
Amal W. Amin
Department of Botany and Microbiology, Faculty of Science, Alexandria University, Post Code 21511, El-Shatby, Alexandria, Egypt
Research Journal of Environmental Toxicology
Year: 2011 | Volume: 5 | Issue: 4 | Page No.: 235-250







ABSTRACT
The importance of the reuse of sewage sludge is derived from its high nutrient content that can improve the characteristics of the soil and kind of crop. Accordingly, it is important to characterize the genotoxicological potential of sewage sludge amended soil with such valuable properties "benefits". Field experiment was conducted to evaluate the residual effect of previous applications of sewage sludge used as biofertilizer on M2 meiocytes of Zea mays with different sludge rates (10, 20, 30 and 40 T/F for one and two additions). The resulting data demonstrated that M2 plants still have mutations in pollen mother cells. The total percentage of aberrant mother cells increased significantly with increasing sewage sludge concentration either by residual one or two additions. Aberrations was restricted to Metaphase one of Division one (DI) in the two groups of treatments, while, aberrant end product was the most frequent stage in Division two (DII). The most frequent kinds of aberrations were abnormal chromosome distribution and orientation, sticky figures, abnormal spindle behavior, overlapping cells, asynchronized cells and abnormal cell contour.
PDF Abstract XML References Citation
Received: January 22, 2011;
Accepted: July 22, 2011;
Published: October 31, 2011
How to cite this article
Amal W. Amin, 2011. Evaluation of the Genotoxicity of Residual Repeated Applications of Sewage Sludge on M2 Meiocytes of Zea Plants. Research Journal of Environmental Toxicology, 5: 235-250.
DOI: 10.3923/rjet.2011.235.250
URL: https://scialert.net/abstract/?doi=rjet.2011.235.250
DOI: 10.3923/rjet.2011.235.250
URL: https://scialert.net/abstract/?doi=rjet.2011.235.250
INTRODUCTION
Organic wastes are added to agricultural soils for increasing soil fertility and plant nutrition. Plant and animal wastes as well as sewage sludge provided by municipal sewage sludge refineries of urban wastes, has been added to agricultural soils to increase its organic matter. Previous studies have demonstrated favorable plant yield responses to the application of sewage sludge (Amin et al., 2009a, b; Ashour et al., 2007). In contrast, the effects of sewage sludge on soil biological processes have been questioned by some authors (Solanoa et al., 2009; Kizilkaya and Bayrakli, 2005; Knight et al., 1997; Banerjee et al., 1997). Knight et al. (1997) observed a decrease of soil biological activity, such as microbial biomass and enzyme activities, due to sewage sludge application. Conversely, Sastre et al. (1996) and Banerjee et al. (1997) found that sewage sludge amendment increased soil microbial activity and CO2 production. (Kizilkaya and Bayrakli, 2005) stated that enzymatic activities in sludge-amended soils tended to decrease by increasing incubation time. These differences might be a result of the difference in toxic metal content of different sources of sewage sludge and/or its stability (Tam and Wong, 1990). Heavy metals are significantly toxic, due to their cumulative nature in different body organs leading to unwanted effects (Jarup, 2003; Sathawara et al., 2004). Metals tend to reach toxic limits even when found in low concentrations. Monitoring programs are being carried out in many countries to avoid the distribution of foodstuffs that could pose a risk to human health if consumed. The chemical interactions of toxic elements can interfere with proteins in living systems render essential reactions important (Diamond and Zalups, 1998).
Applying treated municipal sewage sludge to agricultural soils to improve its organic content has been used in Egypt for several decades. While plant nutrients and organic matter are essential to plants, excessive application of nitrogen and phosphorus was found to contaminate surface and groundwater or interfere with good crop growth (Pivetz, 2001). Sludge also may contain metals and some other constituents that can be harmful to humans if accumulated applications are not managed properly. At the least, sludge application can create a nuisance. Several communities regularly use sludge as biofertilizer to benefit from municipal sewage. However, because of new regulations governing the application of sludge to land, many questions are being asked by farmers, utility managers, government officials and consultants about this alternative (Singh and Agrawal, 2010). The Government’s Regulation for the application of sewage sludge should be applied in order to achieve the following: pathogens disinfection, organic pollutants degradation, less available heavy metals (as Cd, Pb, Ni, Co, Cr, Zn, Cu, Fe, Mn) and seed weeds destructions. In addition, sewage sludge loading to agriculture land should be monitored as excess application (El-Motaium and Abdel Monem, 2000; Ghazy et al., 2009). Fuerhacker and Haile (2011) pointed out the outcomes of different risk assessments.
