
Abdel-hamid Abdallah Abakar
Laboratory of Microbiology, Pharmacology, Biotechnology and Environment, Faculty of Sciences, A�n Chock, University Hassan II, Km 8 route d`ElJadida, B.P. 5366 Maarif, Casablanca, Morocco
LiveDNA: 212.20070
Ali Daher
Polz Laboratory for Microbial Ecology and Evolution, Massachusetts Institute of Technology, 15 Vassar Street, Bldg. 48-417 Cambridge, Massachusetts, USA
Khalid Belghmi
Laboratory of Microbiology, Pharmacology, Biotechnology and Environment, Faculty of Sciences, A�n Chock, University Hassan II, Km 8 route d`ElJadida, B.P. 5366 Maarif, Casablanca, Morocco
Djolo Yigerta Dah Dossounon
Laboratory of Microbiology, Pharmacology, Biotechnology and Environment, Faculty of Sciences, A�n Chock, University Hassan II, Km 8 route d`ElJadida, B.P. 5366 Maarif, Casablanca, Morocco
Mohamed Blaghen
Laboratory of Microbiology, Pharmacology, Biotechnology and Environment, Faculty of Sciences, A�n Chock, University Hassan II, Km 8 route d`ElJadida, B.P. 5366 Maarif, Casablanca, Morocco
Research Journal of Environmental Toxicology
Year: 2017 | Volume: 11 | Issue: 3 | Page No.: 104-111







ABSTRACT
Background and Objective: Heavy metals showed an important environmental problem today. They can be toxic when they are present in excess. Their dangerousness is not only their ubiquitous presence, but also the fact that they exist in several chemical forms. Among the main metal extraction is mercury, its extraction has modified the proportion and the chemical form of this metal in various compartments of ecosystems making it bioavailable and potentially harmful for organisms especially human being. This study evaluated the toxicity of this metal and a bioluminescence method has been optimized. Methodology: Bioluminescence method consisted to optimize the intensity and stability of luminescence emitted by bacteria Vibrio fischeri (V. fischeri) strain NRRL-B-11177 cultivated in a modified LBS medium. Through this method, toxic effects of inorganic mercury (HgCl2) on V. fischeri have been determined. All data were analyzed using GraphPad Prism and values were obtained as means±standard errors. Results: Results showed aeration (600 mL min–1) and agitation (250 rpm) were essential to V. fischeri to emit light. It has found that at concentration 1.4 mg L–1, HgCl2 was capable to inhibit totally the light emitted by V. fischeri. Conclusion: Thereafter, evaluated the detoxification of HgCl2 in fluidized bed reactor using immobilized and free bacteria Escherichia coli strain R100 by the luminescence of V. fischeri.
PDF Abstract XML References Citation
Received: September 27, 2017;
Accepted: November 02, 2017;
Published: November 13, 2017
How to cite this article
Abdel-hamid Abdallah Abakar, Ali Daher, Khalid Belghmi, Djolo Yigerta Dah Dossounon and Mohamed Blaghen, 2017. Detoxification Assessment of Inorganic Mercury by Bioluminescence of Vibrio fischeri. Research Journal of Environmental Toxicology, 11: 104-111.
