
Tarek M. Heikal
Environmental Toxicology Research Unit (ETRU), Department of Pesticide Chemistry, National Research Centre (NRC), Cairo, Egypt
Abdel-Tawab H. Mossa
Environmental Toxicology Research Unit (ETRU), Department of Pesticide Chemistry, National Research Centre (NRC), Cairo, Egypt
Azza W. Ibrahim
Environmental Toxicology Research Unit (ETRU), Department of Pesticide Chemistry, National Research Centre (NRC), Cairo, Egypt
Hala F. Abdel-Hamid
Environmental Toxicology Research Unit (ETRU), Department of Pesticide Chemistry, National Research Centre (NRC), Cairo, Egypt
Research Journal of Environmental Toxicology
Year: 2014 | Volume: 8 | Issue: 2 | Page No.: 53-67







ABSTRACT
Pesticides have contributed for many public health hazards in man including infertility. So, the present study aimed to assess the protective role of green tea extract (GT) against the possibility of reproductive toxicity resulting from chlorpyrifos (CPF), cyromazine (Cyr) and their combination exposure in mature male Wistar rats. Rats were administered CPF (5.4 mg kg-1 b.wt., 1/25 LD50), Cyr (135.48 mg kg-1 b.wt., 1/25 LD50), CPF+Cyr, GT (1.5% w/v in water) as the only drinking fluid, CPF+GT, Cyr+GT and CPF+Cyr+GT daily via gavage for 70 days to complete the spermatogenic cycle. The results revealed that exposure to CPF, Cyr and CPF+Cyr significantly decreased the fertility index, weights of sexual organs (testes, seminal vesicles, epidermis and prostate gland), sperm characteristics (motility and count) as well as serum testosterone level, while increased sperm abnormality. In addition, the testicular tissue level of reduced glutathione (GSH) and the activities of superoxide dismutase (SOD), catalase (CAT), glutathione peroxidase (GPx) and glutathione-S-transferase (GST) enzymes were significantly decreased while increased the level of testicular tissue lipid peroxidation (LPO) compared with the control group. The testicular histopathological lesions were characterized by moderate to severe degenerative changes of seminiferous tubules and incomplete arrest of spermatogenesis. Co-administration of GT to treated-animals alleviates the reproductive toxicity and testicular oxidative damage. In conclusion, the use of green tea extract appeared to be beneficial in attenuating and improving the testicular damage and reproductive toxicity sustained by insecticide exposure in male rats.
PDF Abstract XML References Citation
Received: December 12, 2013;
Accepted: February 21, 2014;
Published: May 02, 2014
How to cite this article
Tarek M. Heikal, Abdel-Tawab H. Mossa, Azza W. Ibrahim and Hala F. Abdel-Hamid, 2014. Oxidative Damage and Reproductive Toxicity Associated with Cyromazine and
Chlorpyrifos in Male Rats: The Protective Effects of Green Tea Extract. Research Journal of Environmental Toxicology, 8: 53-67.
DOI: 10.3923/rjet.2014.53.67
URL: https://scialert.net/abstract/?doi=rjet.2014.53.67
DOI: 10.3923/rjet.2014.53.67
URL: https://scialert.net/abstract/?doi=rjet.2014.53.67
INTRODUCTION
Pesticides have brought the green revolution in the world and are being widely used to control agricultural pests and insects causing public health hazards including infertility. The infertility rate in humans has increased tremendously in the past few decades (Oehninger, 2001; Venkatesh et al., 2009). The decline in sperm counts by about 50% may be the main cause of the infertility (Carlsen et al., 1992). Exposure to chemical agents including pesticides has contributed to this decline (Cox, 1994). Owing to the extensive use of organophosphate pesticides in agriculture, there is a high risk of human exposure to these chemicals (Sarkar et al., 2000).
In fact, pesticides are known to increase the production of Reactive Oxygen Species (ROS), which in turn generate oxidative stress in different tissues (Rai and Sharma, 2007; Heikal et al., 2011). Many studies have implicated oxidative damage as the central mechanism of toxicity (Halliwell and Gutteridge, 2002; Heikal et al., 2013). Oxidative damage primarily occurs through production of Reactive Oxygen Species (ROS) that are generated during the reaction and react with biological molecules, eventually damaging membranes and other tissues (Kalender et al., 2010; Heikal et al., 2012). Many insecticides are hydrophobic molecules that bind extensively to biological membranes, especially phospholipids bilayers (Ogutcu et al., 2008) and they may damage membranes by inducing lipid peroxidation (LPO) (Kalender et al., 2010; Mossa et al., 2012; Heikal et al., 2013).
Chlorpyrifos (CPF), a broad spectrum organophosphate insecticide, is one of the most extensively used organophosphate insecticides (OPIs) in agriculture, domestic and industrial applications all over the world (Joshi et al., 2007; Mossa et al., 2012; Heikal et al., 2013). CPF is thought to be primarily metabolized in the liver involving the intervention of multiple, specific cytochrome P450’s through several reaction pathways (Mutch and Williams, 2006). Like other OPIs, CPF poisoning is primarily through the inhibition of acetylcholinesterase activity in target tissues, resulting in excessive accumulation of acetylcholine (ACh) at the cholinergic receptors in the peripheral and central nervous systems (Ogutcu et al., 2008; Heikal et al., 2013). Toxicity occurs at doses that do not inhibit AChE (Slotkin, 2005). Therefore, other mechanisms have been implicated in OP toxicity such as induction of oxidative stress leading to generation of free radicals and a decrease in antioxidants enzymes or oxygen-free-radical scavenging systems (Sharma et al., 2005; Heikal et al., 2012). However, there is a positive correlation between the level of 3, 5, 6-trichloropyridinol, a metabolite of CPF and low testosterone and sperm count (Meeker et al., 2006).
Cyromazine (Cyr), N-cyclopropyl-1, 3, 5-triazine-2, 4, 6-triamine, is an effective systemic insecticide and an insect growth regulator that acts by inhibiting the moulting processes. It is wildly used as an agricultural insecticide and a feed additive to animal breeding against fly larvae from hatching in manure (Graf, 1993).
