
Yogita Rajput
National Cave Research and Protection Organization, Central Zone, Raipur, 492001, C.G., India
Vibhuti Rai
School of Life Sciences, Pt. Ravishankar Shukla University, Raipur, 492010, C.G., India
Jayant Biswas
National Cave Research and Protection Organization, Central Zone, Raipur, 492001, C.G., India
Research Journal of Environmental Toxicology
Year: 2012 | Volume: 6 | Issue: 1 | Page No.: 13-24







ABSTRACT
Bacterial communities are often found to play a major role in building the foundation of food chain in the food starved cave ecosystem. Earlier it was assumed that due to lack of direct external environmental impact the caves were an almost sterile ecosystem. But today, the ever increasing human activities inside it, in the form of ecotourism exert a major impact on its native microbes, often stopping its growth and polluting the whole ecosystem. The situation is often found to be responsible for producing some human pathogenic bacteria inside it, which might pose a threat of infection to the other tourists. Kotumsar cave is a well known tourist pulling limestone cave from central part of India which is also a harbour of various native cavernicoles. In the present study, the bacterial communities existing in different sediments of this cave were screened and identified. The growth rates of each isolate were also verified against various temperature ranges and the maximum growth was found to coincide with the annual mean temperature of the cave. Further, the impact of the same has been correlated with the existing biodiversity, geophysical factors and the human activities inside the cave. Finally, the probabilities of pathogenic threats to human beings due to the respective bacterial communities have also been discussed.
PDF Abstract XML References Citation
Received: January 13, 2012;
Accepted: February 11, 2012;
Published: June 21, 2012
How to cite this article
Yogita Rajput, Vibhuti Rai and Jayant Biswas, 2012. Screening of Bacterial Isolates from Various Microhabitat Sediments of Kotumsar Cave: A Cogitation on Their Respective Benefits and Expected Threats for Complete Biosphere and Tourists. Research Journal of Environmental Toxicology, 6: 13-24.
DOI: 10.3923/rjet.2012.13.24
URL: https://scialert.net/abstract/?doi=rjet.2012.13.24
DOI: 10.3923/rjet.2012.13.24
URL: https://scialert.net/abstract/?doi=rjet.2012.13.24
INTRODUCTION
Subterranean caves are indeed a discontinuous continental subsurface biosphere; having perpetual darkness, high humidity (almost saturated), low energy sources with almost constant air and water temperatures which usually reflects the mean annual surface temperature of that particular zone. Due to complete lack of autotrophs, the inhabitants of caves usually depend on allochthonous organic inputs (Biswas, 2009) which generally enter via air currents, water streams (river springs or rain floods) and even via dripwaters or sheetflow through moist cave-wall surfaces (Paoletti et al., 2011). Nevertheless, some recent investigations have established the existence of a unique light independent ecosystem in some of the caves where the role of autotrophs is played by some chemosynthetic bacteria (Sarbu et al., 1994, 1996; Schabereiter-Gurtner et al., 2004; Engel et al., 2004; Por, 2007).
Micro-organisms definitely play a key role in the foodweb of subterranean cave ecosystem, the synthesizing activity of bacteria and other autotrophic organisms can be considered as the first stage of an endogenous flow. Further, the microbial biomass itself can also be considered as an important food source for cave-dwelling higher organisms. It has already been established that the bacteria which enter the caves by percolation water, acts as a major food source for metazoans (Gerie et al., 2004). Besides several other relevant examples (Vandel, 1965; Gunn, 2004; Culver and White, 2005; Culver and Pipan, 2009), the free-living subterranean copepods depend mainly on the bacteria, diatoms or other unicellular organisms for their energy sources and which could also be referred as the basic link in a food chain for the epikarst meiofauna (Walter, 2001). Conclusively, the micro-organisms act as a major source of energy in subterranean so called food-starved biosphere.