DOI: 10.3923/rjet.2017.104.111
URL: https://scialert.net/abstract/?doi=rjet.2017.104.111
DOI: 10.3923/rjet.2017.104.111
URL: https://scialert.net/abstract/?doi=rjet.2017.104.111
INTRODUCTION
Heavy metals constitute an important current environmental problem1. Trace metals elements are elements whose concentration in the earth's crust is less than 1 g kg–1 or heavy metals (metals which have densities greater than or equal to 4.5 g cm–3) may be toxic when they present in excess in the environment 2,3. Their dangerousness was not only their ubiquitous presence, but also the fact that they existed in several chemical forms4. Each of these forms had unique characteristics which could modify their solubility or give them the power to pass through the biological membranes5-7. Environment contamination by heavy metals came from their many uses in anthropogenic activities, including industry. For these activities, metals naturally present in the earth's crust, were extracted in large quantities. This extraction has altered the proportion and chemical form of these metals in various compartments of ecosystems, thus making them bioavailable and potentially harmful to organisms. Toxicity measurement of toxicants such as heavy metals is a most important part to evaluate environmental pollution. In order to assess their toxicities several tools and methodologies were available to ecotoxicologists. Among these tools and methods, tests carried out on living organisms, such as algae, crustaceans, fish and bacteria, generally these methods’ aim was to determine the inhibition of an organism8. In case of bacteria several types of tests could be considered on isolated bacterial populations. Toxic effects were studied from bacterial growth, bacterial enzymatic activity, adenosine triphosphate content or inhibition of bioluminescence9-12. Therefore, to evaluate the pollution, it was preferable to use a simple, rapid, sensitive and cost effective such as bioluminescence inhibition method could indicate specific information on the toxicity of heavy metals13. The objective of this study was to find the best conditions of intensity and stability of luminescence of Vibrio fischeri (V. fischeri) and to assess the detoxification of Inorganic mercury (HgCl2)in fluidized bed reactor.
MATERIALS AND METHODS
This study carried out from March, 2016-April, 2017 at Laboratory of Microbiology, Pharmacology, Biotechnology and Environment, Faculty of Sciences Aïn Chock, University Hassan II.
Bacterial strains: Vibrio fischeri (V. fischeri) strain NRRL-B-11177 and Escherichia coli (E. coli) strain R100.
Bacterial growth media: V. fischeri strain was grown at 25°C in a modified Luria-Bertani salt (LBS) medium (10 g nutrient broth, 5 g yeast extract, 20 g NaCl, 50 mL 1 M Tris buffer, pH 7.5±0.02, in 1 L of distilled water).
Escherichia coli was grown at 37°C in broth medium (2% w/v nutrient broth in distilled water) and in minimal medium (MM) (1.36 g of potassium hydrogen phosphate (k2HPO4), 0.10 g of magnesium sulfate (MgSO4), 0.60 g of ammonium sulfate ( [NH4]2SO4), 0.02 g of calcium chloride (CaCl2) and 0.50 g of sodium chloride (NaCl2), 220 μL of manganese (II) sulfate (MnSO4), 40 μL zinc sulfate (ZnSO4), 40 μL of copper (II) sulfate (CuSO4.5H2O) and 28 μL of iron (II) sulfate (FeSO4), in 1 L of distilled water). All products were purchased from Biokar Diagnostics.
Chemicals and samples: 1 mM of HgCl2 suspension was prepared in 2% NaCl (pH7.5±0.02). Serial dilutions (1:1) for HgCl2 were directly prepared in 2% NaCl (pH7.5±0.02). Concentrations of HgCl2 tested were 13.55, 6.78, 3.39, 2, 1.70, 1.4, 1, 0.85, 0.5, 0.42, 0.21 and 0.11 mg L–1. The 2% NaCl (pH7.5±0.02) served as a control and diluent solution for all concentrations.
Luminometer: Luminescence of marine bacterium V. fischeri was performed using a 96-well microplate Luminoskan Ascent Luminometer (LAL) Thermo, manipulated by Ascent software version 2.6 ThermoLab systems. The bacteria growth was determined using spectrophotometer (UV-Visible) at the optical density of 600 nm (OD600 nm). The inhibition of luminescence intensity of V. fischeri caused by toxicants and was determined as follows:
![]()
![]()
| KF | = | Correction factor |
| ICn | = | Luminescence intensity of control after contact time (n time) (mV) |
| IC0 | = | Initial luminescence intensity of control sample (mV) |
| ITn | = | Luminescence intensity of test sample after contact time (n time) (mV) |
| IT0 | = | Initial luminescence intensity of the test sample (mV)14 |
Intensity and stability of luminescence of marine bacterium V. fischeri
Incubation of V. fischeri in LAL: Freeze-dried luminescent bacteria V. fischeri were stabilized first at 4°C for 30 min, then inoculated in 5 mL of a modified LBS medium and incubated at 25°C for 24 h. Cultures were harvested when they grew and 0.05 mL were inoculated into a fresh modified LBS medium (5 mL) and incubated at 25°C for 2 h. Prior to transfer bacterial suspension into microplates, 0.1 mL of 2% NaCl (pH7.5±0.02) was pipetted into well in duplicate and acclimated at 25°C for 30 min, which was supplemented with 0.1 mL of bacterial suspension by automatic dispensing in LAL testing chamber. The bioluminescence was recorded during the first 2 sec at 25°C after the microplate was automatically shacked, then incubated at the same temperature for 51 h. Each 60 min the bioluminescence was recorded again after shaking.