As mechanism of pesticides toxicity often involves oxidative stress, numerous efforts were done to identify dietary compounds able to strengthen the cellular antioxidant defense so as to counteract the oxidative stress. In this respect, herbal medicines derived from plant extracts are being increasingly utilized to treat a wide variety of clinical disease. More attention has been paid to the protective effects of natural antioxidants against chemically induced toxicities (Frei and Higdon, 2003). The increasing interest in the health properties of green tea and its main catechin polyphenols have led to a significant rise in scientific investigation for prevention and therapeutics in several diseases (Mandel et al., 2006; Heikal et al., 2013). Crespy and Williamson reported that green tea extract (GT) displays antioxidants and free radicals scavenger properties (Crespy and Williamson, 2004; Heikal et al., 2011). Owing to the scarce in literature related to the oxidative damage and reproductive toxicity resulting from Cyr, CPF and their combination exposure, the present study aimed to describe the protective effects of green tea extract against cyromazine and chlorpyrifos induced reproductive toxicity and oxidative stress in rat testis.
MATERIALS AND METHODS
Animals: Mature male albino rats of Wistar strain (Rattus norvegicus) weighing 160±10 g (4-5 months old) were obtained from the Animal Breeding House of the National Research Centre (NRC), Dokki, Giza, Egypt and maintained in clean plastic cages in the laboratory animal room (23±2°C). On standard pellet diet, tap water ad libitum and daily dark/light cycle (12/12 h), the rats were acclimatized for 1 week prior to the start of experiments. The experimental work on rats was performed with the approval of the Animal Care and Experimental Committee, National Research Centre, Cairo, Egypt and international guidelines for care and use of laboratory animals.
Chemicals: Pu-erh green tea of post-fermented tea produced in Yunnan province, China. Chloropyrifos (97%) and Cyromazine (99%) were obtained from TaeGeuk Cop., South Korea. Thiobarbituric acid, H2O2, S-2, 4-dinitrophenyl glutathione, 5,50- dithiobis-(2-nitrobenzoic acid), phosphoric acid, butanol, sodium phosphate, sodium carbonate, sodium azide, EDTA, Tris-HCl, epinephrine ware brought from Sigma, St. Louis, USA. Kit of GSH was obtained from Biodiagnostic for diagnostic reagents; Dokki, Giza, Egypt. All other chemicals were of reagent grades and were obtained from the local scientific distributors in Egypt.
Preparation of green tea extract: Likewise, the crude aqueous extract of green tea was prepared according to Maity et al. (1998) and later adopted by El-Beshbishy (2005) by soaking 15 g of instant green tea leaves in 1 L of distilled water whose temperature did not exceed 90°C, for 5 min to obtain soluble polyphenols dissolved in the aqueous extract. The solution was filtered to obtain the final 1.5% (w/v) green tea extract. This solution was substituted in the place of water as the sole source of drinking fluid.
Experimental design: Rats were randomly divided into 8 groups each containing 20 animals. The route of administration selected for the study was oral gavage for 70 consecutive days to complete the spermatogenic cycle and maturation of sperms in epididymis (Sarkar et al., 2000). Rats in group 1 served as control and were given corn oil (4 mL kg-1) and allowed distillate water ad libitum. Rats in group 2 were allowed aqueous green tea extract as the sole drinking fluid during the experimental period at a concentration of 1.5% (w/v). Rats in group 3 were daily given cyromazine (Cyr) in corn oil at a dose of 135.48 mg kg-1 b.wt. (1/25 LD50) (Tomlin, 2004). Rats in group 4 were daily given chlorpyrifos (CPF) in corn oil at a dose of 5.4 mg kg-1 b.wt. (1/25 LD50) (Tomlin, 2004). Rats in group 5 were given a combination of CPF and Cyr. Rats in group 6, 7 and 8 were given the same doses of pesticides as in groups 3, 4 and 5, respectively and simultaneously allowed to an aqueous green tea extract as the sole source of drinking fluid.
After completion of treatment period, blood samples were collected for estimating total testosterone in serum. The rats were sacrificed and the testes, seminal vesicle and prostate glands were removed and weighed. Semen samples were collected from cuda epididymis by cutting the tail of epididymis and squeezing it gently on clean slide. The semen was used for estimating the epididymal sperm characters (motility and count) according to the method adopted by Bearden and Fuquay (1980). The testes were preserved in 10% neutral formalin solution till processed for histopathological examination. The selected dose of the CPF was based on previous studies (McCollister et al., 1974).
Mating and fertility indexes: After the end of the treatment course, males of control and experimental groups of treated rats (n = 20/group), were mated 1:1 with untreated proven fertile, with regular estrus cycle, females for 5 days (complete one estrous cycle) (Nunez et al., 1996). Mating was confirmed by the presence of vaginal plugs or deposition of spermatozoan at the vaginal orifice upon vaginal examination. The day that a vaginal plug was found was considered day 0 of gestation. Then mating and fertility indexes were estimated and recorded.
Preparation of homogenates: The excised testicular tissue was washed with distal water for the removal of blood and later the fatty parts were removed. Tissues were homogenized in ice-cold 50 mM sodium phosphate buffer (pH 7.4) containing 0.1 mM ethylenediaminetetraacetic acid (EDTA), using Potter-Elvehejem homogenizer. The homogenate was centrifuged at 3000xg at 4°C for 15 min to remove cell debris and the supernatant was saved in aliquots and stored at -20°C for assaying protein concentration, lipid peroxidation (LPO) and antioxidant enzymes activity.
Oxidative stress evaluation: Lipid peroxidation (LPO) level: Lipid peroxidation process is determined in supernatant of testicular tissue homogenate by the thiobarbituric acid (TBA) method which estimates the malondialdehyde formation (MDA) according to Esterbauer and Cheeseman (1990). Two hundred fifty microliters of tissue homogenate were added to 1.5 mL of 1% phosphoric acid (pH 2.0) and 1 mL of 0.6% of TBA in air-light tubes and were placed in a boiling water bath for 25 min. After incubation, the sample was cooled to room temperature and MDA-TBA was extracted with 2.5 mL of butanol. Organic phase was separated by centrifugation for 5 min at 2000xg and measured at 532 nm. The concentration of MDA was calculated by the absorbance coefficient of MDA-TBA complex (1.56x105 M-1 cm-1). Lipid peroxidation is expressed as nmoles MDA/mg protein.