Several studies using bean, maize and barley were stated dangerous levels of some heavy metal(s) in the composted sewage sludge obtained from Alexandria General Organization of Sanitary Drainage (AGOSD) of Alexandria (Amin, 2001; Amin and Sherif, 2001; Ashour et al., 2007; Amin et al., 2009a, b).
The sewage sludge production is continuously increasing in Egypt. Therefore, the main current pressing needs are to find and develop more efficient sustainable technologies to allow a safe and suitable reuse of sewage sludge in agriculture. Furthermore, the legislations should be more adapted to Egyptian conditions as well as improvement of the institutional capacity to guarantee the enforcement of their application.
In vivo and in vitro, higher plants function as biosensors of pollutants. Plants are able to give repeatable and quantifiable responses. The biological assay-guided chemical analysis (the association between biological and chemical approach) is the best way to obtain information about the kind of polluting agents, their origin and fate in environment and living organisms. The choice of pollen mother cells of corn as biosensors of pollutants causing mutations in crop plants has advantages in that very large number of pollen grains can be obtained from one tassel. Each kernel arising from treated pollen is potentially heterozygous for a unique induced mutation that can be expressed immediately (if dominant) or in the progeny of the M1 (if recessive). Plewa (1982) recommended that Zea mays bioassays can be used to assess genotoxicity from potential chemical and environmental pollutants. From the many meiotic mutants described in plants and other organisms, it is obvious that meiosis is controlled by a large number of genes (Chaudhury et al., 1992; Bhatt et al., 2001), the knowledge of which is mainly restricted to their cytological and genetic characteristics.
This study provides information to help answer questions and evaluate the option of using municipal sewage water and sludge instead of chemical fertilizers. Therefore, the aim of this work was to monitor the in situ genotoxic and cytotoxic potentialities of maize grown on agricultural soil having residual sewage sludge. The chemical composition of the used sewage sludge was carried out by Amin and Sheriff (2001). Using the second generation of treated Zea plants and comparing its net results with (Amin et al., 2009a) who stated that some heavy metals accumulation in leaves and grains (Cu, Zn, Cd, Pb) as a result of residual sewage sludge treatments may lead to genotoxicity through affecting the genes responsible for some Zea yield parameters and kernel characters.
Analysis of the clastogenic effects of metals and their compounds on higher organisms indicates some general trends by Sharma and Talukder (1987). Following chronic exposure to subtoxic doses, a decrease in mitotic frequency and increase in chromosomal abnormalities were observed. These effects are usually directly proportional to the dose applied and the duration of treatment within the threshold limits. Recovery after acute treatment is inversely related to the dosage. The ultimate expression of the effects depends on the test system used. In plants, the solubility of the metals in water is of much greater importance. The degree of dissociation of metallic salts and the rate of absorption affect significantly the frequency of chromosomal aberrations. Therefore, evaluation toxic metal levels can be assessed and compared by Joint Expert Committee on Food Additives (JECFA) of the Food and Agriculture Organization of the Joint Nations and the World Health Organization United Nations and the World Health Organization International Programme on Chemical Safety FAO and WHO (2000, 2001).
MATERIALS AND METHODS
The present study was carried out at the field station of faculty of agriculture, Alexandria University. Composted sewage sludge was obtained from Alexandria General Organization of Sanitary Drainage (AGOSD) of Alexandria city in February, 1999. The chemical composition of sludge and physical and chemical characteristics of the soil were determined according to (Page et al., 1982) and are presented by Amin (2001). A field experiment was carried out in 1999 to evaluate the effect of residual sewage sludge, one and two additions on M1 yield parameters of Zea mays L. variety H320 (Amin et al., 2009a). Several rates of sewage sludge were used (0, 10, 20, 30 and 40 T/F). During this experiment, immature male tassels at the boot stage (~about eleven weeks old Zea) were collected, from six different plants from each treatment and fixed immediately in fresh solutions of ethanol: glacial acetic acid (3:1, v/v). Semipermanent acetocarmine and acetooreicin meiocytes preparations were examined. Pollen Mother Cells (PMC’s) at different meiotic stages were screened and various chromosomal and cellular irregularities were recorded.
