
M. Pirkhezri
Department of Horticultural Sciences, Faculty of Agriculture, University of Tehran, Karaj 31587, Iran
M.E. Hassani
Department of Horticultural Sciences, Faculty of Agriculture, University of Tehran, Karaj 31587, Iran
J. Hadian
Department of Horticultural Sciences, Faculty of Agriculture, University of Tehran, Karaj 31587, Iran
Research Journal of Medicinal Plants
Year: 2010 | Volume: 4 | Issue: 1 | Page No.: 1-13







ABSTRACT
In present study, morphological traits and RAPD markers were used to analyze variation among different populations of M. chamomilla growing in four different provinces in Southwest of Iran along with some standard cultivars. Variance analysis of morphological traits showed that all evaluated traits were significantly different among populations. Maximum coefficients of variability were belonging to weight of plants (87.94%) and number of flower per plant (62.64%). Dendrogram was drawn based on Euclidean distances from morphological data matrix. All populations were represented into two main groups. With regards to yield of flowers, oil yield, plant height, etc., some populations showed performance equal to standard cultivars. In RAPD analysis 18 selected primers produced 220 bands, of which 205 were polymorphic. The ranges of similarity between populations were varied from 0.15 to 0.78. According to the UPGMA derived dendrogram, at a similarity level of 0.25, the populations were divided into two main groups. With regard to Nei's genetic diversity parameters calculated for different provinces of origins, maximum and minimum number of effective alleles was observed in Khuzestan (1.657) and Fars (1.142) provinces, respectively. The percentage of polymorphic loci (P loci% ), Nei's gene diversity (h) and Shannon's information index (I), calculated for different provinces, indicated that diversity was highest within Khuzestan province (P loci = 90.29%, h = 0.364 and I = 0.528) and lowest within Fars province (P loci = 20.87% , h = 0.16 and I = 0.23). Wide domain of genetic variability revealed in this study could be considered as a gene pool available for German chamomile improvement through selection and hybridization breeding programs. High heterosis effects will be expected for hybrids of these genetically diverse and talent populations.
PDF Abstract XML References Citation
How to cite this article
M. Pirkhezri, M.E. Hassani and J. Hadian, 2010. Genetic Diversity in Different Populations of Matricaria chamomilla L. Growing in Southwest of Iran, Based on Morphological and RAPD Markers. Research Journal of Medicinal Plants, 4: 1-13.
URL: https://scialert.net/abstract/?doi=rjmp.2010.1.13
URL: https://scialert.net/abstract/?doi=rjmp.2010.1.13
INTRODUCTION
German chamomile (Matricaria chamomilla L., syn. Chamomilla recutita, L. Rauschert) is an important and frequently cultivated medicinal plant belongs to Asteraceae family (Salamon, 1992; Newall et al., 1996). Wild chamomile is an annual herb originally from Europe which has dispersed and naturalized on almost every continent. The branched stem is somewhat erect, round, hollow and grows to about 20 inches tall. The leaves are bipinnate, finely divided, light green and feathery. The flowers are daisy-like and bloom from May to October (Gardiner, 1999). Chamomile flowers are used in alternative medicine as anodyne, anti-inflammatory, vulnerary, deodorant, bacteriostatic, antimicrobial, anticatarrhal, carminative, sedative, antiseptic and spasmolytic properties (Ferri and Capresi, 1979; Bruni, 1999). Several applications of dry powder for medicinal effect such as fevers, sore throats, the aches and pains due to cold, flu and allergies have also been reported by McKay and Blumberg (2006). About 120 chemical constituents have been identified in chamomile as secondary metabolites, including: sesquiterpenes [(-)-α- bisabolol, matricin or chamazulene], flavonoids (apigenin glycosides), polyacetylenes [(Z)-ene-nedicycloether], coumarins (herniarin and umbelliferone), mucilages, etc. (Schilcher, 1987; Mann and Staba, 1986).