Incubation of V. fischeri with and without aeration: Luminescent bacteria V. fischeri were suspended in a modified LBS medium then aliquots of bacterial suspension (0.1 mL) were transferred into a fresh modified LBS medium (10 mL), incubated at 25°C and stirred (250 rpm). A series of samples were taken during incubations and immediately measured. The bioluminescence was measured by LAL and the bacteria growth by spectrophotometer at OD600 nm. For bioluminescence measurements, 0.1 mL of 2% NaCl (pH 7.5±0.02) was added into well duplicate and acclimated at 25°C for 30 min and then supplemented with 0.1 mL of bacterial suspension manually and measured. Prior each measurement, samples were automatically shaken for 10 sec in LAL. The same procedure was performed with aeration by supplementing air (600 mL min–1).
Assessment of HgCl2 toxicity: HgCl2 suspension of 1 mM was prepared containing 2% NaCl (pH7.5±0.02). Serial dilutions were prepared as previously described and diluents were placed at 25°C for 30 min 2% NaCl (pH7.5 ±0.02) served as a control. Prior to toxicity testing luminescent bacteria V. fischeri were reconstituted in a modified LBS medium, aliquots of bacterial suspension (0.1 mL) were transferred into a fresh modified LBS medium (10 mL), incubated at 25°C, stirred (250 rpm) and supplemented air (600 mL min–1) for 22 h 0.1 mL of each concentration was added into different wells in duplicate and was supplemented with 0.1 mL of bacterial suspension by automatic dispensing in the LAL testing chamber, followed by incubation at 25°C for 15 min. The bioluminescence was recorded after microplate was automatically shacked. The signal of the light was measured after 15 min in duplicate.
Volatilization of HgCl2 in fluidized bed reactor using immobilized and free bacteria E. coli
Stimulation of the bacteria: Fresh bacteria suspension of 5 mL E. coli were transferred in 500 mL of nutrient broth medium containing 1 mM of HgCl2 and incubated at 37°C with agitation (100 rpm) for 72 h, to induce the mercuric-reductase production. This enzyme reduce (Hg2+) to volatile mercury (Hg0). After 72 h, Cultures were centrifuged at 8600 rpm for 30 min using a centrifuge (SIGMA 2-16PK Heated and Refrigerated Centrifuge).
Fluidized bed reactor set: Two bottles flasks form the fluidized bed reactor, the first one contains 500 mL of minimal medium,10 g of glucose and 1 mM of HgCl2, was incubated at 37°C and inoculated by 0.4 g of E. coli. The mercury vapors obtained after mercury reduction by the bacteria were evacuated by current air using an air compressor to a mercury trap flask containing 500 mL of oxidizing solution potassium permanganate (KMnO4, 0.5 mM). The color of KMnO4 changed from purple to orange, this indicate successful detoxification. A series of samples were taken from both bottles during the procedure to evaluate the reduction of HgCl2. At different time samples were taken from bioreactor, centrifuged at 3000 rpm for 15 min and supernatant were measured using luminescence of V. fischeri to evaluate the reduction of HgCl2. Samples from the oxidizing solution were measured using Spectrophotometer at 530 nm (OD530 nm) to follow the discoloration of KMnO4. The same procedure was performed using immobilized bacteria E. coli until 116 h.
