
G. Sibi
Department of Biotechnology, Indian Academy Degree College, Centre for Research and Post Graduate Studies, Bengaluru, 560043, Karnataka, India
Research Journal of Environmental Toxicology
Year: 2015 | Volume: 9 | Issue: 4 | Page No.: 168-178







ABSTRACT
Autotrophic cultivation of microalgae induces low biomass and lipid production and is difficult to be used in pilot scale production. Heterotrophic and mixotrophic algae allows higher growth rate, which in turn can accumulate higher biomass and lipid. Enhanced biomass and lipid production is crucial to use oleaginous microalgae for industrial application of biodiesel production. A fresh water microalga was isolated from waste water treatment plant and identified as Scenedesmus obliquus. The isolate was grown under varying cultural and nutrient conditions to increase biomass and lipid production. For this purpose, CO2 (10-160 mL min–1), temperature (20-45°C), fructose, maltose, glucose, sucrose and starch (10-50 g L–1), urea, sodium nitrate, potassium nitrate, ammonium nitrate and ammonium chloride (0.2-1.0 g L–1) were used. Optimum conditions were found by calculating specific growth rate and used to determine their influence on biomass and lipid production. At 40 mL min–1 (0.51%) CO2 concentration, 18.17 g L–1 biomass and 64.2% lipid production was obtained from the isolate. Glucose at 20 g L‾1 have produced 18.62 g L–1 and 62.6% whereas sodium nitrate at 0.8 g L–1 resulted in 14.64 g L–1 and 65.1% of biomass and lipid production at the end of 20 days culturing period. Temperature of 25°C was found optimum where 12.38 g L–1 and 60.9% of biomass and lipid were obtained. Based on the results, by choosing optimum cultural conditions and nutrient composition of the growth medium it is possible to increase both biomass and lipid production in microalgae.
PDF Abstract XML References Citation
Received: June 02, 2015;
Accepted: July 08, 2015;
Published: July 31, 2015
How to cite this article
G. Sibi, 2015. Cultural Conditions and Nutrient Composition as Effective Inducers for Biomass and Lipid Production in Fresh Water Microalgae. Research Journal of Environmental Toxicology, 9: 168-178.
DOI: 10.3923/rjet.2015.168.178
URL: https://scialert.net/abstract/?doi=rjet.2015.168.178
DOI: 10.3923/rjet.2015.168.178
URL: https://scialert.net/abstract/?doi=rjet.2015.168.178
INTRODUCTION
Biofuel contributes no net carbon dioxide or sulfur to the atmosphere and emits less gaseous pollutants than fossil fuel (Antolin et al., 2002; Lang et al., 2001; Vicente et al., 2004). The interest in the production of microalgae as biofuels is increasing due to their high oil content, rapid biomass production and small foot print which is a promising alternative for fuel production. However, the production cost of fuel from microalgae is as high as fossil fuel-derived diesel. To solve this problem, many researchers are trying to lower the production costs by screening microalgae that have high lipid content and increase the lipid productivity through various environmental conditions. The ability of microalgae to survive or proliferate over a wide range of environmental conditions results in the production of an array of many secondary metabolites, which are of considerable value in biotechnology fields including aquaculture, health and food industries (Andersen, 1996). Heterotrophic cultivation of microalgae is far cheaper and easier than autotrophic culture which also can produce higher biomass and moderate lipid contents (Perez-Garcia et al., 2011; Fan et al., 2012).
The lipid content, composition and the proportions of various fatty acids of microalgae vary according to the environmental or culturing variables such as light intensity, growth phase photoperiod, temperature, salinity, CO2 concentration, nitrogen and phosphorous concentration (Dunstan et al., 1993; Zhu et al., 1997; Wu et al., 2011; Petkov and Garcia, 2007). Microalgae represent a significant source of valuable lipids and fatty acids when their fatty acid content is manipulated (Behrens and Kyle, 1996). Biomass and lipid production are the main parameters that determine the economic feasibility of biofuel production using microalgae. Commercialization of microalgal biofuel involving large-scale, outdoor open-ponds is still constrained by low lipid productivity (Sharma et al., 2012). Enhanced biomass and lipid production is crucial to use oleaginous microalgae for industrial application of biodiesel production. A recognized approach of enhancing the lipid production is varying cultural and nutrient conditions. Appropriate medium composition is of importance to achieve the best lipid production performance of microalgae species (Yeh and Chang, 2012). In this study, a fresh water microalga was isolated from urban waste water treatment plant followed by enhancing biomass and lipid production through varying cultural conditions as a means of microalgal biofuel source.
