
Waleed Alabdulmonem
Department of Pathology, College of Medicine, Qassim University, P.O. Box 6655, 51452 Buraidah, Saudi Arabia
LiveDNA: 966.23477
Zafar Rasheed
Department of Medical Biochemistry, College of Medicine, Qassim University, Buraidah, Saudi Arabia
Abdullah S.M. Aljohani
Department of Veterinary Medicine, College of Agriculture and Veterinary Medicine, Qassim University, Buraidah, Saudi Arabia
Mohsen Almakrami
School of Biological sciences, University of Essex, Colchester, United Kingdom
Hussain Al Ssadh
School of Biological sciences, University of Essex, Colchester, United Kingdom
LiveDNA: 966.29413
Nelson Fernandez
School of Biological sciences, University of Essex, Colchester, United Kingdom
Research Journal of Immunology
Year: 2020 | Volume: 13 | Issue: 1 | Page No.: 1-9
DOI: 10.3923/rji.2020.1.9







ABSTRACT
Background and Objective: The cellular component of innate immunity relies on cluster of differentiation (CD) 14 and toll-like receptor (TLR)-4 to combat harmful pathogens. This study was undertaken to investigate the effect of lipopolysaccharide (LPS) subtypes smooth (O55:B5) and LPS rough (EH100) on the expression of CD14 and TLR-4 in human leukemia macrophage-derived THP-1 cells. Materials and Methods: Monocyte to macrophage differentiation was achieved in THP-1 cells by phorbol-12-myristate-13-acetate (PMA) treatment. Surface expression of CD14 and TLR-4 were determined by flow cytometry after treatment with LPS subtypes. TNF-α was measured by ELISA. Confocal microscopy was used to determine co-localization of CD14 and TLR-4 receptors on THP-1 cell surface. Results: Treatment of THP-1 cells with LPS subtypes significantly increased the expression of CD14 and TLR-4 receptors (p<0.05). Higher levels of TNF-α were released in LPS-subtypes stimulated cells but the differences in the levels of TNF-α in differentiated and undifferentiated cells still unclear. Co-localization showed positive correlation between CD14 and TLR-4 receptors (r>0.83). Conclusion: This study revealed that monocyte to macrophage differentiation in THP-1 cells increases the surface expression of CD14 and TLR-4 receptors and also leads overproduction of TNF-α.
PDF Abstract XML References Citation
Copyright: © 2020. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Waleed Alabdulmonem, Zafar Rasheed, Abdullah S.M. Aljohani, Mohsen Almakrami, Hussain Al Ssadh and Nelson Fernandez, 2020. LPS Subtypes Activate Inflammatory Signaling Through CD-14 and TLR-4 in Human Monocytic Cells. Research Journal of Immunology, 13: 1-9.
DOI: 10.3923/rji.2020.1.9
URL: https://scialert.net/abstract/?doi=rji.2020.1.9
DOI: 10.3923/rji.2020.1.9
URL: https://scialert.net/abstract/?doi=rji.2020.1.9
INTRODUCTION
Lipopolysaccharide (LPS) is the major constituent of the outer cell wall of gram-negative bacteria and is the principal mediator of inflammatory responses to these pathogens1,2. The LPS is recognized by number of serum and cell surface pattern recognition receptors (PRRs), which participate to induce LPS mediated cellular events1,2. The LPS signalling is mediated through interaction between acute phase protein the cluster of differentiation (CD) 14 and LPS associated co-receptors3,4. Thus the importance of this interaction is that CD14 is unable to transduce the LPS signaling because it lacks of trans-membrane domain4. Therefore, CD14 presumably function by binding to LPS and transferring it other cell surface receptors to trigger the LPS signaling5. Toll-like receptor 4 (TLR-4) is associated with CD14 in inflammatory responses to LPS because TLR4 is a trams-membrane receptor type 16,7 and the presence of myeloid differentiation protein (MD)-2 makes up the LPS receptor complex involved in the cellular recognition of and signaling by LPS8. Recent advances implicate high-mobility group box 1 (HMGB1), a