Reduced glutathione content (GSH) of supernatant estimation was performed by the method of Beutler et al. (1963) using commercial glutathione reduced kits (Biodiagnostic for diagnostic reagents: Dokki, Giza, Egypt). Determination of GSH is based on the reaction of DTNB [5, 5-dithiobis-(2-nitrobenzoic acid)] with GSH and yield a yellow colored chromophore with a maximum absorbance at 412 nm. The amount of GSH present in the testicular tissue was calculated as nmoles g-1 tissue.
The enzyme catalase (CAT) converts H2O2 into water. In brief, 0.25 g of tissue was homogenated in 1 mL of 50 mM Tris-HCl and centrifuged at 2000xg for 15 min. Then 10 μL of supernatant was added to a quartz cuvette containing 980 μL of distilled water and 10 μL of 0.066 M H2O2 (dissolved in sodium phosphate buffer) was added to start the reaction. The testicular CAT activity was measured spectrophotometrically at 240 nm by calculating the rate of degradation of H2O2, the substrate of the enzyme (Xu et al., 1997). Activity of CAT is expressed as units mg-1 protein.
The specific activity of testicular superoxide dismutase (SOD) was determined according to the method described by Misra and Fridovich (1972). Ten micro liters of tissue homogenate were added to 970 μL of EDTA-sodium carbonate buffer (0.05 M) at pH 10.2. The reaction was started by adding 20 μL of epinephrine (30 mM) and the activity was measured at 480 nm for 4 min. A unit of SOD is defined as the amount of enzyme that inhibits by 50% the speed of oxidation of epinephrine and the results were expressed as U mg-1 protein.
Glutathione peroxidase (GPx) catalyzes the reduction of hydroperoxides by utilizing GSH as a reluctant. Determination of testicular GPx activity was carried out according to the method of Chiu et al. (1976). The reaction mixture contained 0.5 mL of 0.4 M sodium phosphate buffer (pH 7.0) and 0.4 mM EDTA, supplemented with 0.25 mL of sodium azide (1 mM), 0.5 mL of GSH (2 mM) and 0.25 mL of D.W and 0.5 mL of homogenate was added and allowed to equilibrate for 5 min at 37°C. The reaction was initiated by adding 0.5 mL of H2O2 (1.25 mM). Absorbance at 340 nm was recorded at 1, 3 and 6 min. The activity of this enzyme was estimated by measurement of the residual reduced glutathione remaining after the action of the enzyme with the Ellman’s reagent (DTNB) in the presence of cumene hydroperoxide as a secondary substrate. Specific activity of this enzyme is expressed as U min-1 mg-1 protein.
Glutathione-S-transferase (GST) activity of testicular was measured spectrophotometrically by the method of Habig et al. (1974) using 1-chloro-2,4-dinitrobenzene as electrophilic substrate that binds to GSH with the participation of the enzyme and forms a colored GSH-substrate complex, detected at 340 nm. The activity of GST was expressed in terms of μmol min-1 mg-1 protein.
Protein concentration: The total protein level was determined according the method described by Lowry et al. (1951) using Bovine Serum Albumin (BSA) as a standard (Lowry et al., 1951).
Serum testosterone concentration: Serum samples of the treated male rats were used for estimating testosterone concentration using radio immunoassay (RIA) method (kit catalog #1119). This method is based on the competitive binding principal where the unknown or standards samples were incubated with radioactive iodine125 labeled testosterone in antibody-coated tubes. After incubation, the liquid contents in the tubes were withdrawn and the bound radioactivity was determined using gamma counter according to method described by Wilke and Utley (1987).
Histopathological examination: For light microscopic investigations, specimens from testes were fixed in 10% phosphate buffer formalin, dehydrated in alcohols and embedded in paraffin. Five micron tissue sections were stained with hematoxylin and eosin stain (H and E) for general histopathological examination. Scoring of histopathological changes was done as follow: (-) absent, (+) mild, (++) moderate, (+++) severe and (++++) extremely severe (Bancroft et al., 1996).
Statistical analysis: The results were expressed as Means±SE. All data were done with the Statistical Package for Social Sciences (SPSS 11.0 for windows). The results were analyzed using one way analysis of variance (ANOVA) followed by Duncan’s test for comparison between different treatment groups. Statistical significance was set at p<0.05.
RESULTS
Effect on general health of rats: During the experiment, no death was observed in any of the experimental groups. Rats in the control group and in green tea extract (GT) treated group did not show any sign of toxicity. However, CPF and Cyr+CPF treated rats showed varying degrees of clinical signs few minutes after dosing. The signs included huddling, mild tremor and diarrhea. The observed signs were related to the cholinergic crisis; a consistent sign in organophosphate poisoning. Except for the huddling, no other significant clinical manifestation was observed following GT supplementation.
Effect on sexual organs weights and sperm characteristics: As recorded in Table 1 oral administration of Cyr, CPF and their combination for 70 successive days caused significant decreases in the weights of testes, seminal vesicles and prostate glands, as compared to the normal control group. In addition, spermatozoa count and motility reduced significantly in all treated groups, as compared to the normal control group. Interestingly, these adverse effects of insecticides administrations were much alleviated in animals treated with the GT extract.