MATERIALS AND METHODS
Isolation and purification of the microalga: Waste water was collected from Bangalore Water Sewerage and Supply Board (BWSSB), Bengaluru (13°04’ N, 77°58’ E), India and poured into a closed 250 mL bottle and exposed in sunlight for 3 weeks. The upper layer of the water was inoculated into BG11 medium enriched agar plates containing 200 μg mL–1 ampicillin. The plates were incubated at 25±2°C under cool white fluorescent light (40 μmol photons m–2 sec–1; 15 h light/9 h dark) until algal growth was detected. Single green colour colonies were inoculated into BG11 medium and the algal growth was measured spectrophotometrically. Among the colonies, cultures shown maximum specific growth rate were selected and identified as Scenedesmus obliquus.
CO2 concentration: The effect of CO2 concentration on lipid composition and productivity was investigated by varying the CO2 concentration. By taking into account the CO2 content in air of about 0.039%, 1.5 mL min–1 CO2 was added under ambient air flow rate of 6 L min–1 initially. Experiments were conducted under the same air flow rate with the addition of 10, 20, 40, 80 and 160 mL min–1 pure CO2 gas (0.12, 0.26, 0.51, 1.02 and 2.05% CO2).
Nutrient composition: The isolate was grown under varying nutrient conditions to determine the effect of culture conditions on lipid composition and productivity. Algal cells in the stationary phase were inoculated into BG11 medium containing five carbohydrates (fructose, maltose, glucose, sucrose and starch) separately supplemented in the range of 10-50 g L–1. The initial concentrations of all carbohydrate sources were calculated as the same carbon atom number of 10 g L–1 glucose. Urea, sodium nitrate, potassium nitrate, ammonium nitrate and ammonium chloride were chosen (0.2-1.0 g L–1) as the nitrogen sources with the initial concentrations computed as the same nitrogen atom number of 0.6 g L–1 sodium nitrate.
Growth temperature: Varying temperatures between 20-45°C were used to culture the isolate to determine temperature influence on lipid composition and production. The algal cells were inoculated and cultured for a period of 30 days and the cells were harvested at stationary phase and analyzed for lipid production and composition.
Measurement of the growth: The initial algal concentration was 0.05 g L–1 during the study and all experiments were conducted in triplicates.
Statistical analysis: The results were expressed as means of the replicates along with Standard Deviation (±SD). The specific growth rate (μ) of microalgae was monitored by measuring the Optical Density (OD) at 680 nm using a microplate reader:
where, Ct are cell concentration at different time points (t1 and t2) respectively. The cultures were harvested by centrifugation at 8000×g for 10 min at late log phase. Pellets were then dried in the drying oven at 40°C for 48 h to determine biomass concentration.
Lipid extraction and estimation: Algal lipids were extracted according to the method of Folch et al. (1957). Briefly, the cells were centrifuged at 10000 rpm for 10 min and the pellet was homogenized with chloroform-methanol (2:1 v/v) solution. The sample was centrifuged and to the supernatant, 0.73% NaCl water was added to produce a final solvent system of 2:1:0.8 chloroform: methanol:water (v/v/v). The mixture was shaken for 5 min and centrifuged for 15 min at 2000 rpm to separate the phases. The lower organic phase was collected and the chloroform-methanol solution was evaporated under a steam of nitrogen. Aliquots of lipid extracts were mixed with 2 mL of dichromate solution and the tubes were placed in boiling water bath for 45 min. The tubes were shaken for 2 or 3 times during the heating and were cooled, removed 1 mL from each, diluted to 100 mL with water and the absorbance was read at 350 nm against water as blank. Palmitic acid was used as standard and the lipid content was determined from the standard curve.