The percentage of aberrant PMCs for each treatment was calculated as the number of aberrant PMCs/total number of cells, while the percent of each aberrant type was calculated as the number of one type of aberrant PMCs/total number of examined PMC's in each treatment. The number of aberrations per PMCs was calculated as total kinds of aberrations/total number of aberrant PMCs. The Percentage of sterile pollen grains was also estimated using acetocarmine stain and calculated from total examined pollen grains.
Statistical analysis: Statistical analysis of data was carried out using the one-way Analysis of Variance (ANOVA) and significant differences between the means were assessed by Duncan’s multiple comparison tests at p = 0.05.
where, n is the sample size (total number of pollen mother cells) and X the percent of aberrant pollen mother cells.
RESULTS AND DISCUSSION
In the present study an appreciable number of meiotic aberrations were recorded in pollen mother cells of M2 plants grown on soil having residual one and two additions with different rates of sewage sludge- 0, 10, 20, 30 and 40 T/F (Table 1-3, Fig. 1, 2).
| Table 1: | Percentage of aberrant stages of M2 meiocytes of plants grown on soil with one and two residual sewage sludge addition (calculated from Total meiocytes) |
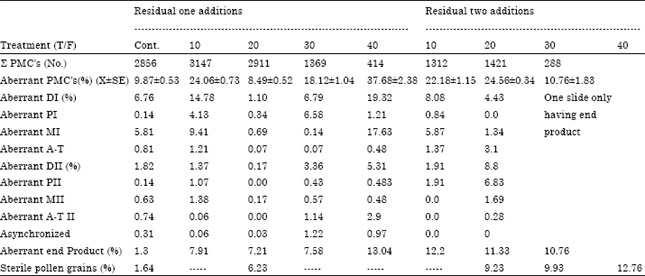 | |
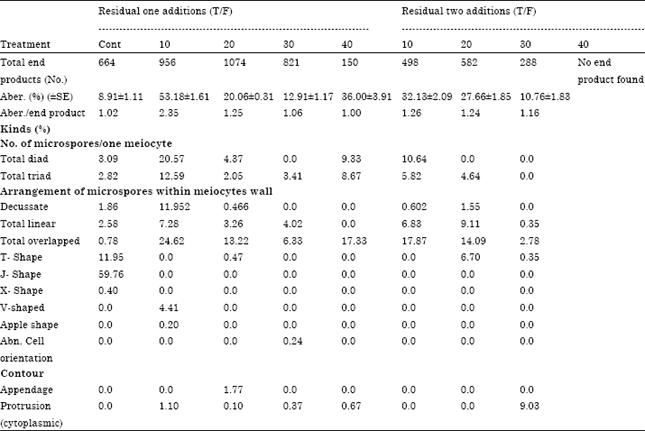
Frequency of aberrant meiocytes and meiotic phases: Generally, the percentage of aberrant mother cells increased significantly by either residual one or two residual sludge addition. The mean percentage of aberrant mother cells increased about three folds than the control with increasing sewage sludge concentration by residual one and/or two additions, except 20 T/F residual one addition (Table 1). This lowest percentage of aberrations of 20 T/F could be attributed to the high content of organic matter in respect to heavy metals present in this low rate of sludge (Sang and Li, 2002). Organic matter ability to sequester most heavy metals found in soil and sludge lowered their cytotoxic effect. This agreed with Ashour et al. (2007) using wheat as a plant bioassay. However, the mean aberration percentage in both treatments indicated that the residual two additions treatments were more cytotoxic than residual one addition (20.06 and 15.96, respectively). The percentage of aberrant PMC’s at meiotic Division two (DII) caused by residual two additions were three times higher than those at meiotic Division one (DI). The most stage of aberrations of DI was restricted to metaphase in all treatments while, aberrant end product formed the most frequent stage in DII. These are all different products than the normal tetrad stage. It amounted to six to ten times higher than those of the control.
Aberrant meiocytes contained about 1.1 kinds of aberrations per PMC for all treatments (Table 2). The highest kind of aberrations in division I was abnormal chromosome distribution which appeared in metaphase I as absence of spindle microtubules or abnormal chromosome movement (precocious, laggards) and unequal polar groups (Fig. 1). These kinds were more frequent in the 10T/F than the other treatments. Sticky figures were induced by all residual one addition and 10T/F residual two additions treatments. In the same time, the percentage of sticky chromosomes increased by increasing sludge concentrations except 20 T/F induced the lowest value (1.6%). Generally, abnormal spindle behavior either inhibition or microtubule orientation and irregular chromosome movement decreased by all sludge treatments except 10 T/F two additions. Prophase one with two nuclei, less or more bivalent number and fragmented bivalents at diakinesis appeared in control and all residual one addition and 10 T/F residual two additions treatments. Telophase cells with protrusion at the position of cell plate were induced only by 10 and 30 T/F in division I and 30 and 40 T/F in division II residual one addition.