Effect on mating and fertility indices: Fertility indexes of the male rats given Cyr, CPF and their combination for 70 consecutive days were 57.1, 58.35 and 55.5%, respectively compared to 100% in the control normal group.
| Table 1: | Effect of oral administration of cyromazine, chlorpyrifos and their combination at doses 1/25 LD50 in the absence and presence of green tea extract for 70 days on the weight of sexual organs and sperm characteristics in male rats |
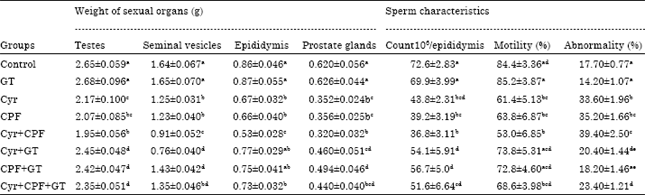 | |
| Each value is a mean of 6 rats±SE, values are not sharing superscripts letters, a, b, c, d, eSignificantly diffrent at p<0.05, Cyr: Cyromazine, CPF: Chlorpyrifos, GT: Green tea extract | |
| Table 2: | Functional fertility parameters of male rats after oral administration of cyromazine, chlorpyrifos and their combination at doses 1/25 LD50 in absence and presence of green tea extract for 70 days |
 | |
| Mating index (%): No. of males inseminated females/total No. of males cohabited with femalesx100,Fertility index (%): No. of cohabited females becoming pregnant/No. of non pregnant with evidence of vaginal plugx100,Cyr: Cyromazine, CPF: Chlorpyrifos, GT: Green tea extract | |
Rats given GT extracts have fertility index of 100% while they were 70.6, 72.4 and 66.6% when co-administered with Cyr, CPF and their combination, respectively (Table 2).
Effect on LPO level in testicular tissue: Administration of Cyr, CPF and Cyr+CPF resulted in a significant increase (p<0.05) in the levels of malondialdehyde (MDA) by +45, +57 and +93%, respectively as compared to the control group. When GT extract co-administrated with Cyr, CPF and Cyr+CPF, it has capable of recovering the activities of MDA level to the normal values (Fig. 1).
Effect on GSH level in testicular tissue: The result of testicular reduced glutathione (GSH) level is presented in Fig. 2. These results clearly indicated that treatment with Cyr, CPF and Cyr+CPF resulted in a significant decrease in the level of testes GSH as compared to control animals. While, male rats treated with GT extract showed non significant increase in testes GSH content as compared to control rats. However, coadministration of GT extract to the treated groups attenuated the level of GSH to the normal values.
Effect on activities of SOD, CAT, GPx and GST in testicular tissue: Present results revealed that Cyr, CPF and their combination produced a statistically significant decrease (p<0.05) in SOD activity in male rats (Table 3) compared to the control value. Administration of GT extract to treated groups of male rats improved the levels of SOD towards the control values although this treatment could not normalize it.
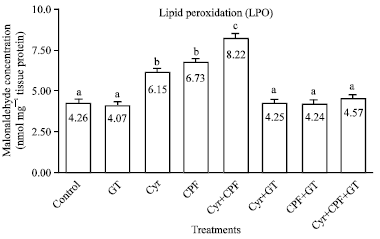 | |
| Fig. 1: | Effect of GT extract on testicular malonaldehyde (MDA) values as index of lipid peroxidation, after 70 day of treatment in control, Cyr-treated rats (135.5 mg kg-1 b.wt), CPF-treated rats (5.4 mg kg-1 b.wt) and Cyr+CPF-treated rats. Data are expressed as mean±SEM of 6 rats. Columns are not sharing above letters (a-e) differ significantly at p<0.05, Cyr: Cyromazine, CPF: Chlorpyrifos, GT: Green tea extract |
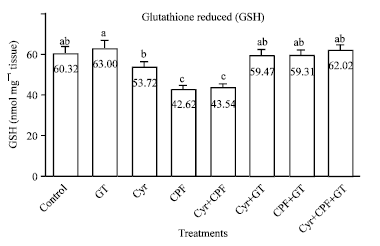 | |
| Fig. 2: | Effect of GT extract on testicular reduced glutathione (GSH) values after 70 days of treatment in control, Cyr-treated rats (135.5 mg kg-1 b.wt), CPF-treated rats (5.4 mg kg-1 b.wt) and Cyr+CPF-treated rats. Data is expressed as Mean±SEM of 6 rats. Columns are not sharing above letters (a-e) differ significantly at p<0.05, Cyr: Cyromazine, CPF: Chlorpyrifos, GT: Green tea extract |
Treatment with GT extract alone did not result in significant alteration in SOD activity compared to control treatment.
The result clearly indicated that treatment with Cyr, CPF and their combination resulted in a significant decrease in the activities of testes CAT, GPx and GST as compared to control animals. However, male rats treated with GT extract showed non significant increase in GPx, GST and CAT as compared to control rats. When GT extract administrated with Cyr, it has capable of recovering the activities of them to the normal values (Table 3).
| Table 3: | Effect of green tea consumption on testicular antioxidant enzymes of male rats treated with cyromazine, chlorpyrifos and their combination |
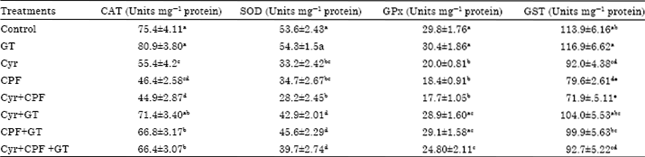 | |
| Each value is a mean of 6 rats±SEM, a,b,c,dValues are not sharing superscripts letters, a,b,c,dSignificantly deffrent at p<0.05, Cyr: Cyromazine, CPF: Chlorpyrifos, GT: Green tea extract | |
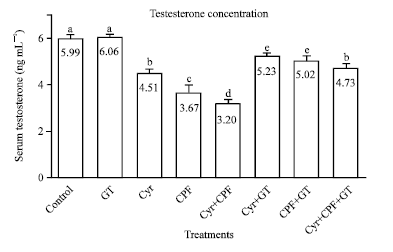 | |
| Fig. 3: | Effect of GT extract on serum testosterone levels after 70 days of treatment in control, Cyr-treated rats (135.5 mg kg-1 b.wt.), CPF-treated rats (5.4 mg kg-1 b.wt.) and Cyr+CPF-treated rats. Data is expressed as Mean±SEM of 6 rats. Columns are not sharing above letters (a-e) differ significantly at p<0.05, Cyr: Cyromazine, CPF: Chlorpyrifos, GT: Green tea extract |
Effect on serum testosterone hormone concentration: Data showed that oral administration of Cyr, CPF and their combination for 70 consecutive days induced a significant (p<0.05) decrease in serum testosterone levels as compared to the control normal group (Fig. 3). GT extract administration has no effect on serum testosterone level. However, co administration of GT to the insecticidal treatments improved the levels of testosterone towards the control values although it could not normalize them (Fig. 3).