RESULTS
Figure 1 shows the growth of algae under different CO2 concentration. At 20 days of culturing period, increasing CO2 concentration of 40 mL min–1 (0.51%) resulted in significant specific growth rate.
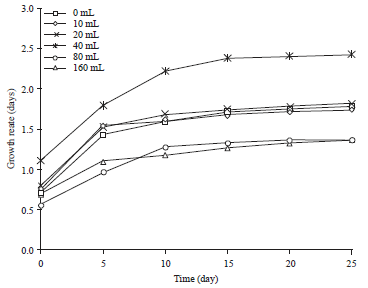 | |
| Fig. 1: | Specific growth rate at an air flow rate of 6 L min–1 with the addition of varying concentration of pure CO2 gas (A680) |
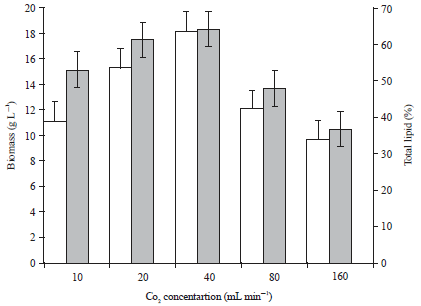 | |
| Fig. 2: | Biomass and lipid production under varying CO2 |
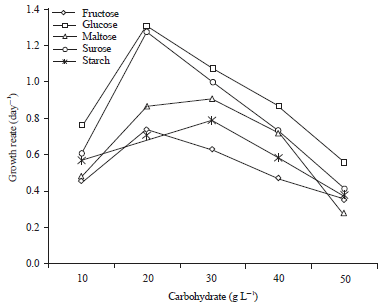 | |
| Fig. 3: | Specific growth rate under different concentration of carbohydrates (A680) |
However increasing CO2 flow rate above 40 mL min–1 has reduced the specific growth rate of microalgae. Biomass of Scenedesmus was highest at 40 mL min–1, which also resulted in higher lipid accumulation (64.1%) (Fig. 2).
Three monosaccharides (fructose, glucose and maltose), one each of disaccharide (sucrose) and polysaccharide (starch) were used at concentrations of 10-50 g L–1 used to determine the influence on specific growth rate for a period of 20 days. The isolate was able to utilize the carbohydrates but resulted in varying growth rate at the end of culturing period. Growth rate was enhanced significantly from initial concentration to 20 g L–1 and was subsequently reduced with increasing concentrations of carbohydrates (Fig. 3). Among the carbon sources, glucose have enhanced significant specific growth rate (1.31 day–1) followed by sucrose (1.28 day–1) at 20 g L–1.
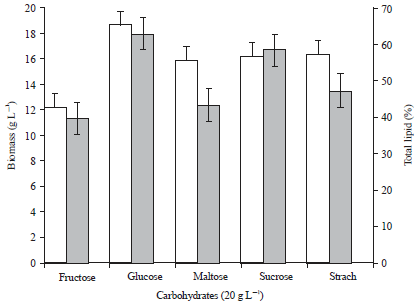 | |
| Fig. 4: | Biomass concentration and total lipid production under varying carbohydrates |
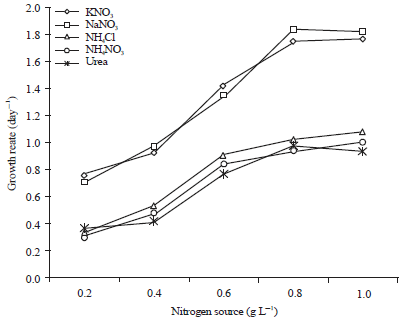 | |
| Fig. 5: | Specific growth rate under different concentration of nitrogen sources (A680) |
At highest concentration (50 g L–1), the growth rate in the presence of glucose and sucrose were 0.56 and 0.41 day–1. Among the carbohydrates, maltose had poor influence on specific growth rate of the isolate. Based on the higher specific growth rate, 20 g L–1 was chosen as optimum concentration to determine the influence on biomass and lipid production. Glucose and sucrose have recorded highest biomass (18.62 and 16.21 g L–1) and lipid production (62.6 and 58.3%) at 20 g L–1 concentration (Fig. 4).