In spite of the above facts, the role of microorganisms in cave environments is still poorly explored. As per Gale (1992) the geophysical nature of most of the subterranean caves and their respective associated limitations for subterranean habitats remain relatively constant for thousands of years. Nevertheless, in the present day, due to the ever increasing anthropogenic pressures either in the form of eco-tourism; internal unrestrained (Gunn, 2004; Biswas et al., 2011) or by excavation/mining; external unrestrained (Biswas, 2009) the complete cave ecosystem is altering. Throughout the world, there is a long history of anthropogenic disturbance for subterranean ecosystems, with clear evidence of bacterial contamination at some sites (Gunn, 2004). Human approaches exert a major impact on microbes; consequently, polluting bacteria may pose a threat of infection to the tourists and guides. Caves, repeatedly visited by human beings are already reported as reservoirs of pathogenic microorganisms (Jurado et al., 2010). Recent reports from some tropical caves have warned about the pathogenic nature of a number of bacteria, fungi and viruses inhabiting caves. However, till date in India no serious attempt has been taken to understand this aspect.
Kotumsar cave is one of the most biologically explored limestone caves of India. Time to time several troglobiotic as well as troglophilic species have been reported from this cave (Biswas, 1992a; 2010). Unfortunately, till date except one (Rajput and Biswas, 2012) no serious attempt has been taken to learn the geomicrobial community of this cave, though day by day the tourist pressure is increasing in this cave. In the present study, the isolation and microscopic characterization of sediment-bacterial communities have been studied by collecting the sediment samples from different zones and microhabitat of Kotumsar cave. Further, their respective role with regard to the energy sources for other biotic components and/or pathogenic activity for the tourism have been tried to establish. Though the temperature is an essential parameter for any bacterial growth (Khan and Husaini, 2006; Mahin et al., 2011; Siddiqui et al., 2000; Khleifat et al., 2006; Williams et al., 2011), in the present study the optimum temperature required for maximum growth has also been established and its ecological importance has been discussed.
MATERIALS AND METHODS
Study site: Kotumsar cave (Fig. 1): It is situated in the Kanger Valley National Park (18°52’09” N; 81°56’05” E) at an altitude of 560 m and the stream of River Kanger is flowing at the foothill of this cave. The ambient external surface of this cave is covered by deciduous to mixed forest vegetation. The main entrance of this cave is formed by a vertical fissure in the wall of a hill which further leads inwards via a narrow, twisted tubular path, measuring about 15 m in length. The main tunnel of the cave is nearly 500 m long and has several lateral and downward passages leading to several irregular chambers. The roofs and walls of the different chambers are lined with colorful dripstone formations resulting from the precipitation of calcite-dissolved carbonate lime. In the deep zone, few epikarst water fed small pools are present (Pipan, 2005), which remain intact throughout the year. The cave is subjected to frequent flooding during the monsoon season which generally begins in the middle of June and continues till the mid of October. The chambers of the cave are always wet, floored with either rocks or pebbles of various dimensions or by surface-derived sediment/clay deposits which altogether makes the path slippery for the tourists. Presently this cave has became a major eco-tourist spot for state Chhattisgarh of India, since (2000 AD onwards) it was carved out from the state Madhya Pradesh as a new political entity.
Sample collection and preservation (Fig. 1): During the month of May (summer) sediment samples were collected from various different zones and microhabitats of the cave which are as follows: (1) Entrance zone (Red-laterite sediment), (2) Twilight zone (Red-laterite sediment), (3) Transient zone (Red-laterite sediment) (4) Transient zone (foval or brown sediment) (5) Deep zone (guano deposition) (6) Deep zone (guano mix sediment) (7) Deep zone (foval or brown sediment) and (8) Deep zone (Red-laterite sediment). Sediment samples were collected from 5-10 cm depth by gently scrapping of sediment, using sterile scrapples and collected in airtight sterile polythene bags after assigning them proper collection number with respective name of place from where the same was collected. Further, within the time duration of 10-12 h, the samples were transferred to the laboratory and preserved at 4°C.