Histopathological findings: The representative pictures of histopathological examination in testes tissue are shown in Fig. 4a-f and the semiquantitative histological scoring of liver damage is presented in Table 4. Histopathological examination of the testes of normal rats revealed mature functioning seminiferous tubules associated with complete spermatogenial cell series (Fig. 4a). The testes of rats given GT extract alone showed also normal histological structure of the seminiferous tubules (Fig. 4b).
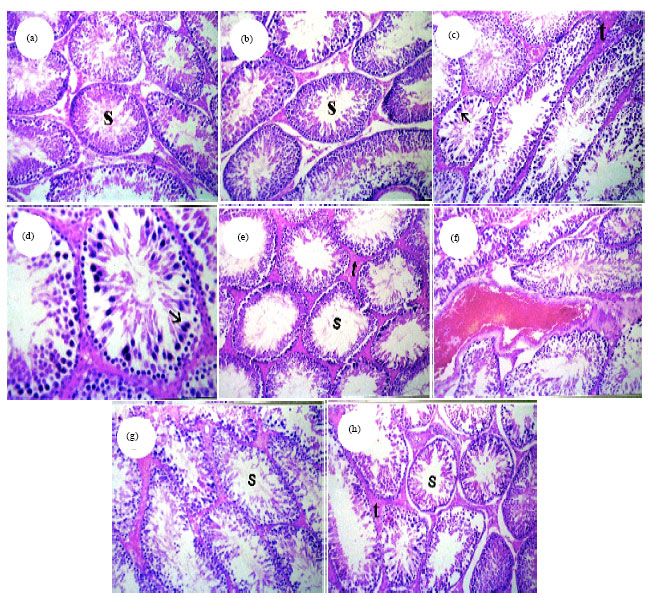 | |
| Fig. 4(a-h): | Testes paraffin sections stained by haematoxylin and eosin (H and E) for histopathological changes after 70 days administration of Cyr, CPF and their combination with and without GT. Control and GT groups (a-b) Normal histological structure of mature seminiferous tubules with complete spermatogenic series (s) (x 40), CPF-treated group, (c) Sertoli cells proliferation in some seminiferous tubules (black arrow) and homogenous eosinophilic materials in between seminiferous tubules replacing the interstitial cell (t) (x 40), (d) Magnification of Fig. 4c, to identify the proliferated sertoli cells in seminiferous tubules (s) (x 80), Cyr-treated group, (e) Homogenous eosinophilic materials in between seminiferous tubules replacing the interstitial cell (t) with absence of spermatozoa in most of the tubular lumen (s) (x 40), Cyr+CPF-treated group, (f) Sever congestion of intertubular blood vessel (x 40), (g) Absence of sperms in some of the seminiferous tubules lumens (s) (H and E x40), Cyr+CPF+GT-treated group, (h) Moderate amount of homogenous eosinophilic materials in between seminiferous tubules replacing the interstitial cell (t) and recovery of the spermatogenesis (s) (x 40). Cyr: Cyromazine, CPF: Chlorpyrifos, GT: Green tea extract |
| Table 4: | Severity of the reaction in testicular tissue according to the histopathological findings |
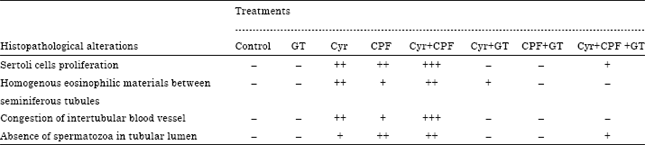 | |
| Cyr: Cyromazine, CPF: Chlorpyrifos, GT: Green tea extract. –: Normal, +: Mild, ++: Moderate, +++: Severe and ++++: Extremely severe | |
The examined testes of rats given CPF at 1/25 of the LD50 showed sertoli cells proliferation in some seminiferous tubules and homogenous eosinophilic materials in betweenseminiferous tubules replacing the interstitial cell (Fig. 4c, d). The examined testes of rats given Cyr at 1/25 of the LD50 showed homogenous eosinophilic materials in between seminiferous tubules replacing the interstitial cell with absence of spermatozoa in most of the tubular lumen (Fig. 4e). The examined testes of rats given Cyr+CPF showed sever congestion of intertubular blood vessel as well as absence of spermatozoa in most of the tubular lumen (Fig. 4g). Testes of rats given Cyr, CPF, their combination and GT extract acid showed moderate amount of homogenous eosinophilic materials in between seminiferous tubules replacing the interstitial cell and recovery of the spermatogenesis (Fig. 4h). Whereas, the severity of the above cited histological abnormalities were ranged from nil to moderate degree and represented in Table 4.
DISCUSSION
The reproductive toxicity of Cyr, CPF and their combination in male rats was manifested by lowered fertility index, decreased weight of the testes, seminal vesicles and prostate glands and lowered semen quantity and quality. These findings were in agreement with those of Joshi et al. (2007) who reported reduction in the weight of testes, hormonal changes and testicular damage after chronic exposure of male rats to CPF and other insecticides (Pant et al., 1996; Berger et al., 2000). The reduction in the testicular weight reflects deleterious changes in seminiferous tubules (Joshi et al., 2007). Since sperm motility is an important functional measurement to predict sperm fertilizing capacity. So, any negative impact on motility would seriously affect fertilizing ability. In this respect, marked inhibition of sperm motility in Cyr, CPF and Cyr+CPF-treated groups may be because of low level of ATP content (Bai and Shi, 2002). Full ATP pool is crucial for normal spermatozoa movement and a slight deprivation of ATP leads to reduction in motility, which may cause infertility. Sperm count is considered to be one of the important factors that affect fertility (Bebb et al., 1996). Suppression of gonadotrophins might have caused decrease in sperm density in testes (Joshi et al., 2007). Also, toxicants have direct effect on sertoli cell function, which appears to be involved in the control of spermiation and when disturbed caused epithelial disorganization, impaired spermatogenesis and subsequent tubular atrophy (Bedwal et al., 1994). The negative fertility test may be attributed to lack of forward progression and reduction in density of spermatozoa and altered biochemical milieu of caudal epididymis. The present study considers the first study that used GT against the reproductive toxicity of CPF and/or Cyr. The male rats that were given combination of GT and Cyr, CPF and their combination have fertility index 70.6, 72.4 and 66.6% while the fertility index in the rat treated with Cyr, CPF and their combination only were 57.1, 58.3 and 55.5%, respectively as compared to control group.