nuclear protein with inflammatory cytokine activities, in stimulating cytokine release8. To date, many candidate receptors were implicated in mediating the biological responses to HMGB1 and TLR2, TLR4, TLR9 but the mechanisms for the production of inflammatory mediators via these receptors protein still not completely explored. Scavenger receptors are another family of receptors expressed on macrophage and recognize and bind both endogenous and exogenous molecules. The SR-A (MARCO) receptor with collagenous structure, plays a pivotal role in the transduction signaling of LPS via TLRs9. The present study was designed to investigate the effect of LPS subtypes LPS smooth (O55:B5) and LPS rough (EH100) on the expression of innate recognition receptors CD14 and TLR-4 on the surface of human macrophage-derived THP-1 cells. Secondly, to investigate the levels of TNF-α production in PMA treatment cells with or without LPS addition. Thirdly, to determine the co-localization of CD14 and TLR-4 receptors on the surface of PMA-differentiated THP-1 cells treated with both LPS serotypes. In short, this is the first study that shows the differentiation of monocyte to macrophage in LPS subtypes stimulated human leukemia THP-1 cells increases the surface expression of CD14 and TLR-4 receptors and also leads to the overproduction of TNF-α.
MATERIALS AND METHODS
This study was performed in School of Biological Sciences, University of Essex, Colchester, United Kingdom from April, 2017 to January, 2019. Interpretation of results and manuscript preparation were done in the College of Medicine, Qassim University, Buraidah, Saudi Arabia.
Treatment of THP-1 cells: The THP-1 cells (5×105 cells mL–1) were treated with varying concentrations (0-30 ng mL–1) of PMA (5 ng mL–1) for 24-96 h. The THP-1 cells were also treated with LPS subtypes EH100 or O55:B5 under the same experimental conditions as described previously10.
Flow cytometry and immunostaining: Flow cytometry analyses were performed by the Aria flow cytometer (FACS Aria II Flow Cytometer, BD Biosciences, CA, USA) and the results analyzed with flowing software (BD Biosciences, CA, USA) as described previously10,11. Immunofluorescent staining was performed after different stimulatory conditions as described previously10-14.
Confocal microscopy: The PMA-differentiated THP-1 cells were cultured at 25×103 cells on cover slips of area 2 cm2×thickness 0.13 cm for 72 h after treatment with 5 ng mL–1 of PMA. The cells were then stimulated with 100 ng mL–1 of either EH100 or O55:B5 LPS for 4 h and the confocal microscopy was performed as described previously10-12. For staining of cell nuclei, 4,6-diamino-2-phenylindole (DAPI, Sigma-Aldrich, Poole, UK) was used and the slides were mounted with Vectashield Mounting Media (Vector Laboratories, Burlingame, CA, USA) and coverslips were sealed with Marabu Fixogum Rubber Cement (Marabuwerke GmbH and Co. KG, Tamm, Germany). The slides were examined with a Nikon confocal microscopy and Pearson correlation coefficient was used to analyze the co-localization between CD14 and TRL-4.
Human TNF-α specific sandwich ELISA: The production of TNF-α level in culture supernatant was measured by human TNF-α specific sandwich ELISA as per instructions of the manufacturer (eBioscience™, Fisher Scientific Company, Ontario, Canada). The optical density of each well was measured at 450 nm using ELISA plate reader (Versa Max, California, USA).
Statistical analysis: All statistical analysis was carried out by Graph Pad Prism version 5.0 (Graph Pad Software Inc., San Diego, CA, USA). One-way ANOVA of variance followed by Tukey-Kramer multiple comparisons test, or Two-way ANOVA of variance followed by Bonferroni comparisons test. The p<0.05 was considered significant. Results are expressed as the Mean+SEM unless stated otherwise.