To determine an optimal nitrogen source for algal growth and lipid accumulation, potassium nitrate, sodium nitrate, ammonium chloride, ammonium nitrate and urea were added separately (0.2-1.0 g L–1) to BG 11 medium. As depicted in Fig. 5, presence of ammonium chloride, ammonium nitrate and urea resulted in lower specific growth rate whereas, sodium nitrate and potassium nitrate has significantly increased the specific growth rate after 20 days culturing period.
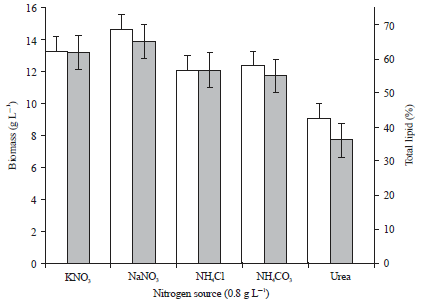 | |
| Fig. 6: | Biomass concentration and total lipid production under varying nitrogen sources |
The specific growth rate was significantly increased up to 0.8 g L–1 in the presence of sodium nitrate (1.84 day–1) and potassium nitrate (1.75 day–1). Whereas, it was increased beyond 0.8 g L–1 when the medium was added with ammonium chloride and ammonium nitrate but the specific growth rate was lesser than other nitrogen sources. Urea has exhibited poor growth throughout the study irrespective of the concentration. The results suggested that 0.8 g L–1 could satisfy the growth requirement of the isolate and was used to determine the biomass and lipid production. The biomass with sodium nitrate was significantly highest (14.64 g L–1) and the second highest was observed in the presence of potassium nitrate (13.27 g L–1). The same trend was reflected in lipid accumulation with 65.1 and 61.8% for sodium nitrate and potassium nitrate, respectively (Fig. 6). Lowest lipid accumulation was observed in the presence of urea (36.2%) and the results revealed that sodium nitrate and potassium nitrate were the suitable nitrogen sources for biomass and lipid accumulation in Scenedesmus sp. under the investigated conditions.
The specific growth rate of Scenedesmus under different cultivation temperature is shown in Fig. 7. The specific growth rate was significant at 20-35°C and was less at 40°C, resulted in decay after 25 days. The biomass concentration at 25°C and 30°C was higher (12.38 and 11.33 g L–1) than other temperatures (Fig. 8). Both biomass and lipid accumulation were decreased with increasing temperature but at 30°C, lipid content was increased though there was a lower biomass concentration.
DISCUSSION
Microalgae are photosynthetic microorganisms able to use solar energy to combine water with carbon dioxide to create biomass. CO2 is known to influence the lipid content of algae (Negoro et al., 1991; Watanabe et al., 1992; Kodama et al., 1993) and alterations in the composition of the fatty acids are dependent on the CO2 concentration during the algal growth (Tsuzuki et al., 1990; Yusof et al., 2011). This study used varying concentrations of CO2 and 40 mL min–1 (0.51%) has recorded maximum specific growth rate, biomass and lipid production. Enhanced lipid production in cell at various CO2 concentrations was also reported (Chiu et al., 2009; Ho et al., 2010).