The world market currently has chamomile drug of various origins and therapeutic values. With regard to increasing demands of drug and cosmetic industries, breeding German chamomile to gain talent cultivars with high yielding potential and homogeneity is necessary. Studies on genetic diversity of available germplasm have been conducted as first, essential and fundamental step of several breeding programs of German chamomile (Circella et al., 1993; Taviana, 2001; D'Andrea, 2002). Taviana (2001) studied genetic diversity of 13 accessions of M. chamomilla based on morphological and production biological traits mainly yield of flowers and oil content. D'Andrea (2002) investigated morphological and phytochemical variability among different chamomile cultivars from Italy. Jaymand and Rezai (2002) and Omidbaigi (1999) reported the essential oil content and chamazulene percentages of some Iranian accessions of M. chamomilla. Wagner et al. (2005) studied genetic diversity of M. chamomilla based on RAPD and AFLP markers. Variability of some Iranian and introduced cultivars of M. chamomilla based on morphological and RAPD markers has been reported by Solouki et al. (2008).
Different approaches in genetic diversity analyses reveal different level of polymorphism (Porter and Smith, 1982). Morphological traits are useful and practical markers in breeding program but they are influenced by environmental conditions while DNA markers are independence of environmental factors and show a greater level of polymorphism (Heywood, 2002). Therefore, they are considered as valuable tools for determining genetic relationships. Among various molecular markers, Random amplified polymorphic DNA (RAPD) markers have proved to be very useful tool providing a convenient and rapid assessment of the genetic differences between genotypes (Williams et al., 1990). RAPD analysis requires use of the Polymerase Chain Reaction (PCR) with 10-mer primers to generate random amplified fragments of DNA. The RAPD technique has several advantages such as a relatively unbiased portion of the genome sampled, simplicity of use, lower cost and the requirement of a very small amount of DNA for analyses (Williams et al., 1990; Fritzch and Rieseberg, 1996). Polymorphism detected by RAPD markers has proven to be useful for discrimination of genetic diversity and relationships in several medicinal plant species as in Ocimum basilicum L. (De Masi et al., 2006), Satureja hortensis (Hadian et al., 2008) and recently on Carthamus tinctorious (Khan et al., 2009). In the study presented here, combination of morphological, production biological and RAPD markers were used to analyze variation among different cultivars and populations of M. chamomilla growing in Southwest of Iran.
MATERIALS AND METHODS
This study was conducted during 2006-2008 at the Agricultural Faculty of Tehran University.
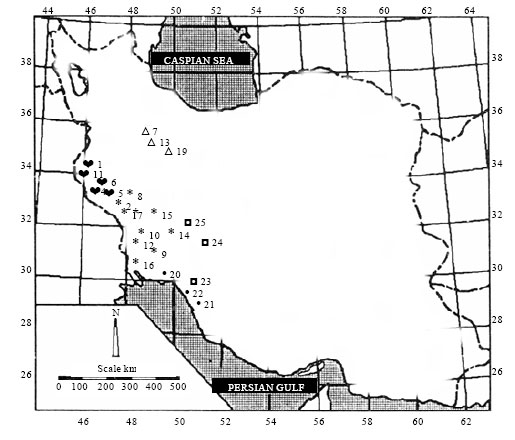 | |
| Fig. 1: | Distribution map of different accessions of Matricaria chamomilla in Lorestan ( |
Plant Material
Extensive effort was made to collect German chamomile from different geographical locations in South and South-West of Iran (Fig. 1), during April-May 2006. The sampling strategy involved tracing several sites in different parts of the investigated area in order to cover as much growing habitat as possible. Twenty one German chamomile populations from four provinces, Lorestan, Khuzestan, Booshehr and Fars and four cultivars from Germany, Hungary and Iran were used in this study. Number of samples taken in each province was depending on geographical distribution of German chamomile.