RESULTS
Morphological alterations in THP-1 cells: Treatment of the monocytic THP-1 cells with PMA caused typical morphology alterations to macrophages. Figure 1 shows morphological alterations in THP-1 cells upon stimulation with PMA. The untreated cells were undifferentiated and appeared with egg-like morphology while floating in the culture medium (Fig. 1a). On the other hand, PMA-induced differentiation of the cells caused them to display macrophage-like characteristics such as adherence and possession of long spindle-like morphology (Fig. 1b). The features acquired by the cells following differentiation included were cell enlargement, increased in granular appearance, adherence to the plate and spreading all of which are hallmarks of macrophages.
Surface expression on THP-1 cells: Measurements of expression of CD14 and TLR-4 on the surface of THP-1 cells with and without differentiation and in the presence of either rough (EH100) or smooth (O55:B5) LPS chemo-types were carried out by means of flow cytometry using FITC stained secondary antibodies and the results were summarized in Fig. 2. The results shown in Fig. 2a-c pointed out that the surface expressions of both CD14 and TLR-4 were not significantly affected by both type of LPS in the THP-1 cells (p>0.05). However, cell surface expression of CD14 was slightly decreased and TLR-4 expression was significantly increased in the THP-1 cells (p<0.05, Fig. 2d). Furthermore, stimulation of non-differentiated cells with smooth, but not rough, for 4 h caused upregulation of CD14 expression and had no effect of TLR-4 levels compared to untreated controls (Fig. 3a). Conversely, the treatment of differentiated cells with smooth LPS caused a weak upregulation of CD14 and a strong upregulation of TLR-4 (Fig. 3b). The stimulation with rough LPS resulted in a strong upregulation of CD14 expression but not TLR-4 expression. Differentiated THP-1 cells showed significantly higher levels of receptor expression compared to undifferentiated cells (p<0.05, Fig. 3).
Production of TNF-α by differentiated and non-differentiated THP-1 cells: The TNF-α levels in the culture supernatants of THP-1 cells stimulated with LPS subtypes were found to be significantly higher than the untreated cells (p<0.01, Fig. 4).The levels of TNF-α production by the differentiated and non-differentiated cells upon 2-6 h treatment with LPS subtypes have been summarized in Fig. 4. The results showed that LPS subtype O55:B5 was more effective than EH100.
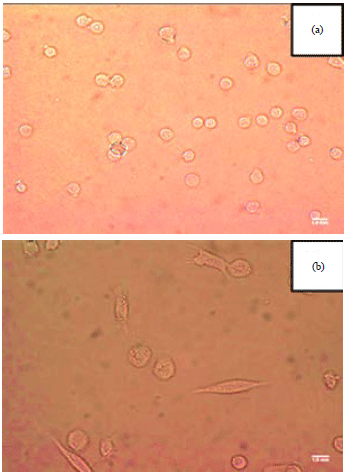 | |
| Fig. 1(a-b): | THP-1 cells (a) Without PMA treatment and (b) With PMA treatment |
Concentration of PMA used was 5 ng mL–1 | |
Moreover, results also pointed out that after 2 h treatment with both subtypes the TNF-α levels were significantly increased by the differentiated cells as compared with non-differentiated THP-1 cells (p<0.05). However, these results were reversed after 4 or 6 h treatment with these LPS subtypes (p<0.01).
Co-localization of CD14 and TLR-4 on the THP-1 cell-surface: The co-localization of CD14 and TLR-4 on the THP-1 cell-surface was studied by confocal laser scanning microscopy. The degree of co-localization of CD14 and TLR-4 on the THP-1 cell surface upon stimulation with LPS subtypes was shown in Fig. 5.The degree of co-localization of CD14 in THP-1 stimulated with LPS-subtype EH100 was shown in Fig. 5A1, whereas, co-localization of TLR-4 in same stimulated THP-1 cells with EH100 was shown in Fig. 5A2. Not only have these, the degree of co-localization of CD14 and TLR-4 together with EH100 was shown in Fig. 5A3. On the other hand, the degree of co-localization of CD14 with LPS-subtype O55:B5 was shown in Fig. 5B1, whereas co-localization of TLR-4 with LPS-subtype O55:B5 was shown in Fig. 5B2.