Isolation of cave bacteria: Bacterial isolations were carried out by employing dilution plate (Waksman and Fred, 1922) and direct plate (Warcup, 1950) techniques, under which one gram of dried sediment sample was taken in 9 mL of sterile distilled water and mixed to nutrient agar medium (peptone, 5 g; beef extract, 3 g; NaCl, 5 g; agar 20 g; distilled water 1000 mL; pH 7.2). During the process, the colony-forming unit (CFU) was counted with the help of Colony counter, Digital, Lapiz, Medica Instrument, India.
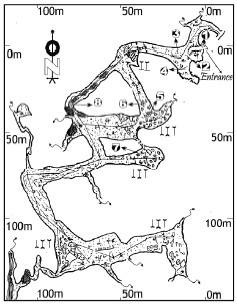 | |
| Fig. 1: | Layout of Kotumsar Cave (after Biswas, 1992b). One to eight numbers representing the site of soil collections: (1) Entrance zone (Red-laterite sediment), (2) Twilight zone (Red-laterite sediment), (3) Transient zone (Red-laterite sediment) (4) Transient zone (foval or brown soil sediment) (5) Deep zone (guano deposition) (6) Deep zone (guano mixed sediment) (7) Deep zone (foval or brown soil sediment) and (8) Deep zone (Red-laterite sediment) |
Selected colonies of bacteria were transferred from mother culture plates onto respective agar plates, incubated at 28°C for 24-48 h. Pure cultures were stored at 40°C until further examination.
Taxonomic grouping of bacterial isolates: Bacterial colonies were identified on the basis of morphological identity, biochemical test and their respective physiological activities, following the methods suggested by the Bergey’s Manual of Systematic Bacteriology (Schleifer, 1989; Aneja, 2005). Cultural characteristics of pure isolates in Nutrient agar media were recorded after incubation for 24-48 h at 28°C. Growth patterns of the cultivated bacterial colonies were evaluated on the basis of their respective sizes, pigmentations, configurations, margins and elevations. Morphological observations were made with a compound microscope (Leica, DSC).
Identification of bacteria: Bacterial identification at the preliminary stage was done by using Probabilistic Identification of Bacteria (PIB) Win software, which is one of the data base developed for the identification of bacteria based on numerical taxonomy (Langham et al., 1989).
Temperature dependent bacterial growth: The bacterial isolates were grown in different temperature ranges starting from 10 till 75°C and the temperature at which maximum growth were seen was tabulated.
RESULTS
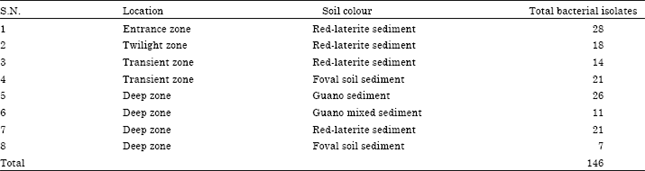
During isolation, total 146 bacterial isolates were recorded from sediment samples collected from eight different microhabitats of Kotumsar cave (Table 1). Among these about 19% (28) of bacterial population was isolated from the sediment collected from the entrance zone (nearer to the cave entrance), 12% (18) from the twilight zone, 10% (14) from the transient zone (Red-laterite sediment), 14% (21) from transient zone (foval soil sediment), 18% (26) from deep zone (guano sediment), 8% (11) from deep zone (guano mixed sediment), 14% (21) from deep zone (foval soil sediment) and 5% (7) from deep zone (ordinary red-laterite sediment). By employing PIB-win (Probabilistic identification of bacteria) identification system we identified and selected twenty-one bacterial isolates (Table 2) for further tests.