The seeds of all accessions were planted in propagation pots and then 45 plantlets of each accession transplanted to field with 60x30 cm spacing in a Randomized Complete Block Design (RCBD) with three replications. The soil was soft, well drained, with a pH value close to 7. The cultural operations consisted of manual elimination of weeds, frequent irrigation in order to maintain the soil wet and fertilizer administration. Plants were harvested individually at flowering stage and several morphological, production biological traits were evaluated. These traits were evaluated on the base of means of 30 individual plants (10 plants for each replication) for each accession. Essential oil yield of each accession was measured by hydro-distillation of 20 g shade dried flowers in three replications using a Clevenger apparatus.
DNA Extraction
Genomic DNA was extracted from freshly collected leaves according to the CTAB method of Murray and Thompson (1980). The purity and quantity of genomic DNA was determined spectrophotometrically and confirmed using 0.8% agarose gel (Roche Co., Germany) electrophoresis against known concentrations of Lambda DNA.
Amplification Reactions
RAPD analysis was conducted according to Williams et al. (1990). One hundred TIBM primers (TIBMOLBIOL Co., Germany) were used for PCR amplification. Polymerase chain reaction was performed in a total volume of 25 μL containing 10 ng template DNA, 1xPCR buffer, 1.75 mM MgCl2, 200 mM dNTPs (CinnaGen Co., Iran), 0.2 mM of a primer and 1 U Taq DNA polymerase (CinnaGen Co., Iran). Error was minimized by making one large batch (master mix) of all reagents (except template DNA) for each primer. PCR was performed in a thermocycler (iCycler, Bio Rad Co., USA) programmed as follows: 94°C/4 min, followed by 5 cycles of 92°C/1 min, 37°C/1 min, 72°C/1 min and followed by 35 cycles of 92°C/30 sec, 37°C/1 min, 72°C/45 sec and a final extension at 72°C for 7 min. Amplified products were separated by 1.5% agarose gel electrophoresis in TBE buffer. The PCR products were visualized by ethidium bromide staining and photographed under UV light, by a Gel Doc system (UVP, Bio Doc Co., USA).
Data Analysis
Quantitative analysis of morphological traits were carried out using the SAS system for Windows software, released 8.02 (SAS| Institute, Cary, NC, USA). Analysis of Variance (ANOVA) was performed and then the means of results were compared by Duncan’s multiple range tests. In order to determine the degree of associations among the characteristics, Pearson’s coefficients were used. The SPSS software was used to produce a distance matrix and a dendrogram based on morphological data. Average Euclidian distance was calculated for each variety-pair and the resulting distance matrix was used to construct a phenetic dendrogram among different accessions using Average Linkage (between groups) cluster analysis (Mohammadi and Prasanna, 2003).
Fractionated PCR products were analyzed by scoring the present (1) or absent (0) polymorphic bands in individual lane. The NTSYS-pc software ver. 2.02 (Rohlf, 1993) was used to estimate genetic similarities with the Jaccard’s (1908) coefficient. The matrix of generated similarities was analyzed by the Unweighted Pair Group Method with Arithmetic Average (UPGMA), using the SAHN clustering module. The cophenetic module was applied to compute a cophenetic value matrix using the UPGMA matrix. The MXCOMP module was then used to compute the cophenetic correlation, i.e., to test the goodness of fit of the cluster analysis to the similarity matrix.
POPGENE, Ver.1.31 (Yeh et al., 1997) software was used to describe structure of studied populations in different provinces and cultivars. To examine patterns of genetic diversity, genetic variation statistics for all loci (Nei, 1987) was performed. In addition, number of observed alleles (na), number of effective alleles (ne) (Kimura and Crow, 1963), Nei's gene diversity (h) (Nei, 1973) and Shannon's information index (I) (Lewontin, 1972) for each population was measured. Nei's analysis of gene diversity in subdivided populations (Nei, 1987) carried out with counting total heterozygosity (Ht), heterozygosity within populations (Hs), diversity coefficient among populations (Gst) and estimation of gene flow from Gst or Gcs (Nm) parameters. Fst index (Wright, 1951) was measured via this formula (Lynch and Milligan, 1994):
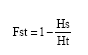 |
Genetic distances between Provinces were measured by Nei's original (Nei, 1972).