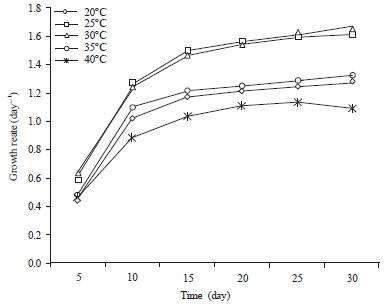 | |
| Fig. 7: | Specific growth rate under varying temperature (A680) |
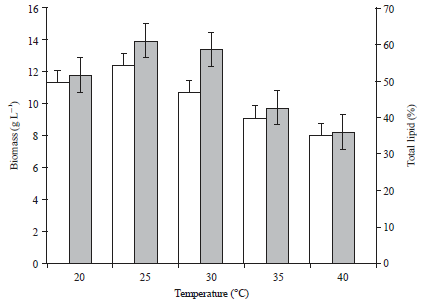 | |
| Fig. 8: | Biomass concentration and total lipid production under varying temperature |
Riebesell et al. (2000) reported CO2 concentration of up to 1% of air will increase the lipid production in algae and in this study both biomass and lipid accumulation was increased up to 0.51% and declined at higher CO2 percent in air. The specific growth rate was significant with increased CO2 levels and above 40 mL min–1 reduced growth was observed. At highest CO2 levels (160 mL min–1, 2.05%), reduction rate of 46.7 and 42.6% were observed for biomass and lipid production, respectively. This could be due to low pH as a result of higher CO2 concentration based on fact that higher CO2 concentrations, unutilized CO2 will be converted to bicarbonate (Hailing-Sorensen et al., 1996) thereby, decreasing the pH of growth medium.
Change in macronutrients of growing environment will result in the change of cellular macromolecular composition of microalgae. Different carbohydrates were used to cultivate the algae during which glucose and sucrose were found to enhance the growth rate, biomass and lipid production in Scenedesmus. Glucose is the most commonly used carbon source for heterotrophic cultures of many microalgae (Perez-Garcia et al., 2011; Griffiths et al., 1960) and it has produced highest biomass concentration in green algae (Gim et al., 2014). It was noted that higher concentrations of carbohydrates resulted in inhibition of algal growth and 20 g L–1 was found as optimum for maximum specific growth rate. In this study, 18.62 g L–1 biomass was obtained in the presence of glucose followed by sucrose (16.21 g L–1) at 20 g L–1. Cheirsilp and Torpee (2012) reported glucose has increased the algal growth rather than lipid production and in the present work, both biomass and lipid production were increased. It was also found that starch has increased the biomass production (16.33 g L–1) but failed to increase the lipid production. On the other hand, fructose recorded the lowest biomass (12.21 g L–1) and lipid (39.4%) production among the carbohydrates used. The growth and lipid accumulation of microalgae are affected by nutrition concentration of the growth medium (Richardson et al., 1969; Khozin-Goldberg and Cohen, 2006; Hu et al., 2008; Li et al., 2010; Pruvost et al., 2011). Nitrogen affects the biomass growth and lipid productivity of various microalgae (Griffiths and Harrison, 2009). In this study, nutrient composition of the medium was modified by supplementing different nitrogen sources at different concentrations. Microalgae grew best at limited concentration of nitrogen which triggers high amount of carbohydrate and lipid (Suen et al., 1987). Highest specific growth rate was observed in the presence of sodium nitrate and potassium nitrate at 0.8 g L–1. Similar nitrogen sources have increased biomass production in Scenedesmus (Arumugam et al., 2013), however urea exhibited less influence on biomass (9.11 g L–1) and lipid (36.2%) production in this study. Potassium nitrate and sodium nitrate were reported to increase the biomass concentration of other algal species (Li et al., 2008; Dayananda et al., 2005) and the present work observed increased lipid production along with biomass of Scenedesmus. In a study by Gonzalex-Garcinuno et al. (2014), ammonium nitrate was produced maximum lipid yield by up to 3.55 mg day–1 in Scenedesmus abundans whereas, sodium nitrate yielded highest lipid production in this study (65.1%).
Changes in culture conditions divert the biosynthetic metabolism to lipid synthesis, thus obtaining fats as the main constituent instead of proteins. Among the varying culture conditions, temperature affects the lipid accumulation in microalgal cells. Microalgae respond to decreased growth temperature by increasing the ratio of unsaturated to saturated fatty acids and highest total lipid was observed at lower temperatures (Bohnenberger and Crossetti, 2014; Cao et al., 2014; Hoffmann et al., 2010). In this study, 15.2% increase in lipid production was observed at 25°C when compared to 20°C, which is in accordance to Converti et al. (2009), where increase of the cultivation temperature from 20-25°C has increased the lipid content. At the same time, further increase in cultivating temperature up to 40°C resulted in lower lipid production at a decreased rate of 40.9% when compared to 25°C. However, higher temperature has increased the biomass concentration and lipid productivity (Yang et al., 2013; Wu et al., 2013; Subhash et al., 2014). Another observation in the study was at 20°C, biomass and lipid production were 11.33 g L–1 and 51.6% whereas, it was 10.69 g L–1 and 58.6% at 30°C revealing that lipid production is not dependent on biomass concentration.