Immobilization of bacteria E. coli : The 100 mL sterile sodium alginate solution (2% w/v) was mixed with 0.832 g of bacteria biomass until homogenous. The mixture was extruded through a needle (2 mm i.d.) into 150 mM CaCl2 solution at room temperature for 30 min and rinsed with 50 mM Tris- HCl buffer pH 7.515.
Statistical analysis: All data were analyzed using GraphPad Prism (GraphPad Software version 6.0), values are means±standard errors. All experiments were done in triplicate.
RESULTS AND DISCUSSION
The aim of this study was to optimize the intensity and stability of luminescence emitted by bacteria V. fischeri cultivated in a modified LBS medium prior to assess the detoxification of HgCl2 which was detoxified by free and immobilized bacteria E. coli in fluidized bed reactor. The intensity of luminescence of V. fischeri while incubated in a water bath without aeration at 25°C and stirred (250 rpm) was less higher. Therefore, the intensity of luminescence of V. fischeri while incubated within LAL instrument was higher. Thereafter, the intensity of luminescence of V. fischeri while incubated in a water bath with aeration (600 mL m–1) at 25°C and stirred (250 rpm) was the highest one compared to other two cases. It was found that free bacteria E. coli detoxified more rapidly HgCl2 than the immobilized bacteria.
Intensity and stability for luminescence emission of marine bacterium V. fischeri : The best conditions for intensity and stability of luminescence emission and growth of the V. fischeri were determined in a modified LBS medium prior to assess HgCl2 toxicity. The modified medium used in this study was different from other LBS medium published16-22, thus 10 g of bacto tryptone was replaced by 10 g of nutrient broth. This modified medium provides also a high intensity of luminescence and strong growth as well as LBS medium does. During the incubation of V. fischeri in LAL instrument at 25°C for 51 h in aerobic conditions, it was found that V. fischeri started to emit light after 8 h of growth and the intensity of luminescence continued to rise (Fig. 1). The highest value (2540.2 mV) of the intensity of luminescence was at 34 h, then it decreased. During the incubation of V. fischeri in a water bath without aeration at 25°C and stirred (250 rpm). It was found that the intensity of luminescence rose with cell growth until it reached a peak (284.7 mV) at 50 h, then it dropped as the cell went into stationary phase (Fig. 2). During the incubation of V. fischeri in a water bath with aeration (600 mL min–1) at 25°C and stirred (250 rpm). It was found that the emission of luminescence started just 7 h of aeration. The intensity of luminescence rose with cell growth until it reached a peak (4372.5 mV) at 25 h, the corresponding cell growth was OD600 nm (1.8). Then the luminescence dropped as the cell went into stationary phase (Fig. 3). The intensity of luminescence of V. fischeri while incubated within LAL instrument was higher. Therefore, the intensity of luminescence of V. fischeri while incubated in a water bath without aeration at 25°C and stirred (250 rpm) was less higher.
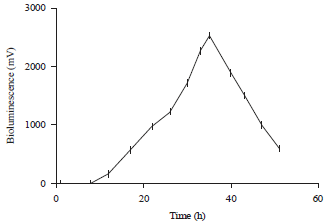 | |
| Fig. 1: | Luminescence intensity of V. fischeri versus time (h) during incubation within 96-well microplate LAL instrument |
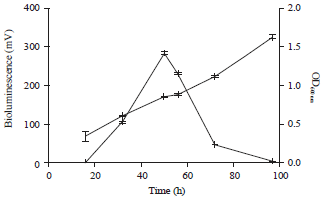 | |
| Fig. 2: | Growth and luminescence intensity of V. fischeri in a modified LBS medium incubated without aeration at 25°C and stirred (250 rpm) for 97 h |
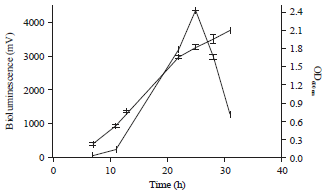 | |
| Fig. 3: | Growth and luminescence intensity of V. fischeri in a modified LBS medium incubated with aeration (600 mL min–1) at 25°C and stirred (250 rpm) for 30 h |
The intensity of luminescence of V. fischeri while incubated in a water bath with aeration (600 mL m–1) at 25°C and stirred (250 rpm) was the highest one compared to other two cases. It was found that the bacteria began to emit luminescence at almost same time with different intensity in two cases. Indeed the maximum intensity of luminescence during aeration was much higher (4372.5 mV) than that incubated in the instrument (2540.2 mV).