RESULTS
Morphological Analysis
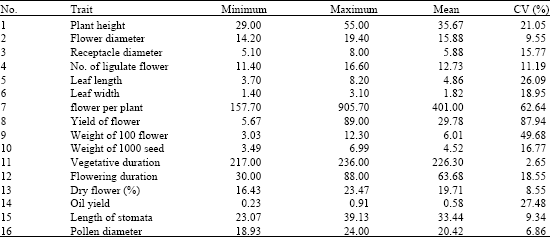
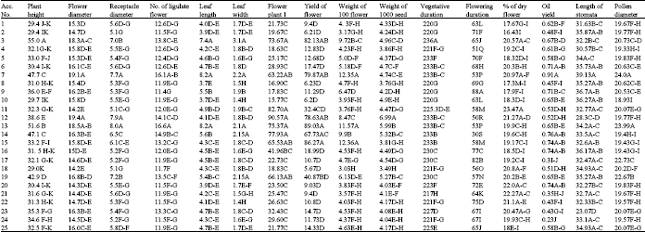
Several morphological and production biological characters were measured among populations and cultivars. Variance analysis showed that all evaluated traits were significantly different. Mean, min and max values of each trait are represented in Table 1. Maximum coefficients of variability were belonging to weight of plants (87.94%) and number of flower per plant (62.64%). Mean comparisons based on Duncan test are represented in Table 2. Results showed that with regard to height of plants, flower diameter and receptacle diameter and number of flower per plant, some local accessions were superior or equal to breaded cultivars. Correlation coefficients (Table 3) were significant for some traits. Significant positive correlation observed between yield and number of flower per plants (0.90), flower diameter and number of ligulate flowers (0.83), flower diameter and yield (0.82), height of plant and leaf length (0.82) and yield and number of ligulate flowers (0.81). Correlation coefficients were also significant but negatively for days of flowering and flower diameter (-0.53) and days of flowering and number of ligulate flowers (-0.50).
Dendrogram was drawn to display the phenetic relationships among different populations of German chamomile based on Euclidean distances from morphological data matrix. All populations were represented into two main groups (Fig. 2). In group A, 18 populations from Booshehr, Fars, Lorestan and Khuzestan provinces were represented. Group B comprised seven populations of which, three cultivars from Iran (No. 3), Hungary (No. 7) and Germany (No.13) were represented in the same subgroup (BI) while three populations from Khuzestan province (No. 12, 14 and 15) and Cv. Iran (No. 19) were represented in BII subgroup. Principal Component Analysis can explain the traits that are more differentiated among accessions. In PCA analysis of German chamomile accessions the first three principal components explained about 76.89% of total variation. First PC, explaining about 58.46% of variation, was linked to variables related to plant height, flower and receptacle diameter, number of ligulate flowers, width of leaves, number of flower per plants, yield of plant, weight of 100 flowers, weight of 1000 seeds, days to flowering and flowering duration. Second PC that was responsible for 11.83% of variations was linked to variables related to leaf length, oil yield and pollen diameter. Third PC, explaining only 6.6% of variation, was linked to yield of dry flowers.