The isolate Scenedesmus was grown under heterotrophic conditions and the results revealed the optimum CO2 (40 mL min–1), carbohydrate (20 g L–1), nitrogen (0.8 g L–1) and temperature (25°C) conditions. Varying specific growth rate was observed during culturing in which glucose and sodium nitrate were found as suitable carbon and nitrogen sources with higher biomass and lipid production. In general, lipid production was directly proportional to biomass concentration biomass production. By choosing optimum cultural conditions and nutrient composition of the growth medium it is possible to increase both biomass and lipid production in microalgae.
REFERENCES
- Antolin, G., F.V. Tinaut, Y. Briceno, V. Castano, C. Perez and A.I. Ramırez, 2002. Optimisation of biodiesel production by sunflower oil transesterification. Bioresour. Technol., 83: 111-114.
CrossRefDirect Link - Arumugam, M., A. Agarwal, M.C. Arya and Z. Ahmed, 2013. Influence of nitrogen sources on biomass productivity of microalgae Scenedesmus bijugatus. Bioresour. Technol., 131: 246-249.
CrossRefDirect Link - Behrens, P.W. and D.J. Kyle, 1996. Microalgae as a source of fatty acids. J. Food Lipids, 3: 259-272.
CrossRefDirect Link - Bohnenberger, J.E. and L.O. Crossetti, 2014. Influence of temperature and nutrient content on lipid production in freshwater microalgae cultures. Anais Academia Brasileira Ciencias, 86: 1239-1248.
CrossRefDirect Link - Cao, J., H.L. Yuan, B.Z. Li and J.S. Yang, 2014. Significance evaluation of the effects of environmental factors on the lipid accumulation of Chlorella minutissima UTEX 2341 under low-nutrition heterotrophic condition. Bioresour. Technol., 152: 177-184.
CrossRefDirect Link - Cheirsilp, B. and S. Torpee, 2012. Enhanced growth and lipid production of microalgae under mixotrophic culture condition: Effect of light intensity, glucose concentration and fed-batch cultivation. Bioresour. Technol., 110: 510-516.
CrossRefDirect Link - Chiu, S.Y., C.Y. Kao, M.T. Tsai, S.C. Ong, C.H. Chen and. C.S. Lin, 2009. Lipid accumulation and CO2 utilization of Nannochloropsis oculata in response to CO2 aeration. Bioresour. Technol., 100: 833-838.
CrossRefPubMedDirect Link - Converti, A., A.A. Casazza, E.Y. Ortiz, P. Perego and M. del Borghi, 2009. Effect of temperature and nitrogen concentration on the growth and lipid content of Nannochloropsis oculata and Chlorella vulgaris for biodiesel production. Chem. Eng. Process.: Process Intensif., 48: 1146-1151.
CrossRefDirect Link - Dunstan, G.A., J.K. Volkman, S.M. Barrett and C.D. Garland, 1993. Changes in the lipid composition and maximisation of the polyunsaturated fatty acid content of three microalgae grown in mass culture. J. Appllied Phycol., 5: 71-83.
CrossRefDirect Link - Dayananda, C., R. Sarada, S. Bhattacharya and G.A. Ravishankar, 2005. Effect of media and culture conditions on growth and hydrocarbon production by Botryococcus braunii. Process Biochem., 40: 3125-3131.
CrossRefDirect Link - Fan, J., J. Huang, Y. Li, F. Han and J. Wang et al., 2012. Sequential heterotrophy-dilution-photoinduction cultivation for efficient microalgal biomass and lipid production. Bioresour. Technol., 112: 206-211.