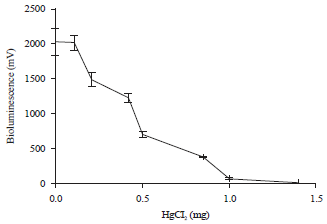 | |
| Fig. 4: | Measurement of dilutions of 1 mM HgCl2 using luminescence of V. fischeri which has grown in a modified LBS medium incubated in a water bath with aeration (600 mL min–1) at 25°C and stirred (250 rpm) |
| 2% NaCl (pH7.5 ±0.02) served as a control and diluent | |
In previous researches, the best conditions for intensity and stability of luminescence and growth were obtained when V. fischeri were grown in a media at pH (7.2-7.8)23-25, best temperatures between (20-25°C)26,27, strong aeration and stirring between (300-400 rpm)24. Therefore, in this study V. fischeri were cultivated in a modified LBS medium, containing 10 g nutrient broth, 5 g yeast extract, 2% NaCl, 50 mL 1 M Tris buffer, pH 7.5±0.02, temperate at 25°C, aerated (600 mL min–1) and stirred (250 rpm), was in accordance with previous researches. In this best conditions of luminescence emission, the intensity of luminescence increases with cell growth. Thus the best time of toxicity assessment by luminescence of V. fischeri is between 16-22 h.
Assessment of HgCl2 toxicity: V. fischeri were cultivated in a modified LBS medium and were successfully applied to assess the toxicity of HgCl2. Concentrations of HgCl2 tested were 13.55, 6.78, 3.39, 2, 1.70, 1.4, 1, 0.85, 0.5, 0 .42, 0.21 and 0.11 mg L–1. The 2% NaCl (pH7.5±0.02) served as a control and diluent solution for all concentrations. The control and the concentration (0.11 mg L–1) of HgCl2 were clearly similar which showed that this concentration was not sufficient to inhibit the light emitted by V. fischeri consequently it was not toxic to this strain. Therefore, six concentrations (13.55, 6.78, 3.39, 2, 1.70, 1.4 mg L–1) of HgCl2 were extremely toxic, similarly capable to inhibit the luminescence totally. The concentration 0.42 mg L–1 presented moderate toxic. Thereafter, inhibition of bioluminescence was closely related to concentrations of HgCl2. Greater concentration induce greater inhibition of luminescence, at the concentration 0.11 mg L–1 of HgCl2 there was no inhibition but from this concentration the light began to decrease, at the concentration 1.4 mg L–1 of HgCl2 inhibition of the light was almost total. The correlation between bioluminescence and concentrations of HgCl2 was very significant (0.99). Thus, bioluminescence was inversely proportional to the different concentrations of HgCl2 (Fig. 4).