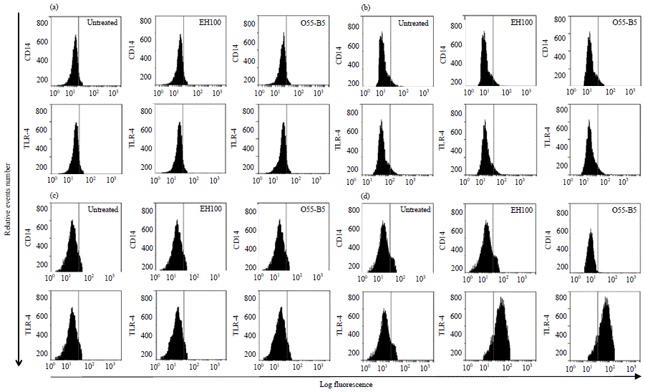 | |
| Fig. 2(a-d): | Expression of CD14 and TLR-4 in (a) Non-differentiated THP-1 cells after 2 h treatment with LPS subtypes, (b) After 6 h, (c) Differentiated THP-1 cells after 2 h and (d) 6 h |
The cells (5×105) being cultured in RPMI 1640 medium supplemented with 10% FCS were stimulated with both rough (EH100) and smooth (O55:B5) LPS and then incubated for 2-6 h | |
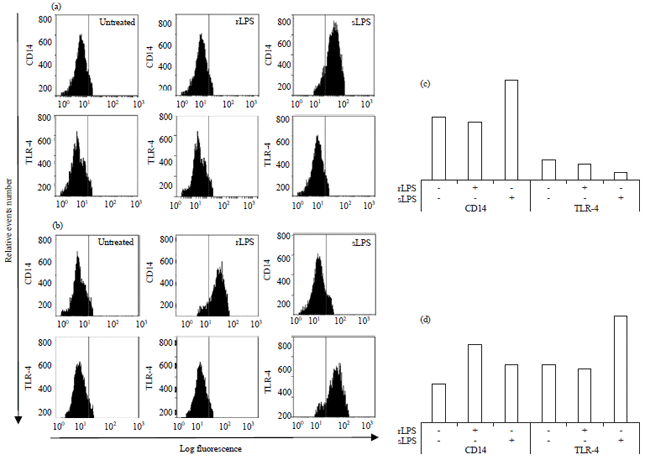 | |
| Fig. 3(a-d): | Expression of CD14 and TLR-4 on the non-differentiated (a) THP-1 cells and (b) Differentiated THP-1 cells after 4 h LPS subtypes stimulation. The bar graphs show the expression of CD14 and TLR-4 in the presence or absence of rough or smooth LPS, respectively on the same, (c) Non-differentiated and (d) Differentiated THP-1 cells |
Non T: Non treated cells, R: Rough (EH100) LPS, S: Smooth (O55:B5) LPS it is also follows the same order as the bar chart | |
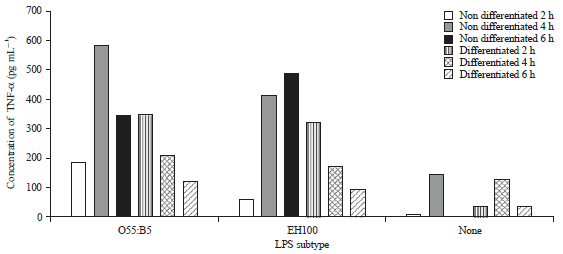 | |
| Fig. 4: | Production of TNF-α by differentiated and non-differentiated THP-1 cells after stimulation with either smooth (O55:B5) or rough (EH100) LPS subtypes |
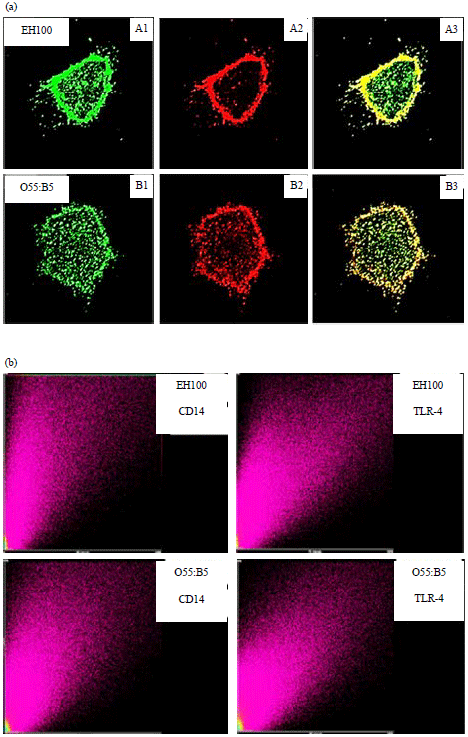 | |
| Fig. 5(a-b): | (a) Co-localization of CD14 and TLR-4 on the THP-1 cell-surface examined by confocal laser scanning microscopy and (b) Scatter plots show co-localization of CD14 and TLR-4 on the THP-1 cells stimulated by LPS subtypes as indicated |
| A1/B1, green: THP-1 cells stained with either CD14 primary antibody and labeled with Alexa Fluor® 488, A2/B2 (red): TLR-4 primary antibody and labeled with Alexa Fluor® 555, A3/B3 (red and green): Merged images, where co-localization, if present, can be observed | |
Whereas, co-localization of CD14 and TLR-4 together with O55:B5 was shown in Fig. 5B3. These results were further supported by the Scatter plots also showing co-localization of CD14 and TLR-4 on the THP-1 cells stimulated by LPS subtypes as indicated in Fig. 5B.
DISCUSSION
This study attempted to investigate the immunomodulation of two innate recognition receptors on human monocytes/macrophages and to explore the differences in the surface expression of CD14 and TLR-4 in THP-1 cells with and without differentiation. The differentiation in THP-1 cells was carried out by PMA treatment, which is a well-known potent stimulator for monocyte to macrophage transformation15. Previous studies have shown that the innate recognition receptors CD14, TLR4 have a key role in the pathogenesis of sepsis and septic shock and other inflammatory diseases16,17. However, the roles of PMA in these processes have not been fully explored. In recent years, the knowledge in the field of monocytes subpopulations and their