In the present findings, reduction in the serum testosterone, clearly demonstrated the inhibitory effect of insecticides on the secretion of pituitary gonadotrophins (FSH and LH) and in turn on the testosterone biosynthesis in the testes of rat (Singh and Pandey, 1990). The decrease in testosterone production may be induced by the stimulation of P450 aromatase (P450 arom), which catalyzes estrogen production from androgen; thereby decreasing androgen levels (Saitoh et al., 2001). These results suggest that CPF and/or Cyr exert suppressive effects on testicular function and leads to infertility in rats which in turn could be enhanced by using GT extract.
The decrease in serum testosterone levels, relative testes and epididymis weights observed here confirms earlier results of (Grote et al., 2004) in rats, (Sarpa et al., 2007) in mice and in rabbits (Yousef et al., 2010). The present study declared that CPF and/or Cyr caused a decrease in epididymal sperm count and sperm viability of rat. These results could be suggested that CPF and/or Cyr impair male reproduction in rat by decreasing circulatory testosterone. The observed decrease in sperm motility could be attributed in part to the concomitant abnormality of the sperms and a decrease in their viability. However, overproduction of ROS can be detrimental to sperm as it is may be associated with male infertility (Akiyama, 1999).
Oxidative stress refers to disrupted redox equilibrium between the production of free radicals and the ability of cells to protect against damage caused by these species. Defense against oxidative stress are maintained by using several mechanisms which include antioxidant machinery (Bergamini et al., 2004). The main cellular components susceptible to damage by free radicals are lipids (peroxidation of unsaturated fatty acids in cell membrane); this in turn can impair cellular structure and function (Bergamini et al., 2004). It has been indicated that the LPO is one of the molecular mechanisms involved in pesticide-induced cytotoxicity (Abdollahi et al., 2004; Heikal et al., 2013). A significant increase in the LPO level was observed in the present study. These results are in line with the observations of previous researchers following OP insecticides administration (Mansour and Mossa, 2010; Heikal et al., 2011, 2013).
The results of the current study revealed that green tea extract (GT) reversed the elevation of lipid peroxidation. Hence, it is possible that the mechanism of green tea extract may be attributed to epicatechins (antioxidant present in green tea) that scavenge a wide range of free radicals including the most active hydroxyl radical, which may initiate lipid peroxidation. Moreover, it was reported previously that it chelates metal ions, especially iron and copper, which, in turn inhibit generation of hydroxyl radicals and degradation of lipid hydroperoxides (Azam et al., 2004).
Reproductive toxicity could also be explained by the impaired antioxidant enzyme activities in the testes of the rat. The current data displayed Cyr and/or CPF-induced reduction in the activities of the antioxidant enzymes (SOD, CAT, GPx and GST) and the content of non-enzymatic antioxidant (GSH). These enzymes work together to eliminate active oxygen species. In this respect, SOD accelerates the dismutation of superoxide radicals (O2¯) into molecular oxygen (O2) and hydrogen peroxide (H2O2) (Gupta, 2006). H2O2 is neutralized by the combined action of CAT and GPx in all vertebrates (Salvi et al., 2007). This inhibition may be due to the decreased synthesis of enzymes or oxidative inactivation of enzyme protein.
Glutathione peroxidases are antioxidant selenoenzymes that are present in the cytosol of cells or plasma; the kidney secretes GPx into plasma. The major function of these enzymes, which use glutathione as a substrate, is to reduce soluble hydrogen peroxide and alkyl peroxidases (Demir et al., 2011). GPx converts hydrogen peroxide into H2O in the presence of oxidated glutathione (Kanbur et al., 2009). Considering that glutathione-S-transferases (GST) are detoxifying enzymes that catalyze the conjugation of a variety of electrophilic substrates to thiol g roup of GSH, producing less toxic forms (Mossa et al., 2012; Heikal et al., 2012). OP pesticides have been reported to significantly inhibit GPx and GST activities in various rat tissues (Abdollahi et al., 2004; Demir et al., 2011; Heikal et al., 2013). In the present study, the decreased GPx and GST activities might reflect cellular oxidative stress due to pesticides exposure.
As regards to the histopathological results, the testicular degenerative changes induced by CPF and/or Cyr exposure in intoxicated rats, as demonstrated in this study, agree with many previous investigators who reported variable degrees of degenerative changes and accumulation of cellular debris in the seminiferous tubules up to total cellular destruction after exposure of male rats to different insecticides (Pant et al., 1996; Mahgoub and El-Medany, 2001). The testicular damage induced by CPF and/or Cyr in this study confirms the reported lowered fertilizing capacity of the treated rats. The toxic effect on male reproductive system of the rat could be possibly explained by its direct cytotoxic effect and/or indirectly via decreased serum testosterone level.
Most of the biochemical alterations accompanied by histopathological changes were alleviated following GT administration. This could be attributed to the antioxidant capacity of GT that reduce the lipid peroxidation which in turn restore the integrity of the cell membrane and improve the disturbance in permeability.
Since the oxidative damage as the central mechanism of pesticides toxicity occurs primarily through production of Reactive Oxygen Species (ROS), the use of antioxidants to counteract the formed ROS is the corner stone in alleviating such hazards. So, the major nutraceutical compounds in green tea are tea catechins that have the most effective antioxidant activity. Tea catechins are an efficient free radical scavenger due to their one electron reduction potential (Higdon and Frei, 2003; Dubick and Omaye, 2007). In addition, tea contains minerals that function as co-factors in antioxidant enzymes: zinc, selenium and manganese. Polyphenols have additional mechanisms in which they reduce oxidation level besides direct role as antioxidants: (1) Binding of metal ions such as iron and copper and preventing their participation in oxidation reactions (leading to the formation of hydroxyl radical), (2) Prevention of redox sensitive transcription factors activation that amongst others things serve as mediators of inflammatory reactions, (3) Suppression of oxidation stimulants such as induced nitric oxide synthase (iNOS), cyclooxygenase 2 (COX-2), lipoxygenase 2 (LOX-2) and xanthine oxidase and (4) Induction of antioxidant enzymes such as glutathione S-transferase and super oxide dismutase (Cabrera et al., 2006).