The growth patterns of bacterial isolates were strictly found to be temperature dependent. Minimum growth rate were recorded at initial temperature of 10°C, whereas the maximum growth in most of the bacterial isolates were seen at 28°C. An exceptional result was withstood with Deinococcus radiodurans, isolated from guano sediment of deep zone, which recorded maximum growth at 45 to 55°C (Table 3).
| Table 1: | Number of bacterial strains isolated from various microhabitat sediments of Kotumsar cave |
 | |
| Table 2: | Cultural and biochemical characteristics of bacteria isolated from various sediments of Kotumsar cave |
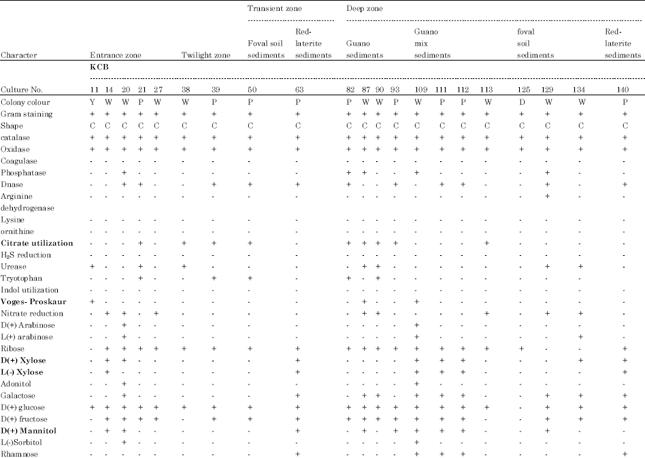 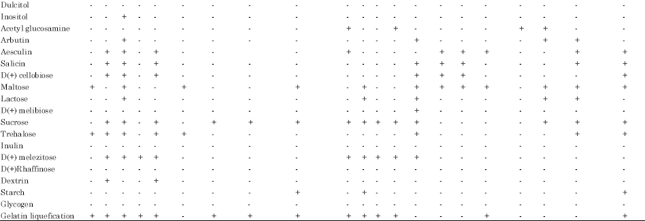 | |
| Identified name and its identification score: KCB11: Micrococcus luteus 4, KCB14: Stomatococcus mucilaginosus, KCB20: Staphylococcus sciuri, KCB21: Deinococcus radiodurans, KCB26: Stomatococcus mucilaginosis from outside of the entrance gate; KCB38: Micrococcus luteus 3, KCB39: Deinococcus radiodurans from entrance zone; KCB50: Deinococcus radiodurans, KCB63: Micrococcus agilis from middle zone; KCB87: Staphylococcus epidermis, KCB90: Micrococcus luteus 1, KCB93: Deinococcus radiodurans, KCB82: Deinococcus radiodurans Inner zone (guano sediments); KCB109: Streptococcus grpQ1, KCB111: Micrococcus agilis, KCB112: Micrococcus agilis, KCB113: Micrococcus luteus 2 from inner zone (black sediments), KCB125: Micrococcus agilis, KCB134: Staphylococcus varians, KCB129: Staphylococcus simulans from inner zone (brown sediments); KCB140: Micrococcus agilis from innermost zone, Whereas; KCB: Kotumsar cave bacteria, C: Cocci, O: Oxidative, F: Fermentative, +: Positive test, -: Negative test, Y: Yellow orange, W: White cream, P: Pink red, D: Dark yellow | |
| Table 3: | Effect of temperature on the growth of bacterial isolates from Kotumsar cave (values Mean ± SE) |
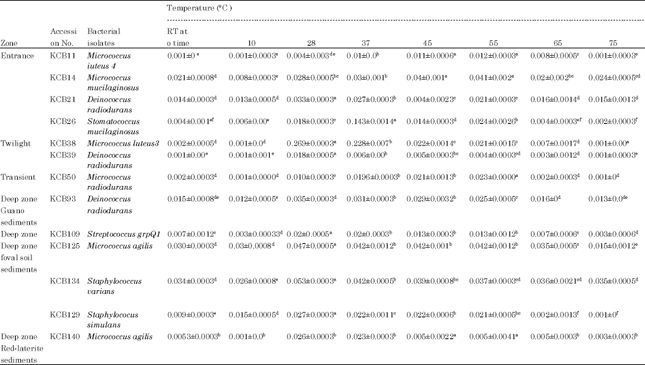 | |
| Whereas, means having similar alphabates within the individual bacterial isolates as superscript, were not statistically significant from each other at p<0.05 (based on Duncan’s multiple-range test) | |
DISCUSSION
In caves, the microbial activity is known to degrade the allochthonous organic inputs into the simpler detrital form (Humphreys, 1991; Bottrell, 1996), though the complete processes are similar to surface sediments (Bottrell, 1996). The microbes which are presumably introduced in the caves with the surface-derived soil are however capable enough to break down the complex surface-derived organic inputs in the sediment and thus form the basis of subterranean food chains. Microbes may also produce some growth factors, such as vitamins, needed by higher organisms (Northup and Lavoie, 2004).