| Table 1: | Mean, max, min and coefficient of variability (CV %) of different traits, studied among 25 populations of Matricaria chamomilla |
 | |
| Table 2: | Duncan test for mean comparisons of different traits among different accessions of Matricaria chamomilla |
 | |
Means with same or two letters are non significant, with different letters are significant | |
| Table 3: | Simple correlation between different traits among different accessions of Matricaria chamomilla |
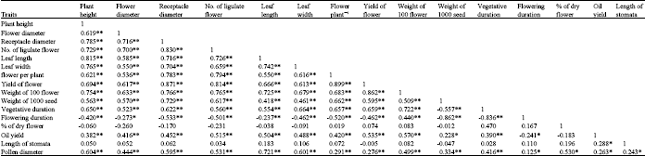 | |
| **,* Significant at 1 and 5%, respectively | |
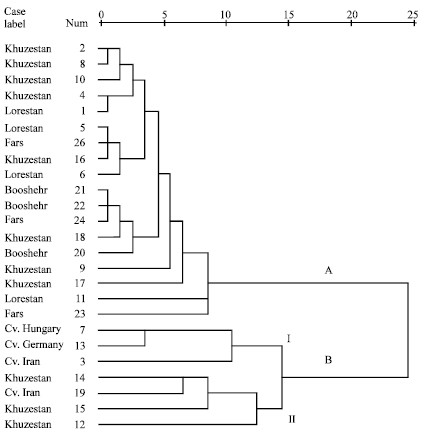 | |
| Fig. 2: | Dendrogram showing the phenetic relationships among 25 German chamomile populations based on Euclidean distances from morphological data matrix |
RAPD Assay
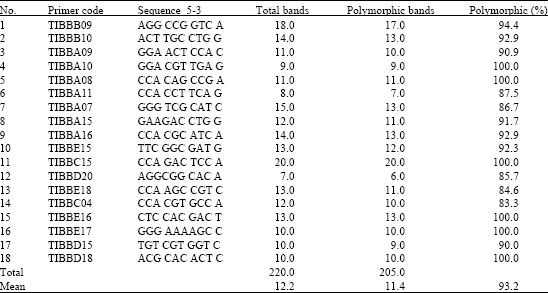
Molecular analysis of German chamomile populations revealed a considerable polymorphism, indicating a potential source of genetic materials for breeding programs. To optimize PCR and reproducibility of products, it was found that the concentration of template DNA was crucial to obtain the maximum number of reproducible bands. Varying the concentration of template DNA (5, 10, 15, 20 and 25 ng) in PCR reactions revealed that 10 ng resulted to the maximum number of reproducible bands. One hundred TIBM random primers were tested and only 18 primers with suitable and reproducible polymorphic bands among chamomile populations were selected for further analysis. These 18 primers produced 220 DNA fragments (Table 4), with 205 polymorphic DNA fragments in chamomile populations. The lowest and the highest number of polymorphism were observed with primers TIBMBD-20 and TIBMBC-15, respectively. The size of amplified fragments ranged between 300 and 3000 bp for all primers.
The similarity matrix is represented in Table 5. The lowest similarity (0.15) was between the populations No. 2 from Khuzestan and No. 23 from Fars. The highest similarity (0.78) was observed between the population No. 9 and 10 from Khuzestan province.
The cophentic correlation coefficient indicated a correlation of r = 0.95 between the similarity matrix and the UPGMA dendrogram, indicating a remarkable representation of the relationships among the populations. According to the dendrogram (Fig. 3), at a similarity level of 0.25-0.30 the populations were divided into two main groups (A and B). Group A involved all populations of Lorestan and Khuzestan provinces and all Iranian and foreign cultivars. While group B, included all populations of Booshehr and Fars provinces.
| Table 4: | Primer sequences and related data in 25 German chamomile populations |
 | |
| Table 5: | Similarity matrix among 25 German chamomile populations based on Jaccard coefficient based on RAPD bands |
 | |
In general, representation of different populations in the cluster was in congruent with geographical origins. In group A, four populations of Lorestan and eight populations from Khuzestan province were represented very close together, while population No. 15 from Lorestan province and No. 14 and 2 from Khuzestan province were represented more divergent from others and in close relationships with breaded cultivars. In group B, population No. 22 from Booshehr was more differentiated from other populations of Booshehr and Fars provinces.
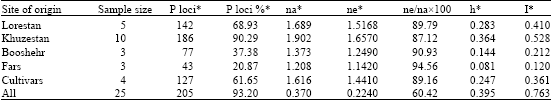
With regard to Nei's genetic diversity parameters (Table 6), observed number of alleles (na) was highest in Khuzestan province (1.902) and lowest in Booshehr province (1.373). Maximum and minimum number of effective alleles was observed in Khuzestan province (1.657) and Fars province (1.142), respectively. Consistent allelic distribution within each population was calculated via the ratio of the number of effective alleles to the number of observed alleles. The maximum consistent distribution was observed in populations of Fars province (94.56).