CONCLUSION
This study revealed that chlorpyrifos and cyromazine insecticides induce reproductive toxicity in male rats manifested by decreases in the fertility index, weights of the sexual organs, semen characteristics and serum testosterone level as well as testicular damage manifested by induction of lipid peroxidation and depletion of antioxidant enzymes in testes of rat. However, the ultimately effects was observed in their combination. In contrast, co-administration of GT extract with the insecticides antagonizes their reproductive toxicity and oxidative damage. Based on our present observations, we propose that GT may provide a cushion for prolonged therapeutic option against toxins-induced reproductive toxicity and oxidative damage without harmful side effects.
REFERENCES
- Abdollahi, M., A. Ranjbar, S. Shadnia, S. Nikfar and A. Rezaie, 2004. Pesticides and oxidative stress: A review. Med. Sci. Monit., 10: RA141-RA147.
PubMedDirect Link - Akiyama, M., 1999. [In vivo scavenging effect of ethylcysteine on reactive oxygen species in human semen]. Nippon Hinyokika Gakkai Zasshi, 90: 421-428, (In Japanese).
PubMedDirect Link - Azam, S., N. Hadi, N.U. Khan and S.M. Hadi, 2004. Prooxidant property of green tea polyphenols epicatechin and epigallocatechin-3-gallate: Implications for anticancer properties. Toxicol. In Vitro, 18: 555-561.
CrossRef - Bai, J.P. and Y.L. Shi, 2002. Inhibition of Ca2+ channels in mouse spermatogenic cells by male antifertility compounds from Tripterygium wilfordii Hook. f. Contraception, 65: 441-445.
CrossRef - Bedwal, R.S., M.S. Edwards, M. Katoch, A. Bahuguna and R. Dewan, 1994. Histological and biochemical changes in testis of zinc deficient BALB/c strain of mice. Indian J. Exp. Biol., 32: 243-247.
PubMed - Bergamini, C.M., S. Gambetti, A. Dondi and C. Cervellati, 2004. Oxygen, reactive oxygen species and tissue damage. Curr. Pharm. Des., 10: 1611-1626.
PubMedDirect Link - Berger, T., M.G. Miller and C.M. Horner, 2000. In vitro fertilization after in vivo treatment of rats with three reproductive toxicants. Reprod. Toxicol., 14: 45-53.
CrossRef - Bebb, R.A., B.D. Anawalt, R.B. Christensen, C.A. Paulsen, W.J. Bremner and A.M. Matsumoto, 1996. Combined administration of levonorgestrel and testosterone induces more rapid and effective suppression of spermatogenesis than testosterone alone: A promising male contraceptive approach. J. Clin. Endocrinol. Metab., 81: 757-762.
CrossRefDirect Link - Beutler, E., O. Duron and B.M. Kelly, 1963. Improved method for the determination of blood glutathione. J. Lab. Clin. Med., 61: 882-888.
PubMedDirect Link - Cabrera, C., R. Artacho and R. Gimenez, 2006. Beneficial effects of green tea-A review. J. Am. Coll. Nutr., 25: 79-99.
PubMedDirect Link - Carlsen, E., A. Giwercman, N. Keiding and N.E. Skakkebaek, 1992. Evidence for decreasing quality of semen during past 50 years. Br. Med. J., 305: 609-613.
PubMedDirect Link - Chiu, D.T.Y., F.H. Stults and A.L. Tappel, 1976. Purification and properties of rat lung soluble glutathione peroxidase. Biochem. Biophys. Acta Enzymol., 445: 558-566.
CrossRefDirect Link - Crespy, V. and G. Williamson, 2004. A review of the health effects of green tea catechins in in vivo animal models. J. Nutr., 134: 3431S-3440S.
PubMedDirect Link - Demir, F., F.G. Uzun, D. Durak and Y. Kalender, 2011. Subacute chlorpyrifos-induced oxidative stress in rat erythrocytes and the protective effects of catechin and quercetin. Pesticide Biochem. Physiol., 99: 77-81.
CrossRef - El-Beshbishy, H.A., 2005. Hepatoprotective effect of green tea (Camellia sinensis) extract against tamoxifen-induced liver injury in rats. J. Biochem. Mol. Biol., 38: 563-570.
CrossRefPubMedDirect Link - Esterbauer, H. and K.H. Cheeseman, 1990. Determination of Aldehydic Lipid Peroxidation Products: Malonaldehyde and 4-hydroxynonenal. In: Methods in Enzymology, Shukla, A.K. (Ed.), Academic Press, Cambridge, Massachusetts, ISBN: 9780121820879, pp: 407-421.
CrossRefDirect Link - Frei, B. and J.V. Higdon, 2003. Antioxidant activity of tea polyphenols in vivo: Evidence from animal studies. J. Nutr., 133: 3275S-3284S.
PubMedDirect Link - Graf, J.F., 1993. The role of insect growth regulators in arthropod control. Parasitol. Today, 9: 471-474.
CrossRefDirect Link - Grote, K., B. Stahlschmidt, C.E. Talsness, C. Gericke, K.E. Appel and I. Chahoud, 2004. Effects of organotin compounds on pubertal male rats. Toxicology, 202: 145-158.
CrossRefDirect Link - Habig, W.H., M.J. Pabst and W.B. Jakoby, 1974. Glutathione S-transferases: The first enzymatic step in mercapturic acid formation. J. Biol. Chem., 249: 7130-7139.
CrossRefPubMedDirect Link - Heikal, T.M., M. El-Sherbiny, S.A. Hassan, A. Arafa and H.Z. Ghanem, 2012. Antioxidant effect of selenium on hepatotoxicity induced by chlorpyrifos in male rats. Int. J. Pharm. Pharmaceut. Sci., 4: 603-609.