Due to the absence of several pollutant factors, the cave ecosystem could be referred as atmospherically sterile biosphere. Microorganisms in caves grow in some specific and distinct habitats where they develop various interactions (Mulec, 2008). Thus, it could be stated that the microbes occurring in the cave are usually microhabitat dependent, though the various geophysical factors are found to be almost constant in it. Kotumsar cave ordeals bi-annual flooding (Biswas, 1992a) during which high amount of allochtonous organic matter alongwith extraneous organisms enter and consequently heterotrophic decomposition processes start as an immediate effect. Thus, after flooding a long period is obviously required to restore the normal situation in this cave. In the present study, the sample for bacterial cultivations were collected during summer (an almost constant period) and as per expectation, the maximum bacterial isolates were recovered from the soil sediment samples collected from outside the cave i.e., from the sediment exposed to the open environmental conditions (19%). Interestingly, the isolates in almost same proportion were found from the guano sediment (18%) of Deep zone, followed by foval soil sediment (14%) collected from both transient and deep zones. In Kotumsar cave, the guano depositions have already been reported to support a vast community of cavernicoles (Biswas, 2010), following which the occurrence of healthy community of bacterial isolates is highly justified. Simultaneously, the healthy communities of bacteria from the clayey slimy soil sediment “foval” collected either from transient or deeper zone also somehow support the earlier findings (Camassa, 1997). Further, except the Deinococcus radiodurans; isolated from guano sediment of deep zone, rest all the other isolated bacterial communities showed their maximum growth at 28°C and which is already reported as the mean temperature of the cave (Biswas, 1992a, 2010). However, earlier reports already suggests the Deinococcus radiodurans, a extremophilic radioresistant bacterium. Therefore, it clearly indicates that the environmental conditions of the Kotumsar cave offer the best suited growth conditions to the isolated microbes for this particular zone.
Today the frequency of human visits to many caves; either in the form of eco-tourism, research or other religious aspects (especially in South Asian countries) has increased many folds. It is therefore the need of the hour to know about the possible potentially pathogenic microorganisms present in caves and to make the tourists alert regarding the degree of vulnerability to which currently they are unknown. Groth et al. (1999) have isolated several species of the genera Amycolatopsis, Aureobacterium, Brevibacterium, Nocardia, Nocardioides, Rhodococcus, Streptomyces and the family Micrococcaceae from the caves of Spain and most of which have been catalogued as pathogenic to human beings. Although, it is difficult to associate any specific infection to a single visit to a subterranean environment, but the repetitive visits may prove a fatal factor to the visitors. In our study we isolated few such bacterial communities, the occurrences of which could only be possible by human intervention. Stomatococcus mucilaginosus; isolated from the sediment collected from the entrance zone is a grampositive, catalase variable bacterium which is usually present in the saliva and upper respiratory tract of human beings. Recent reports suggest that it could be a cause for certain pathogenic diseases in human beings (Guermazi et al., 1995; Van Tiel et al., 1995; Nielsen, 1994). Further, Micrococcus luteus, which was also recovered from the same site is a common pathogen (Oladipo and Omo-Adua, 2011; Tambekar et al., 2008; Adesina et al., 2011) and is usually related with skin disease and affects mainly immune compromised patients (Salar et al., 1997). Threats of the same isolates have also been found to exist in the sediment contents of transient as well as deeper zones. The bacterial species; Staphylococcus simulans isolated from the foval soil sediment of the deep zone has also been reported to prove osteomyelitis related fatal diseases in some recent cases (Al-Kline, 2010; Razonable et al., 2001). Staphylococcus sciuri which was isolated from the collected sediment sample of the entrance zone was also recognized as pathogen for some human diseases (Wang et al., 2007; Chen et al., 2007). Further, the existence of human pathogenic fungi and yeasts are also reported to be common in caves (Sugita et al., 2005), the existence of which in the present cave could not be ruled out.