differentiation has been gained significantly but still it is many things associated with their behavior are largely unknown and need to be investigated18,19. But it is now well established that monocytes can sense with the environmental changes as they express number of receptors through which they controls their functions18. Although, monocytes are highly heterogeneous this can change their functional phenotypes under various abnormal conditions as in inflammatory disorders20. For this reason many investigators have differentiated monocytes into 2 subtypes inflammatory or anti-inflammatory subsets. Under several pathological conditions or infection, monocytes rapidly recruited at the site of infection, where they can differentiate into tissue macrophages and also sometimes dendritic cells21,22. Inflammatory monocytes are very important cell types as they are having TLRs and other receptors which are very quick for their action towards the neutralization of pathogens and their associated molecules and also removing microorganisms, lipids and dying cells through phagocytosis18-22. To study the behaviors of monocytes upon stimulation of novel stimulants LPS subtypes, the human monocytic THP-1 cells were selected as they are well considered as a model for human monocytes23.
In the present study, the expression of CD14 and TLR-4 was on the surface of THP-1 cells with and without differentiation and in the presence of either rough (EH100) or smooth (O55:B5) LPS chemo-types. The expression of CD14 on THP-1 slightly up regulated in presence LPS, whereas, the expression of TLR-4 receptor was not affected. To determine whether the expression of these receptors were affected by induced differentiation cells were treated with different concentrations of PMA, A dose of 5 ng mL–1 of PMA showed maximum expression of CD14, TLR4 compared with undifferentiated THP-1 cells. Cell activation occurs as a result of cell response to LPS which was triggered via CD14 receptors, this activation required 15-30 min between the LPS binding to the cells. This clearly suggested that the activation process was a time consuming which might involve internalization before induction of the signaling pathway. However, the maximum response in the cells was achieved after 4 h of LPS stimulation. The data showed that stimulation of both undifferentiated THP-1 and PMA-differentiated THP-1 cells with LPS subtypes up-regulated the expression of both CD14 and TLR4. These results were fully supported by other previously published reports showed that monocytic cells treated with PJ34, an inhibitor of PARP, in the presence of LPS led to a down-regulation of surface expression of both CD14 and TLR-4 receptors24,25.