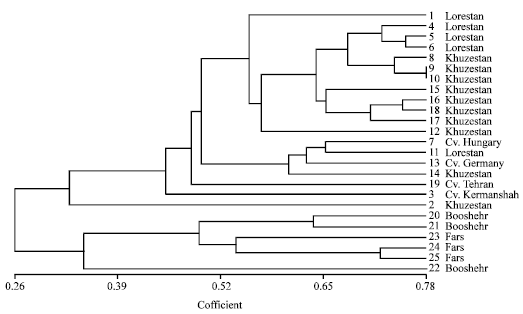 | |
| Fig. 3: | UPGMA dendrogram of 25 Iranian German chamomile populations based on RAPD primers |
| Table 6: | Genetic variability within each Province and cultivars based on RAPD data |
 | |
| *The number of polymorphic loci, *The percentage of polymorphic loci, *na: Observed No. Of alleles, *ne: Effective No. Of alleles (Kimura and Crow, 1964), *h: Nei’s (1973) gene diversity, *I: Shannon’s information index (Lewontin, 1972) | |
| Table 7: | Nei's genetic identity (above diagonal) and genetic distance (below diagonal) between different Provinces of origins and cultivars of German chamomile based on RAPD data |
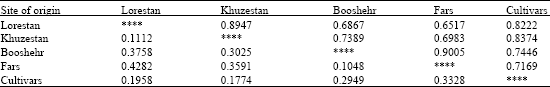 | |
The percentage of polymorphic loci (P loci%), Nei's gene diversity (h) and Shannon's information index (I), calculated for different provinces, indicated that diversity within Khuzestan province (P loci = 90.29%, h = 0.364 and I = 0.528) was more and within Fars province (P loci = 20.87% , h = 0.16 and I = 0.23) was less than other populations (Table 6). This finding confirmed that genetic diversity is highly correlated with the level of geographical distribution. Diversity coefficient among populations obtained Gst = 0.395. Estimate of gene flow from Gst (Nm) was 0.763. FST is a measure of genetic differentiation over subpopulations and its amount ranged from 0 to 1. In this investigation, FST was 0.394. When the subpopulations are identical in allele frequencies FST equals to 0, whereas, FST equals to 1 when they are fixed for different alleles (Weir, 1996). Genetic distance between provinces was calculated based Nei's original (Nei, 1972) (Table 7). Maximum (0.428) genetic distance was obtained between Lorestan and Fars provinces while minimum (0.104) genetic distance was obtained between Booshehr and Fars provinces. Measures of genetic distances were in congruent with geographic distance between provinces.
DISCUSSION
Morphological and RAPD analysis of different accessions of M. chamomilla, revealed considerable but different patterns of variability as indicated previously by Solouki et al. (2008). Morphological traits are based on phenotypic expressions of the genotypes and are influenced by environmental and ontogenetic factors while DNA markers are independent of environment and reveal transcribed and non-transcribed regions of the genome (Heywood, 2002). In general projection of studied populations in the RAPD cluster was more in agreement with geographic distribution that was reported in Persian cumin (Kouhestani et al., 2009).
Significant variation observed for different morphological traits among studied populations. Plant height was varied from 55 cm in population No. 3 to 29 cm in population No. 18 with a CV of 21.0%. In this study, almost all Iranian populations showed lower plant height than German and Hungarian cultivars in contrast to reported results of Solouki et al. (2008) while Iranian populations are diploids but German and Hungarian cultivars in this study are tetraploid. Domain of flower diameter and receptacle diameter was varied from 14.2 to 19.4 mm and 5.1 to 8.0 mm, respectively. Solouki et al. (2008) reported that flower diameter was varied from 9.1-11.4 mm. Flower diameter was varied from 6-23 mm among Hungarian populations (Gosztola et al., 2007) and from 14-17.3 mm among Italian populations (D'Andrea, 2002), while this trait was not significantly varied among European populations studied by Taviani et al. (2002).