Conclusively, the bacterial species isolated from the different habitats of Kotumsar cave sediment could be easily assumed to support directly or indirectly the biodiversity of the cave in providing energy stuffs. Possibly, few isolates recovered from the sediment of this cave (especially Micrococcus luteus) may also participate in speleothem processes as it was speculated about the same species in the caves of Meghalaya (Baskar et al., 2009) India. Similarly, Staphylococcus sciuri; reported from the cave of China (Wang et al., 2007) and Micrococcus varians from Poland cave pleaded to be a part of speleothem. However, various human beings oriented bacteria (Staphylococcus epidermis, Stomatococcus mucilaginosus, Staphylococcus salivarius, Staphylococcus sciuri, Micrococcus varians) were also isolated from this cave which directly suggests that the cave is under high anthropogenic pressure, though in normal case those microbes are not too fatal for human beings. The tourist visiting the cave; frequently comes in contact with the floor sediment, especially due to its slippery path they have to take support of the ground and walls. Thus, it is advisable, to clean their hand after leaving the spot and not use their unwashed hands to eat anything during caving.
This piece of work is just preliminary information regarding the sediment bacterial activities present in this cave. However, the work on molecular level is absolutely needed to suggest perfectly regarding the cave management practices, which will certainly help the tourist to take precautions before and after caving to get rid by pathogenic microbes and simultaneously to save the cave environment.
ACKNOWLEDGMENTS
We are highly grateful to the director of Kanger Valley National Park of Chhattisgarh, India for allowing us to conduct this study. Thanks are due to Dr. Anil Kumar Ramteke and Siddharth Biswas, National Cave Research and Protection Organization, India for collecting the sediment samples for this study.
REFERENCES
- Mahin, A.A., M.G. Sorrowar, M.Z. Hasan, M.M. Haque and H. Rashid, 2011. Effect of low temperature, radiation and their combination treatments on microorganisms associated with fresh water mola fish, Amblypharyngodon mola (Hamilton- Buchanan, 1822). Asian J. Biotechnol., 3: 507-518.
Direct Link - Adesina, K., A.A. Oshodi, T.A.M. Awoniyi and O.O. Ajayi, 2011. Microbiological assessment of cow milk under traditional management practices in Ado-Ekiti, Nigeria. Pak. J. Nutr., 10: 690-693.
CrossRefDirect Link - Rajput, Y. and J. Biswas, 2012. Subterranean depth dependent protein constitutions of the Micrococcus sp., isolated from the Kotumsar Cave, India. Asian J. Biochem., 7: 90-97.
CrossRefDirect Link - Baskar, S., R. Baskar, N. Lee and P.K. Theophilus, 2009. Speleothems from Mawsmai and Krem Phyllut caves, Meghalaya, India: Some evidences on biogenic activities. Environ. Geol., 57: 1169-1186.
CrossRef - Biswas, J., 2009. The biodiversity of Krem Mawkhyrdop of Meghalaya, India, on the verge of extinction. Curr. Sci., 96: 904-910.
Direct Link - Biswas, J., 2010. Kotumsar Cave biodiversity: A review of cavernicoles and their troglobiotic traits. Biodivers. Conser., 19: 275-289.