Direct Link - Heikal, T.M., A.T.H. Mossa, M.A. Abdel Rasoul and G.I.K. Marei, 2013. The ameliorating effects of green tea extract against cyromazine and chlorpyrifos induced liver toxicity in male rats. Asian J. Pharm. Clin. Res., 6: 48-55.
Direct Link - Higdon, J.V. and B. Frei, 2003. Tea catechins and polyphenols: Health effects, metabolism and antioxidant functions. Crit. Rev. Food Sci. Nutr., 43: 89-143.
CrossRefPubMedDirect Link - Kalender, S., F.G. Uzun, D. Durak, F. Demir and Y. Kalender, 2010. Malathion-induced hepatotoxicity in rats: The effects of vitamins C and E. Food Chem. Toxicol., 48: 633-638.
CrossRefDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Mahgoub, A.A. and A.H. El-Medany, 2001. Evaluation of chronic exposure of the male rat reproductive system to the insecticide methomyl. Pharmacol. Res., 44: 73-80.
CrossRefPubMedDirect Link - Maity, S., J.R. Vedasiromoni and D.K. Ganguly, 1998. Role of glutathione in the antiulcer effect of hot water extract of black tea (Camellia sinensis). Jpn. J. Pharmacol., 78: 285-292.
CrossRefPubMedDirect Link - Mandel, S., O. Weinreb, L. Reznichenko, L. Kalfon, T. Amit, 2006. Green tea catechins as brain-permeable, non toxic iron chelators to iron out iron from the brain. J. Neural Transm. Suppl., 71: 249-257.
PubMed - Mansour, S.A. and A.T.H. Mossa, 2010. Oxidative damage, biochemical and histopathological alterations in rats exposed to chlorpyrifos and the antioxidant role of zinc. Pestic. Biochem. Physiol., 96: 14-23.
CrossRefDirect Link - McCollister, S.B., R.J. Kociba, C.G. Humiston, D.D. McCollister and P.J. Gehring, 1974. Studies of the acute and long-term oral toxicity of chlorpyrifos (O, O-diethyl-O-(3, 5, 6-trichloro-2-pyridyl) phosphorothioate). Food Cosmet. Toxicol., 12: 45-61.
CrossRef - Meeker, J.D., L. Ryan, D.B. Barr and R. Hauser, 2006. Exposure to nonpersistent insecticides and male reproductive hormones. Epidemiology, 17: 61-68.
CrossRefPubMedDirect Link - Misra, H.P. and I. Fridovich, 1972. The role of superoxide anion in the autoxidation of epinephrine and a simple assay for superoxide dismutase. J. Biol. Chem., 247: 3170-3175.
CrossRefPubMedDirect Link - Mossa, A.T.H., T.M. Heikal and E.A.A. Omara, 2012. Physiological and histopathological changes in the liver of male rats exposed to paracetamol and diazinon. Asian Pac. J. Trop. Biomed., 2: S1683-S1690.
CrossRefDirect Link - Nunez, S.B., R.P. Blye, P.M. Thomas, J.R. Reel, K.M. Barnes, J.D. Malley and G.B. Cutler Jr., 1996. Recovery of reproductive function in rats treated with the aromatase inhibitor fadrozole. Reprod. Toxicol., 10: 373-377.
CrossRefDirect Link - Ogutcu, A., Z. Suludere and Y. Kalender, 2008. Dichlorvos-induced hepatotoxicity in rats and the protective effects of vitamins C and E. Environ. Toxicol. Pharmacol., 26: 355-361.
CrossRefDirect Link - Pant, N., R. Shankar and S.P. Srivastava, 1996. Spermatotoxic effects of carbaryl in rats. Human Exp. Toxicol., 15: 736-738.
CrossRef - Rai, D.K. and B. Sharma, 2007. Carbofuran-induced oxidative stress in mammalian brain. Mol. Biotechnol., 37: 66-71.
CrossRefPubMedDirect Link - Saitoh, M., T. Yanase, H. Morinaga, M. Tanabe and Y.M. Mu et al., 2001. Tributyltin or triphenyltin inhibits aromatase activity in the human granulosa-like tumor cell line KGN. Biochem. Biophys. Res. Commun., 289: 198-204.
CrossRefDirect Link - Salvi, M., V. Battaglia, A.M. Brunati, N. La Rocca and E. Tibaldi et al., 2007. Catalase takes part in rat liver mitochondria oxidative stress defense. J. Biol. Chem., 282: 24407-24415.
CrossRef - Sarkar, R., K.P. Mohanakumar and M. Chowdhury, 2000. Effects of an organophosphate pesticide, quinalphos, on the hypothalamo-pituitary-gonadal axis in adult male rats. J. Reprod. Fertil., 118: 29-38.
PubMedDirect Link - Sarpa, M., R.R. De-Carvalho, I.F. Delgado and F.J. Paumgartten, 2007. Developmental toxicity of triphenyltin hydroxide in mice. Regul. Toxicol. Pharmacol., 49: 43-52.
CrossRef - Sharma, Y., S. Bashir, M. Irshad, S.D. Gupta and T.D. Dogra, 2005. Effects of acute dimethoate administration on antioxidant status of liver and brain of experimental rats. Toxicology, 206: 49-57.
CrossRefPubMedDirect Link - Singh, S.K. and R.S. Pandey, 1990. Effect of sub-chronic endosulfan exposures on plasma gonadotrophins, testosterone, testicular testosterone and enzymes of androgen biosynthesis in rat. Indian J. Exp. Biol., 28: 953-956.
PubMed - Venkatesh, S., M. Deecaraman, R. Kumar, M.B. Shamsi and R. Dada, 2009. Role of reactive oxygen species in the pathogenesis of mitochondrial DNA (mtDNA) mutations in male infertility. Indian J. Med. Res., 129: 127-137.
PubMedDirect Link - Wilke, T.J. and D.J. Utley, 1987. Total testosterone, free androgenic index and calculated free testosterone by analog RIA method. Clin. Chem., 33: 1372-1375.
PubMed - Yousef, M.I., K.I. Kamel, M.S. Hassan and A. El-Morsy, 2010. Protective role of propolis against reproductive toxicity of triphenyltin in male rabbits. Food Chem. Toxicol., 48: 1846-1852.
CrossRef