Phenotypic variability of important traits such as number of flower per plant, flower yield and essential oil yield made promising results for future breeding program. Number of flower per plant was varied between15.7-90.5. Maximum number of flower per plant observed in population No. 12 (90.5), No. 11 (82.7), No. 13 (75.3) and No. 3 (73.6). Flower yield of population's No. 12 (78.6 g plant-1) and 15 (86.2 g plant-1) from Khuzestan province showed equal amount to breaded cultivars (79.8-89.0 g plant-1). With regard to yield of 100 flowers, population No. 15 (12.3 g) and No. 7 from Hungary were superior populations. Population No. 7 from Hungary showed highest oil yield (0.91%). Among Iranian populations, No. 6 (0.71%), 14 (0.76%), 15 (0.74%), 16 (0.74%) and 20 (0.74%), showed highest oil content even more than No. 13 from Germany (0.65%). Oil yield of M. chamomilla has been reported to be varied between 0.1-0.89 percent (D'Andrea, 2002; Solouki et al., 2008; Gosztola et al., 2007). With regards to yield of flowers, oil yield, plant height and uniformity in flowering, population No. 12 from Khuzestan province showed best performance. This population was placed in the same group along with other superior populations, No. 14, 15 and 19 and German and Hungarian cultivars. Based on RAPD assay similarity coefficients of population No. 12 with German (No.13) and Hungarian (No.7) cultivars obtained 0.54 and 0.47, respectively. Among other superior populations, No. 15 showed minimum similarity with German (Gs = 0.45) and Hungarian (Gs = 0.48) cultivars. This wide domain of genetic variability can be considered as a gene pool available to breeders for German chamomile improvement through selection and hybridization breeding programs. High heterosis effects will be expected for hybrids of these genetically diverse and talent populations.
The absence of a complete relationship between the morphological and genetic similarities was also found for wild populations of other plant (Greene et al., 2004; Steiner and los Santos, 2001). Several reasons may account for the discordance between the morphological traits and RAPD marker. First, the less number of random primers could not cover vast area of matricaria genome. Second, morphological variation is strongly associated with environmental variation; the morphological similarities observed may be due to different combinations of alleles producing similar phenotypes that might result in morphological similarities or differences that are not proportional to the underlying genetic differences. The information found here evidenced the high genetic diversity. It could be valuable to use both variations obtain from molecular and morphological to select parents of improved varieties. A breeding program can be started within any morphological and RAPD cluster found in this study without risk of inbreeding.
In conclusion, this is the first assessment on the genetic diversity of M. chmomilla populations based on combined analyses of RAPD and morphological data. It is strongly recommended that both morphological and molecular assays could be used as complementary methods in describing the population diversity in the populations. However, it is worth stressing that this work needs to be further strengthened with more exhaustive sampling of populations and more advanced molecular techniques.
ACKNOWLEDGMENT
We thank A. Sarkhosh for technical assistance. This research was supported by department of horticultural sciences, University of Tehran.
REFERENCES
- D'Andrea, L., 2002. Variation of morphology yield and essential oil components in common chamomile (Chamomilla recutita (L.) Rauschert) cultivars grown in southern Italy. J. Herb. Specie. Med. Plant, 9: 359-365.
CrossRef - De Masi, L., C. Esposito, D. Castaldo, F. Siano and B. Laratta, 2006. Assessment of agronomic, chemical and genetic variability in common basil (Ocimum basilicum L.). Eur. Food Res. Technol., 223: 273-281.
CrossRef - Greene, S.L., M. Gritsenko and G. Vandemark, 2004. Relating morphologic and RAPD variation to collection site environment in wild populations of red clover (Trifolium pratense L.). Genet. Resour. Crop. Evol., 51: 643-663.
CrossRef - Gosztola, B., E. Nemeth, A. Kozak, S.Z. Sarosi and K. Szabo, 2007. Comparative evaluation of Hungarian chamomile (Matricaria recutita) populations. Acta Hort., 749: 157-162.