The data also determined that native THP-1 cells released less amount of TNF-α compared with PMA differentiated THP-1 cells in response to LPS subtypes. Confocal microscopy was used to detect and quantify the co-localization of CD14 and TLR-4 receptors. The data showed that the THP-1 cells showed no co-localization between CD14 and TLR-4, while PMA-differentiated THP-1 cells showed co-localization between CD14 and TLR-4 and these results clearly explained that the PMA-differentiated THP-1 cells produced more pro-inflammatory cytokines such as TNF-α compared with native THP-1 cells. Excess production of TNF-α upon PMA treatment on THP-1 cells was well supported by number of previous studies26. The TNF-α values obtained in this study clearly reflected a positive correlation or potential co-localization between CD14 and TLR-4 receptors. These results were also supported by previous studies performed on other cell types27. Stimulation of THP-1 and PMA-THP-1cells with LPS subtypes showed co-localization between CD14 and TLR-4 and the high production of pro-inflammatory cytokines also justified by up-regulation of CD14 receptors. These findings suggested that there might be other receptors also participate in production of TNF-α in response to LPS. Stimulation of native and PMA-differentiated THP-1 cells with LPS subtypes led to increase co-localization between CD14 and TLR-4 receptors and the high levels of TNF-α released could be justified by the corresponding up-regulation of CD14 receptors after stimulation with LPS subtypes. In short, this study was an attempt to discover the novel pathophysiological aspects of LPS subtypes induced inflammatory response through cellular receptors proteins of CD14 and TLR-4 in human monocytic-macrophage cell line THP-1. The CD14 and TLR-4 have several unique biological functions, especially in their central roles in innate immunity. Therefore, the knowledge gained from this study will help researchers to expand the options for effective intervention so that inflammatory diseases mediated by CD14 or TLR4 can be treated or prevented. Furthermore, findings from this study have also been implicated in a novel therapeutic application for the management of immune-mediated inflammatory disorders. As the results clearly indicated that the pharmacological blockade of CD14 and TLR-4 represents a useful therapeutic tool for the therapy of immune-mediated inflammatory disorders. Despite of these important implications and therapeutic application, this study has few limitations. The most obvious limitation is the study limited to the THP-1 cells only. Although, THP-1 cells has now been well considered as a model for human monocytes23. But it will be better to perform similar study on the primary monocytes which will obtain directly either from the acute monocytic leukemia patients or from the patients with immune-mediated inflammatory disorders. In addition, this study also recommends further studies that will be conducted on CD14 or TLR-4 knockout mouse model, so that the improved understanding on the connection between CD14 or TLR-4 with the onset of immune mediated inflammatory conditions will be drawn.
CONCLUSION
This is the first study that shows the differentiation of monocyte to macrophage in THP-1 cells increases the surface expression of CD14 and TLR-4 receptors and also leads overproduction of TNF-α. The positive correlation between these receptors concludes that they may interact with each other in their activities. Improved understanding on the connection between CD14 and TLR4 may lead to the development of novel therapies for controlling the onset of immune mediated inflammatory conditions.
SIGNIFICANCE STATEMENT
This study was an attempt to discover the novel pathophysiological aspect of LPS subtypes induced inflammatory response through cellular receptors proteins CD14 and TLR-4 in human monocytic-macrophage THP-1 cells. CD14 and TLR-4 have several unique biological functions, especially of their central roles in innate immunity. Therefore, the knowledge gained from this study will help researchers to expand the options for effective intervention so that inflammatory diseases mediated by CD14 or TLR-4 can be treated or prevented.