CrossRefDirect Link - Biswas, J., S. Shrotriya, Y. Rajput and S. Sasmal, 2011. Impacts of ecotourism on bat habitats in caves of Kanger Valley National Park, India. Res. J. Environ. Sci., 5: 752-762.
CrossRefDirect Link - Bottrell, S.H., 1996. Organic carbon concentration profiles in recent cave sediments: Records of agricultural pollution or diagenesis? Environ. Pollut., 91: 325-332.
CrossRefPubMedDirect Link - Chen, S., Y. Wang, F. Chen, H. Yang, M. Gan and S.J. Zheng, 2007. A highly pathogenic strain of Staphylococcus sciuri caused fatal exudative epidermitis in piglets. PLoS ONE, Vol. 2, No. 1.
CrossRefDirect Link - Engel, A.S., M.L. Porter, L.A. Stern, S. Quinlan and P.C. Bennett, 2004. Bacterial diversity and ecosystem function of filamentous microbial mats from aphotic (cave) sulfidic springs dominated by chemolithoautotrophic Epsilonproteobacteria. FEMS Microbiol. Ecol., 51: 31-53.
CrossRefPubMedDirect Link - Gale, S.J., 1992. Long-term landscape evolution in Australia. Earth Surf. Processes Landforms, 17: 323-343.
CrossRefDirect Link - Gerie, B., T. Pipan and J. Mulec, 2004. Diversity of culturable bacteria and meiofauna in the epikarst of Skocjanske jame caves (Slovenia). Acta Carsol., 33: 301-309.
Direct Link - Groth, I., R. Vetterman, B. Schuetze, P. Schumann and C. Saiz-Himenez, 1999. Actinomycetes in karstic caves of Northern Spain (Altamira and Tito Bustillo). J. Microbiol. Methods, 36: 115-122.
CrossRef - Guermazi, A., Y. Miaux and M. Laval-Jeantet, 1995. Imaging of choroid plexus infection by Stomatococcus mucilaginosus in neutropenic patients. AJNR Am. J. Neuroradiol., 16: 1331-1334.
PubMedDirect Link - Humphreys, W.F., 1991. Experimental re-establishment of pulse-driven populations in a terrestrial troglobite community. J. Anim. Ecol., 60: 609-623.
Direct Link - Jurado V., L. Laiz, V. Rodriguez-Nava, P. Boiron, B. Hermosin, S. Sanchez-Moral and C. Saiz-Jimenez, 2010. Pathogenic and opportunistic microorganisms in caves. Int. J. Speleol., 39: 15-24.
Direct Link - Khan, F.A.B.A. and A.A.S.A. Husaini, 2006. Enhancing α-amylase and cellulase in vivo enzyme expressions on sago pith residue using Bacilllus amyloliquefaciens UMAS 1002. Biotechnology, 5: 391-403.
CrossRefDirect Link - Khleifat, K., M. Abboud, W. Al-Shamayleh, A. Jiries and K.A. Tarawneh, 2006. Effect of chlorination treatment on gram negative bacterial composition of recycled wastewater. Pak. J. Biol. Sci., 9: 1660-1668.
CrossRefDirect Link - Langham, C.D., S.T. Williams, P.H. Sneath and A.M. Mortimer, 1989. New probability matrices for identification of Streptomyces. J. Gen. Microbiol., 35: 121-133.
PubMed - Mulec, J., 2008. Microorganisms in hypogean: Examples from Slovenian Karst caves. Acta Carsol., 37: 153-160.
Direct Link - Nielsen, H., 1994. Vertebral osteomyelitis with Stomatococcus mucilaginosus. Eur. J. Clin. Microbiol. Infect. Dis., 13: 775-776.
CrossRefPubMedDirect Link - Oladipo, I.C. and R.O. Omo-Adua, 2011. Antibiotics resistance among bacteria isolated from evaporated milk. Asian J. Biol. Sci., 4: 77-83.
CrossRefDirect Link - Paoletti, M.G., M. Beggio, A.L. Dreon, A. Pamio and T. Gomiero et al., 2011. A new foodweb based on microbes in calcitic caves: The Cansiliella (Beetles) case in Northern Italy. Int. J. Speleol., 40: 45-52.