CrossRefDirect Link - Folch, J., M. Lees and G.H.S. Stanley, 1957. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem., 226: 497-509.
CrossRefPubMedDirect Link - Gim, G.H., J.K. Kim, H.S. Kim, M.N. Kathiravan, H. Yang, S.H. Jeong and S.W. Kim, 2014. Comparison of biomass production and total lipid content of freshwater green microalgae cultivated under various culture conditions. Biosyst. Eng., 37: 99-106.
CrossRefDirect Link - Gonzalex-Garcinuno, A., A. Tabernero, J.M. Sanchez-Alvarez, E.M.M. del Valle and M.A. Galan, 2014. Effect of nitrogen source on growth and lipid accumulation in Scenedesmus abundans and Chlorella ellipsoidea. Bioresour. Technol., 173: 334-341.
CrossRefDirect Link - Griffiths, D.J., C.L. Thresher and H.E. Street, 1960. The heterotrophic nutrition of Chlorella vulgaris (Brannon No. 1 strain): With two figures in the text. Ann. Bot., 24: 1-11.
Direct Link - Griffiths, M.J. and S.T.L. Harrison, 2009. Lipid productivity as a key characteristic for choosing algal species for biodiesel production. J. Applied Phycol., 21: 493-507.
CrossRefDirect Link - Ho, S.H., W.M. Chen and J.S. Chang, 2010. Scenedesmus obliquus CNW-N as a potential candidate for CO2 mitigation and biodiesel production. Bioresour. Technol., 101: 8725-8730.
CrossRefDirect Link - Hoffmann, M., K. Marxen, R. Schulz and K.H. Vanselow, 2010. TFA and EPA productivities of Nannochloropsis salina influenced by temperature and nitrate stimuli in turbidostatic controlled experiments. Mar. Drugs, 8: 2526-2545.
CrossRefDirect Link - Hu, Q., M. Sommerfeld, E. Jarvis, M. Ghirardi, M. Posewitz, M. Seibert and A. Darzins, 2008. Microalgal triacylglycerols as feedstocks for biofuel production: Perspectives and advances. Plant J., 54: 621-639.
CrossRefPubMedDirect Link - Khozin-Goldberg, I. and Z. Cohen, 2006. The effect of phosphate starvation on the lipid and fatty acid composition of the fresh water eustigmatophyte Monodus subterraneus. Phytochemistry, 67: 696-701.
CrossRefDirect Link - Lang, X., A.K. Dalai, N.N. Bakhshi, M.J. Reaney and P.B. Hertz, 2001. Preparation and characterization of bio-diesels from various bio-oils. Bioresour. Technol., 80: 53-62.
CrossRefDirect Link - Li, X., H.Y. Hu, G. Ke and Y.X. Sun, 2010. Effects of different nitrogen and phosphorus concentrations on the growth, nutrient uptake and lipid accumulation of a freshwater microalga Scenedesmus sp. Bioresour. Technol., 101: 5494-5500.
CrossRefDirect Link - Li, Y., M. Horsman, B. Wang, N. Wu and C.Q. Lan, 2008. Effects of nitrogen sources on cell growth and lipid accumulation of green alga Neochloris oleoabundans. Applied Microbiol. Biotechnol., 81: 629-636.
CrossRefDirect Link - Negoro, M., N. Shioji, K. Miyamoto and Y. Miura, 1991. Growth of microalgae in high CO2 gas and effects of SOx and NOx. Applied Biochem. Biotechnol., 28-29: 877-886.
CrossRefPubMedDirect Link - Perez-Garcia, O., F.M.E. Escalante, L.E. de-Bashan and Y. Bashan, 2011. Heterotrophic cultures of microalgae: Metabolism and potential products. Water Res., 45: 11-36.
CrossRefPubMedDirect Link - Petkov, G. and G. Garcia, 2007. Which are fatty acids of the green alga Chlorella? Biochem. Syst. Ecol., 35: 281-285.
CrossRefDirect Link - Pruvost, J., G. Van Vooren, B. Le Gouic, A. Couzinet-Mossion and J. Legrand, 2011. Systematic investigation of biomass and lipid productivity by microalgae in photobioreactors for biodiesel application. Bioresour. Technol., 102: 150-158.