Direct Link - Hadian, J., S.M.F. Tabatabaei, M.R. Naghavi, Z. Jamzad and T.R. Masoumi, 2008. Genetic diversity of Iranian accessions of Satureja hortensis L. based on horticultural traits and RAPD markers. Sci. Hort., 115: 196-202.
CrossRef - Jaccard, P., 1908. Nouvelles recherches sur la distribution florale. Bull. Soc. Vaud. Sci. Nat., 44: 223-270.
CrossRefDirect Link - Khan, M.A., S.W. Ehbrecht, B.L. Maass and H.C. Becker, 2009. Relationships among different geographical groups, agro-morphology fatty acid composition and RAPD marker diversity in Safflower (Carthamus tinctorius). Genet. Reso. Crop Evol., 56: 19-30.
CrossRef - Kimura, M. and J.F. Crow, 1963. The measurement of effective population number. Evolution, 17: 279-288.
Direct Link - Lewontin, R.C., 1972. The Apportionment of Human Diversity. In: Evolutionary Biology, Dobzhansky, T., M.K. Hecht and W.C. Steere (Eds.), Springer, New York, ISBN: 978-1-4684-9063-3, pp: 381-398.
CrossRefDirect Link - Lynch, M. and B.G. Milligan, 1994. Analysis of population genetic structure within RAPD markers. Mol. Ecol., 3: 91-99.
PubMed - McKay, D.L. and J.B. Blumberg, 2006. A review of the bioactivity and potential health benefits of chamomile tea (Matricaria recutita L.). Phytother. Res., 20: 519-530.
CrossRefPubMedDirect Link - Mohammadi, S.A. and B.M. Prasanna, 2003. Analysis of genetic diversity in crop plants-salient statistical tools and considerations. Crop Sci., 43: 1235-1248.
CrossRefDirect Link - Murray, M.G. and W.F. Thompson, 1980. Rapid isolation of high molecular weight plant DNA. Nucl. Acids Res., 8: 4321-4326.
CrossRefPubMedDirect Link - Nei, M., 1973. Analysis of gene diversity in subdivided populations. Proc. Natl. Acad. Sci. USA., 70: 3321-3323.
PubMedDirect Link - Porter, W.M. and D.H. Smith, 1982. Detection of identification errors in germplasm collections. Crop Sci., 22: 701-703.
CrossRefDirect Link - Solouki, M., H. Mehdikhani, H. Zeinali and A.A. Emamjomeh, 2008. Study of genetic diversity in chamomile (Matricaria chamomilla) based on morphological traits and molecular markers. Sci. Hortic., 117: 281-287.
CrossRef - Steiner, J.J. and G.G. los Santos, 2001. Adaptive ecology of Lotus corniculatus L. genotypes: I plant morphology and RAPD maker characterizations. Crop. Sci., 41: 552-563.
Direct Link - Taviani, P., D. Rosellini and F. Veronesi, 2002. Variation for agronomic and essential oil traits among wild population of Chamomilla recutita L. from central Italy. J. Herb. Spices Med. Plants, 9: 353-358.
Direct Link - Wagner, C., F.T. Wolfgang, R.A. Marquard and F. Ordon, 2005. Molecular analyses on the genetic diversity and inheritance of (-)-α-bisabolol and chamazulene content in tetraploid chamomile (Chamomilla recutita (L.) Rausch.). Plant Sci., 169: 917-927.
Direct Link - Williams, J.G.K., A.R. Kubelik, K.J. Livak, J.A. Rafalski and S.V. Tingey, 1990. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res., 18: 6531-6535.
CrossRefPubMedDirect Link - Wright, S., 1949. The genetical structure of populations. Ann. Eugenics, 15: 323-354.
CrossRefPubMedDirect Link - Yeh, F.C., R.C. Yang, T.B.J. Boyle, Z.H. Ye and J.X. Mao, 1997. POPGENE, the User-Friendly Shareware for Population Genetic Analysis. Molecular Biology and Biotechnology Center, Alberta.
Direct Link