Evaluation of volatilization of HgCl2 in fluidized bed reactor: A series of samples were taken from both bottles during fluidized bed reactor procedure using immobilized and free bacteria E. coli to evaluate the degradation of HgCl2. Samples from bottle one are centrifuged at 3000 rpm for 15 min and supernatants were measured using luminescence of V. fischeri to evaluate the degradation of HgCl2. Samples from bottle two were measured using a spectrophotometer at OD530 nm to evaluate the discoloration of KMnO4. The same procedure was performed using immobilized bacteria E. coli until 116 h. Free and immobilized bacteria E. coli have been used to detoxicate HgCl2, it was found that the bacteria began progressively to degrade HgCl2. In the case of free bacteria, 14 h and 30 min of the degradation, the quantity of HgCl2 remained was not enough to inhibit the luminescence of V. fischeri. Thus, E. coli reduced the quantity of HgCl2 from 13.55 mg L–1 to nearly 0.42 mg L–1. because 0.42 mg L–1 was moderately toxic to V. fischeri, thus it was not enough to inhibit the luminescence (Fig. 5a). In immobilized case, it was found that after 23 h and 30 min of detoxification, the quantity of HgCl2 remained was moderately toxic to V. fischeri, thus the complete detoxification was after 108 h and 30 min (Fig. 5b). The discoloration of KMnO4 was evaluated using spectrophotometer at OD530 nm, it was proportional in both cases. The color changed from purple to orange, this indicate successful detoxification. In the case of free bacteria, after 14 h and 30 min of detoxification OD530 nm was reduced from 0.78 to nearly 0.70. In the Immobilized case, after 23 h and 30 min of detoxificationOD530 nm was 0.68 (Fig. 6a and b). Therefore, the discoloration was perfectly proportional to the detoxification of HgCl2. Free E. coli bacteria is more rapid than immobilized one in detoxification of HgCl2. It was concluded that luminescence of V. fischeri which cultivated in a modified LBS medium is simple, rapid, sensitive and cost effective, can be used to evaluate toxicants.
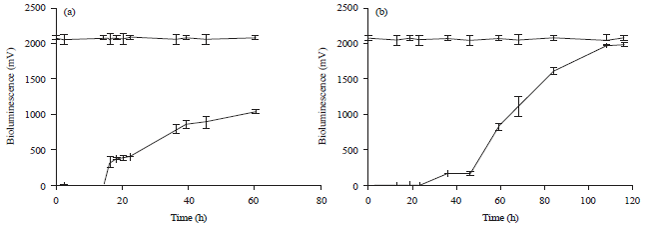 | |
| Fig. 5(a-b): | Kinetics of luminescence of V. fischeri exposed to suspensions of HgCl2 degraded by (a) free and (b) Immobilized bacteria E. coli, measured in a 96-well microplate LAL instrument. 2% NaCl served as control |
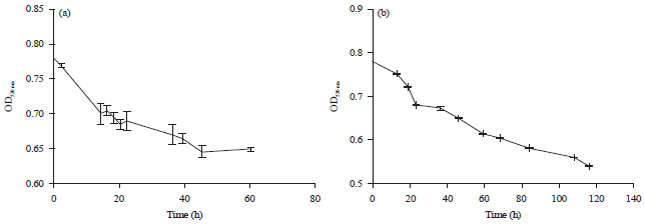 | |
| Fig. 6(a-b): | Curves (a) Free and (b) Immobilized bacteria E. coli represent discoloration of KMnO4 by gaseous monatomic mercury (Hg0). Measured using spectrophotometer at OD530 nm |
CONCLUSION
According to the different results obtained during this study, culture of V. fischeri in a modified LBS medium containing 10 g nutrient broth, 5 g yeast extract, 20 g NaCl, 50 mL 1 M Tris buffer, pH 7.5±0.02, temperated at 25°C, aerated (600 mL min–1) and stirred (250 rpm), was the best conditions for intensity and stability of luminescence emission. It was found that, the intensity of luminescence increases with cell growth but the bacteria must be at a certain critical threshold of its growth OD (1-1.8) beyond this threshold the intensity of light decreases. The best time of toxicity assessment by luminescence of V. fischeri is between 16-22 h of incubation. HgCl2 is extremely toxic to V. fischeri, indeed at a very low concentrations capable to inhibit totally the light emitted by this strain. Thus, inhibition of bioluminescence is closely related to concentrations of HgCl2. Greater concentration induce greater inhibition of light, at the concentration (0.11 mg L–1) of HgCl2 there was no inhibition of light but from this concentration the light began to decrease. The concentration (0.42 mg L–1) of HgCl2 was moderately inhibited the light. In the case of the concentration (1.4 mg L–1) of HgCl2, the inhibition light was total. The correlation between bioluminescence and concentrations of HgCl2 was very significant (0.99). Thus, bioluminescence is inversely proportional to the different concentrations of HgCl2. Thereafter, the discoloration is perfectly proportional to the detoxification of HgCl2. Free E. coli bacteria is more rapid than immobilized one in detoxification of HgCl2. It was concluded that luminescence of V. fischeri which cultivated in a modified LBS medium is a simple, rapid, sensitive and cost effective, can be used to evaluate toxicants.