CrossRefDirect Link - Richardson, B., D.M. Orcutt, H.A. Schwertner, C.L. Martinez and H.E. Wickline, 1969. Effects of nitrogen limitation on the growth and composition of unicellular algae in continuous culture. Applied Environ. Microbiol., 18: 245-250.
Direct Link - Riebesell, U., A.T. Revill, D.G. Holdsworth and J.K. Volkman, 2000. The effects of varying CO2 concentration on lipid composition and carbon isotope fractionation in Emiliania huxleyi. Geochimica Cosmochimica Acta, 64: 4179-4192.
CrossRefDirect Link - Sharma, K.K., H. Schuhmann and P.M. Schenk, 2012. High lipid induction in microalgae for biodiesel production. Energies, 5: 1532-1553.
CrossRefDirect Link - Hailing-Sorensen, B., N. Nyhohn and A. Baun, 1996. Algal toxicity tests with volatile and hazardous compounds in air-tight test flasks with CO2 enriched headspace. Chemosphere, 32: 1513-1526.
CrossRefDirect Link - Subhash, G.V., M.V. Rohit, M.P. Devi, Y.V. Swamy and S.V. Mohan, 2014. Temperature induced stress influence on biodiesel productivity during mixotrophic microalgae cultivation with wastewater. Bioresour. Technol., 169: 789-793.
CrossRefDirect Link - Suen, Y., J.S. Hubbard, G. Holzer and T.G. Tornabene, 1987. Total lipid production of the green alga Nannochloropsis sp. QII under different nitrogen regimes. J. Phycol., 23: 289-296.
CrossRefDirect Link - Tsuzuki, M., E. Ohnuma, N. Sato, T. Takaku and A. Kawaguchi, 1990. Effects of CO2 concentration during growth on fatty acid composition in microalgae. Plant Physiol., 93: 851-856.
CrossRefDirect Link - Vicente, G., M. Martinez and J. Aracil, 2004. Integrated biodiesel production: A comparison of different homogeneous catalysts systems. Bioresour. Technol., 92: 297-305.
CrossRefDirect Link - Watanabe, Y., N. Ohmura and H. Saiki, 1992. Isolation and determination of cultural characteristics of microalgae which functions under CO2 enriched atmosphere. Energy Convers. Manage., 33: 545-552.
CrossRefDirect Link - Wu, H., J.V. Volponi, A.E. Oliver, A.N. Parikh, B.A. Simmons and S. Singh, 2011. In vivo lipidomics using single-cell Raman spectroscopy. Proc. Natl. Acad. Sci. USA., 108: 3809-3814.
CrossRefDirect Link - Wu, L.F., P.C. Chen and C.M. Lee, 2013. The effects of nitrogen sources and temperature on cell growth and lipid accumulation of microalgae. Int. Biodeterior. Biodegrad., 85: 506-510.
CrossRefDirect Link - Yang, Y., B. Mininberg, A. Tarbet and P. Weathers, 2013. At high temperature lipid production in Ettlia oleoabundans occurs before nitrate depletion. Applied Microbiol. Biotechnol., 97: 2263-2273.
CrossRefDirect Link - Yeh, K.L. and J.S. Chang, 2012. Effects of cultivation conditions and media composition on cell growth and lipid productivity of indigenous microalga Chlorella vulgaris ESP-31. Bioresour. Technol., 105: 120-127.
CrossRefDirect Link - Yusof, Y.A.M., J.M.H. Basari, N.A. Mukti, R. Sabuddin and A.R. Muda et al., 2011. Fatty acids composition of microalgae Chlorella vulgaris can be modulated by varying carbon dioxide concentration in outdoor culture. Afr. J. Biotechnol., 10: 13536-13542.
Direct Link - Zhu, C.J., Y.K. Lee and T.M. Chao, 1997. Effects of temperature and growth phase on lipid and biochemical composition of Isochrysis galbana TK1. J. Applied Phycol., 9: 451-457.
CrossRefDirect Link