ACKNOWLEDGMENT
Authors are grateful acknowledged to the support of College of Medicine, Qassim University, KSA (Grant # QU1731439) and also School of Biological Sciences, University of Essex, UK.
REFERENCES
- Blairon, L., X. Wittebole and P.F. Laterre, 2003. Lipopolysaccharide-binding protein serum levels in patients with severe sepsis due to gram-positive and fungal infections. J. Infect. Dis., 187: 287-291.
CrossRefPubMedDirect Link - Da Silva Correia, J., K. Soldau, U. Christen, P.S. Tobias and R.J. Ulevitch, 2001. Lipopolysaccharide is in close proximity to each of the proteins in its membrane receptor complex: Transfer from CD14 TO TLR4 and MD-2. J. Biol. Chem., 276: 21129-21135.
CrossRefDirect Link - Schröder, N.W.J., B. Opitz, N. Lamping, K.S. Michelsen, U. Zähringer, U.B. Göbel and R.R. Schumann, 2000. Involvement of lipopolysaccharide binding protein, CD14, and Toll-like receptors in the initiation of innate immune responses by Treponema glycolipids. J. Immunol., 165: 2683-2693.
CrossRefPubMedDirect Link - Haziot, A., E. Ferrero, F. Köntgen, N. Hijiya and S. Yamamoto et al., 1996. Resistance to endotoxin shock and reduced dissemination of gram-negative bacteria in CD14-deficient mice. Immunity, 4: 407-414.
CrossRefPubMedDirect Link - Haziot, A., N. Hijiya, S.C. Gangloff, J. Silver and S.M. Goyert, 2001. Induction of a novel mechanism of accelerated bacterial clearance by lipopolysaccharide in CD14-deficient and Toll-like receptor 4-deficient mice. J. Immunol., 166: 1075-1078.
CrossRefPubMedDirect Link - Singh, B.P., R.S. Chauhan and L.K. Singhal, 2003. Toll-like receptors and their role in innate immunity. Curr. Sci., 85: 1156-1164.
Direct Link - Yang, H. and K.J. Tracey, 2010. Targeting HMGB1 in inflammation. Biochim. Biophys. Acta (BBA)-Gene Regul. Mech., 1799: 149-156.
CrossRefDirect Link - Schromm, A.B., E. Lien, P. Henneke, J.C. Chow and A. Yoshimura et al., 2001. Molecular genetic analysis of an endotoxin nonresponder mutant cell line: A point mutation in a conserved region of MD-2 abolishes endotoxin-induced signaling. J. Exp. Med., 194: 79-88.
CrossRefPubMedDirect Link - Bowdish, D.M., K. Sakamoto, M.J. Kim, M. Kroos and S. Mukhopadhyay et al., 2009. MARCO, TLR2 and CD14 are required for macrophage cytokine responses to mycobacterial trehalose dimycolate and Mycobacterium tuberculosis. PLoS Pathog., Vol. 5, No. 6.
CrossRefDirect Link - Jiang, Q., S. Akashi, K. Miyake and H.R. Petty, 2000. Cutting edge: Lipopolysaccharide induces physical proximity between CD14 and toll-like receptor 4 (TLR4) prior to nuclear translocation of NF-κB. J. Immunol., 165: 3541-3544.
CrossRefDirect Link - Alabdulmonem, W., H.T. Alhomaidan, Z. Rasheed, I.H. Madar, N. Alasmael, S. Alkhatib and H. Al Ssadh, 2018. CD74 a potential therapeutic target for breast cancer therapy: Interferon gamma up-regulates its expression in CAMA-1 and MDA-MB-231 cancer cells. Int. J. Cancer Res., 14: 58-69.
CrossRefDirect Link - Ulevitch, R.J. and P.S. Tobias, 1995. Receptor-dependent mechanisms of cell stimulation by bacterial endotoxin. J. Annu. Rev. Immunol., 13: 437-457.