SIGNIFICANCE STATEMENTS
This study discovers the possible utilization of bioluminescence of V. fischeri to assess the detoxification of HgCl2 that can be beneficial for toxicity evaluation of the metal. This study will help researchers to uncover the critical areas of toxicology and ecotoxicology. Thus, a new, simple, rapid, sensitive and cost effective method on the assessment of toxicants, may be arrived at.
ACKNOWLEDGMENT
Authors gratefully to Dr. Bouhilleoui Amina at Laboratory of Marine Biotoxins, National Institute of Halieutic Research, Casablanca, Morocco for providing bacteria V. fischeri.
REFERENCES
- Senthamilselvan, D., A. Chezhian, E. Suresh and R. Ezhilmathy, 2012. Toxic effects of heavy metals (cadmium plus mercury) on haematological parameters and DNA damage in Lates calcarifer. J. Toxicol. Environ. Health Sci., 4: 156-161.
Direct Link - Tchounwou, P.B., C.G. Yedjou, A.K. Patlolla and D.J. Sutton, 2012. Heavy Metal Toxicity and the Environment. In: Molecular, Clinical and Environmental Toxicology, Luch, A. (Ed.), Springer, Basel, Switzerland, ISBN: 978-3-7643-8339-8, pp: 133-164.
CrossRefDirect Link - Kim, Y.H., A.L. Khan, D.H. Kim, S.Y. Lee and K.M. Kim et al., 2014. Silicon mitigates heavy metal stress by regulating P-type heavy metal ATPases, Oryza sativa low silicon genes and endogenous phytohormones. BMC Plant Biol., Vol. 14.
CrossRefDirect Link - Uraguchi, S., T. Kamiya, T. Sakamoto, K. Kasai and Y. Sato et al., 2011. Low-affinity cation transporter (OsLCT1) regulates cadmium transport into rice grains. Proc. Nat. Acad. Sci., 108: 20959-20964.
CrossRefDirect Link - Satoh-Nagasawa, N., M. Mori, N. Nakazawa, T. Kawamoto and Y. Nagato et al., 2011. Mutations in rice (Oryza sativa) heavy metal ATPase 2 (OsHMA2) restrict the translocation of zinc and cadmium. Plant Cell Physiol., 53: 213-224.
CrossRefDirect Link - Ivask, A., I. Kurvet, K. Kasemets, I. Blinova and V. Aruoja et al., 2014. Size-dependent toxicity of silver nanoparticles to bacteria, yeast, algae, crustaceans and mammalian cells in vitro. Plos One, Vol. 9.
CrossRefDirect Link - Radzig, M.A., V.A. Nadtochenko, O.A. Koksharova, J. Kiwi, V.A. Lipasova and I.A. Khmel, 2013. Antibacterial effects of silver nanoparticles on gram-negative bacteria: influence on the growth and biofilms formation, mechanisms of action. Colloids Surf. B: Biointerfaces, 102: 300-306.
CrossRefDirect Link - Bacmaga, M., A. Borowik, J. Kucharski, M. Tomkiel and J. Wyszkowska, 2015. Microbial and enzymatic activity of soil contaminated with a mixture of diflufenican+ mesosulfuron-methyl+ iodosulfuron-methyl-sodium. Environ. Sci. Pollut. Res., 22: 643-656.
CrossRefDirect Link - Chingate, S., G. Delgado, L.M. Salazar and C.Y. Soto, 2015. The ATPase activity of the mycobacterial plasma membrane is inhibited by the LL37-analogous peptide LLAP. Peptides, 71: 222-228.
CrossRefDirect Link - Mortimer, M., K. Kasemets, M. Heinlaan, I. Kurvet and A. Kahru, 2008. High throughput kinetic Vibrio fischeri bioluminescence inhibition assay for study of toxic effects of nanoparticles. Toxicol. In vitro, 22: 1412-1417.