CrossRefDirect Link - Por, F.D., 2007. Ophel: A groundwater biome based on chemoautotrophic resources. The global significance of the Ayyalon cave finds, Israel. Hydrobiologia, 592: 1-10.
CrossRefDirect Link - Razonable, R.R., D.G. Lewallen, R. Patel and D.R. Osmon, 2001. Vertebral osteomyelitis and prosthetic joint infection due to Staphylococcus simulans. Mayo Clin. Proc., 6: 1067-1070.
CrossRefPubMedDirect Link - Salar, A., J. Carratala, A. Fernandez-Sevilla, D. Marin and A. Granena, 1997. Pneumonia caused by Micrococcus species in a neutropenic patient with acute leukemia. Eur. J. Clin. Microbiol. Infect. Dis., 16: 546-548.
PubMedDirect Link - Sarbu, S.M., B.K. Kinkle, L. Vlasceanu, T.C. Kane and R. Popa, 1994. Microbiological characterization of a sulfide-rich groundwater ecosystem. Geomicrobiol. J., 12: 175-182.
CrossRefDirect Link - Sarbu, S.M., T.C. Kane and B.K. Kinkle, 1996. A chemoautotrophically based cave ecosystem. Science, 272: 1953-1955.
CrossRefDirect Link - Schabereiter-Gurtner, C., C. Saiz-Jimenez, G. Pinar, W. Lubitz and S. Rolleke, 2004. Phylogenetic diversity of bacteria associated with Paleolithic paintings and surrounding rock walls in two Spanish caves (Llonyin and La Garma). FEMS Microbiol. Ecol., 1606: 235-247.
CrossRefDirect Link - Siddiqui, H., T. Masud and M. Arif, 2000. Effect of incubation temperature and pH on Lactococcus lactis ssp. Diacetylactis and their virulent phages. Pak. J. Biol. Sci., 3: 1588-1590.
CrossRefDirect Link - Sugita, T., K. Kikuchi, K. Makimura, K. Urata and T. Someya et al., 2005. Trichosporon species isolated from guano samples obtained from bat-inhabited caves in Japan. Applied Environ. Microbiol., 71: 7626-7629.
CrossRefDirect Link - Tambekar, D.H., P.B. Gulhane, S.G. Dahikar and M.N. Dudhane, 2008. Nosocomial hazards of doctor's mobile phones in hospitals. J. Medical Sci., 8: 73-76.
CrossRefDirect Link - Van Tiel, F.H., B.F. Slangen, H.C. Schouten and J.A. Jacobs, 1995. Study of Stomatococcus mucilaginosus isolated in a hospital ward using phenotypic characterization. Eur. J. Clin. Microbiol. Infect. Dis., 14: 193-198.
PubMedDirect Link - Waksman, S.A. and E.B. Fred, 1922. A tentative outline of the plate method for determining the number of micro-organisms in the soil. Soil Sci., 14: 27-28.
Direct Link - Wang, H., X. Yang, C. Hu, S. Mao, X. Zhao, Q. Liu and C. Zeng, 2007. Preliminary Study on Microbial Communities in Dripping Waters of Heshang Cave, Hubei Province, P.R. China. In: Water-Rock Interaction, Wang, Y. and T.D. Bullen (Eds.). Taylor and Francis Group, London, UK., ISBN-13: 9780415451369, pp: 925-928.
- Warcup, J.H., 1950. The soil-plate method for isolation of fungi from soil. Nature, 166: 117-118.
PubMedDirect Link - Biswas, J., 1992. Kotumsar cave ecosystem: An interaction between geophysical, chemical and biological characteristics. NSS Bull., 139: 7-10.
Direct Link - Al-Kline, D.P.M., 2010. Staphylococcus simulans osteomyelitis of the foot: A case report. Foot Ankle Online J., Vol. 3.
CrossRefDirect Link