., A. Kumar, S. Kant, D. Gupta and S.B. Singh et al., 2007. Surface plasmon resonance studies and biochemical evaluation of a potent peptide inhibitor against cyclooxygenase-2 as an anti-inflammatory agent. Biochem. Biophys. Res. Commun., 361: 37-42.
CrossRef - Somvanshi, R.K., A.K. Singh, M. Saxena, B. Mishra and S. Dey, 2008. Development of novel peptide inhibitor of Lipoxygenase based on biochemical and BIAcore evidences. Biochim. Biophys. Acta, 1784: 1812-1817.
PubMed - Srivastava, K.C. and T. Mustafa, 1989. Spices: Antiplatelet activity and prostanoid metabolism. Prostaglandin, Leukot Essent. Fatty Acids, 38: 255-266.
PubMedDirect Link - Thomson, M., K. Al-Qattan, M. Al-Sawan, M.M. Alnaqeeb, I. Khan and M. Ali, 2002. The use of ginger (Zingiber officinale Rosc.) as a potential anti-inflammatory and antithrombotic agent. Prostaglandins Leukot Essent Fatty Acids, 67: 475-478.
PubMedDirect Link - Wang, H.X. and T.B. Ng, 2003. Purification of castamollin, a novel antifungal protein from Chinese chestnuts. Protein Expr. Purif., 32: 44-51.
PubMed - Wang, H. and T.B. Ng, 2005. An antifungal protein from ginger rhizomes. Biochem. Biophys. Res. Commun., 336: 100-104.
PubMed - Flynn, DL., M.F. Rafferty and A.M. Boctor, 1986. Inhibition of human neutrophil 5- lipoxygenase activity by gingerdione, shogaol, capsaicin and related pungent compounds. Prostaglandins. Leukot. Med., 24: 195-198.
PubMed - Tagoe, D.N.A., H.D. Nyarko and R. Akpaka, 2011. A comparison of the antifungal properties of onion (Allium cepa), ginger (Zingiber officinale) and garlic (Allium sativum) against Aspergillus flavus, Aspergillus niger and Cladosporium herbarum. Res. J. Med. Plant, 5: 281-287.
CrossRefDirect Link - Harliansyah, N.A. Murad, W.Z. Wan Ngah and Y.A.M. Yusof, 2007. Antiproliferative, antioxidant and apoptosis effects of Zingber officinal and 6-Gingerol on HepG2 cells. Asian J. Biochem., 2: 421-426.
CrossRef
Divyash Singh Reply
Sir I m also interested to pursue my research on GINGER, So Pz guide me what type of new and advanced research cd b done on GINGER. Sir I m Asst. proff. In PHARMACY COLLEGE. Thanks