CrossRefDirect Link - Parvez, S., C. Venkataraman and S. Mukherji, 2006. A review on advantages of implementing luminescence inhibition test (Vibrio fischeri) for acute toxicity prediction of chemicals. Environ. Int., 32: 265-268.
CrossRefDirect Link - Jarque, S., P. Masner, J. Klanova, R. Prokes and L. Blaha, 2016. Bioluminescent Vibrio fischeri assays in the assessment of seasonal and spatial patterns in toxicity of contaminated river sediments. Front. Microbiol., Vol. 7.
CrossRefDirect Link - Visick, K.L., K.P. Quirke and S.M. McEwen, 2013. Arabinose induces pellicle formation by Vibrio fischeri. Applied Environ. Microbiol., 79: 2069-2080.
CrossRefDirect Link - Bose, J.L., C.S. Rosenberg and E.V. Stabb, 2008. Effects of luxCDABEG induction in Vibrio fischeri: Enhancement of symbiotic colonization and conditional attenuation of growth in culture. Arch. Microbiol., Vol. 19.
CrossRefDirect Link - Lyell, N.L. and E.V. Stabb, 2013. Symbiotic characterization of Vibrio fischeri ES114 mutants that display enhanced luminescence in culture. Applied Environ. Microbiol., 79: 2480-2483.
CrossRefDirect Link - Lyell, N.L., A.K. Dunn, J.L. Bose and E.V. Stabb, 2010. Bright mutants of Vibrio fischeri ES114 reveal conditions and regulators that control bioluminescence and expression of the lux operon. J. Bacteriol., 192: 5103-5114.
CrossRefDirect Link - Bose, J.L., U. Kim, W. Bartkowski, R.P. Gunsalus and A.M. Overley et al., 2007. Bioluminescence in Vibrio fischeri is controlled by the redox‐responsive regulator ArcA. Mol. Microbiol., 65: 538-553.
CrossRefDirect Link - Marsden, A.E., K. Grudzinski, J.M. Ondrey, C.R. DeLoney-Marino and K.L. Visick, 2017. Impact of salt and nutrient content on biofilm formation by Vibrio fischeri. Plos One, Vol. 12.
CrossRefDirect Link - Brooks, J.F., M.C. Gyllborg, A.A. Kocher, L.E. Markey and M.J. Mandel, 2015. TfoX-based genetic mapping identifies Vibrio fischeri strain-level differences and reveals a common lineage of laboratory strains. J. Bacteriol., 197: 1065-1074.
CrossRefDirect Link - Berglind, R., P. Leffler and M. Sjostrom, 2010. Interactions between pH, potassium, calcium, bromide and phenol and their effects on the bioluminescence of Vibrio fischeri. J. Toxicol. Environ. Health, Part A, 73: 1102-1112.
Direct Link - Scheerer, S., F. Gomez and D. Lloyd, 2006. Bioluminescence of Vibrio fischeri in continuous culture: Optimal conditions for stability and intensity of photoemission. J. Microbiol. Methods, 67: 321-329.
CrossRefDirect Link - Cheng, C.Y., J.T. Kuo, Y.C. Lin, Y.R. Liao and Y.R. Chung, 2010. Comparisons of Vibrio fischeri, Photobacterium phosphoreum and recombinant luminescent using Escherichia coli as BOD measurement. J. Environ. Sci. Health Part A, 45: 233-238.
CrossRefDirect Link - Alves, E., M.A. Faustino, J.P. Tome, M.G. Neves and A.C. Tome et al., 2011. Photodynamic antimicrobial chemotherapy in aquaculture: Photoinactivation studies of Vibrio fischeri. PLoS One, Vol. 6.
CrossRefDirect Link - Soto, W., J. Gutierrez, M.R. Remmenga and M.K. Nishiguchi, 2009. Salinity and temperature effects on physiological responses of Vibrio fischeri from diverse ecological niches. Microbiol. Ecol., 57: 140-150.
CrossRefDirect Link