CrossRefPubMedDirect Link - Al Ssadh, H. and W. Al Abdulmonem, 2019. Immunophenotyping of the cluster of differentiation 74, migration inhibitory factor and cluster of differentiation 44 expression on human breast cancer-derived cell lines. Int. J. Health Sci., 13: 17-24.
PubMedDirect Link - Al Ssadh, H. and W. Al Abdulmonem, 2019. Novel predication of protein biomarkers in interferon-gamma-stimulated breast cancer cells. Int. J. Health Sci., 13: 35-43.
PubMedDirect Link - Al Ssadh, H., P.S. Spencer, W. Alabdulmenaim, R. Alghamdi, I.H. Madar, J.M. Miranda-Sayago and N. Fernandez, 2017. Measurements of heterotypic associations between cluster of differentiation CD74 and CD44 in human breast cancer-derived cells. Oncotarget, 8: 92143-92156.
CrossRefDirect Link - Noubir, S., Z. Hmama and N.E. Reiner, 2004. Dual receptors and distinct pathways mediate interleukin-1 receptor-associated kinase degradation in response to lipopolysaccharide involvement of CD14/TLR4, CR3 and phosphatidylinositol 3-Kinase. J. Biol. Chem., 279: 25189-25195.
CrossRefPubMedDirect Link - Lichtman, S.N., J. Wang and J.J. Lemasters, 1998. LPS receptor CD14 participates in release of TNF-α in RAW 264.7 and peritoneal cells but not in Kupffer cells. Am. J. Phsiol. Gastrointest. Liver Phsiol., 275: G39-G46.
CrossRefPubMedDirect Link - Yona, S. and S. Jung, 2010. Monocytes: Subsets, origins, fates and functions. Curr. Opin. Hematol., 17: 53-59.
CrossRefPubMedDirect Link - Strauss-Ayali, D., S.M. Conrad and D.M. Mosser, 2007. Monocyte subpopulations and their differentiation patterns during infection. J. Leukoc. Biol., 82: 244-252.
CrossRefDirect Link - Gordon, S. and P.R. Taylor, 2005. Monocyte and macrophage heterogeneity. Nat. Rev. Immunol., 5: 953-964.
CrossRefDirect Link - Yang, J., L. Zhang, C. Yu, X.F. Yang and H. Wang, 2014. Monocyte and macrophage differentiation: Circulation inflammatory monocyte as biomarker for inflammatory diseases. Biomark. Res., Vol. 2, No. 1.
CrossRefDirect Link - Italiani, P. and D. Boraschi, 2014. From monocytes to M1/M2 macrophages: Phenotypical vs. functional differentiation. Front. Immunol., Vol. 5.
CrossRefDirect Link - Bosshart, H. and M. Heinzelmann, 2016. THP-1 cells as a model for human monocytes. Ann. Transl. Med., Vol. 4, No. 21.
CrossRefDirect Link - Marchant, A., J. Duchow, J.P. Delville and M. Goldman, 1992. Lipopolysaccharide induces up‐regulation of CD14 molecule on monocytes in human whole blood. Eur. J. Immunol., 22: 1663-1665.
CrossRefPubMedDirect Link - Kameoka, M., O.T.A. Katsuya, T. Tetsuka, Y. Tanaka, A. Itaya, T. Okamoto and K. Yoshihara, 2000. Evidence for regulation of NF-κB by poly (ADP-ribose) polymerase. Biochem. J., 346: 641-649.
CrossRefPubMedDirect Link - Kim, J.K., S.M. Lee, K. Suk and W.H. Lee, 2011. A novel pathway responsible for lipopolysaccharide-induced translational regulation of TNF-α and IL-6 expression involves protein kinase C and fascin. J. Immunol., 187: 6327-6334.
CrossRefPubMedDirect Link - Levy, E., G. Xanthou, E. Petrakou, V. Zacharioudaki, C. Tsatsanis, S. Fotopoulos and M. Xanthou, 2009. Distinct roles of TLR4 and CD14 in LPS-induced inflammatory responses of neonates. Pediatr. Res., 66: 179-184.
CrossRefPubMedDirect Link