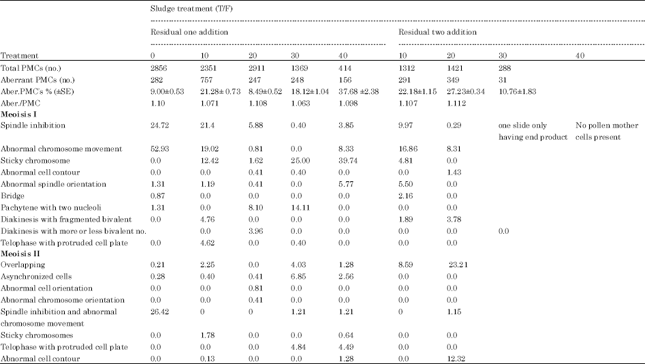
| Table 2: | The percentage of aberrant end products of aberrant M2 Meiocytes of plants grown on soil with one and two residual sewage sludge addition (calculated from aberrant meiocytes) |
 | |
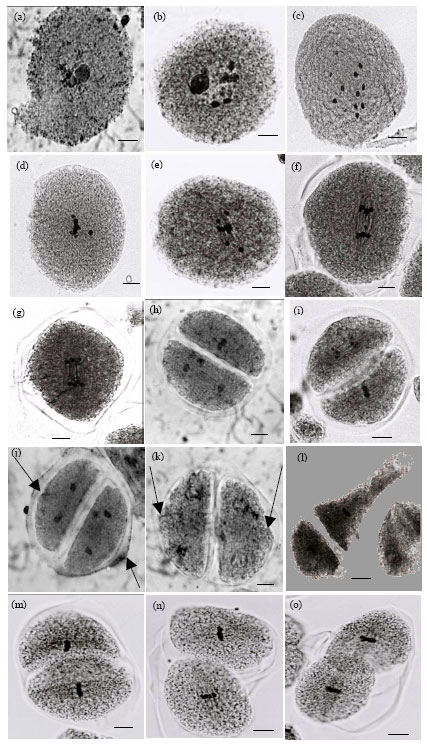 | |
| Fig. 1(a-o): | M2 Meiocytes of Zea mays at DI and II with different kinds of aberrations: (a): Pachytene with two nuclei, (b): Diakinesis with fragmented and less bivalent number, (c): C-Metaphase I, (d): Metaphase I with precocious chromosomes, (e): Metaphase I with laggard, (f): Anaphase I with laggard chromosome, (g): Anaphase I with bridges, (h): Asynchronized cell, (i): Overlapped and asynchronized cell, (j and k): Telophase II with Protrusions at the position of cell plate, (l): Metaphase II with abnormal chromosome orientation and cell contour, (m-o): different degrees of overlapping and abnormal cell orientation |
The highest kind of aberrations in division II was induced by one-addition treatments. Division II meiocytes contained high percent of overlapping cells and abnormal cell contour with highest value induced by 20T/F two additions. The highest percentage of asynchronized cells and telophase cells with protrusion at the position of cell plate occurred after 30T/F one addition.
However, while abnormal cell contour, abnormal chromosome orientation and sticky figures were found in sludge treatments only, high percent of abnormal chromosome movement was found in control and decreased to very low values by some sludge treatments.
In the present study, although all flower buds were collected from all treatments at the same time, the highest sludge rates (30 and 40 T/F) of the residual two additions were found to be pollen grains only in addition to the absence of meiotic end product in 40 T/F treatment. This may be suggested that residual sludge enhanced the rate of meiotic division leading to the enhancement of maturity of zea plant (earliness). Similar result was found by Ashour et al. (2007) using wheat plant.
Cytological events of meiosis are controlled by a large number of genes acting from pre-meiosis to the post-meiotic mitosis. Natural and induced mutations that affect many steps of the process have been widely described in maize and other species (Golubovskaya, 1989). Therefore, mutations of these genes may cause anomalies that impair plant fertility (Ricci et al., 2007; Kaul and Murthy, 2004).
In the present study, chromosome stickiness might be due to a recessive gene mutation resembling that discovered in maize by Beedle (McGill et al., 1974), ending with immediate reactions with DNA causing DNA-DNA or DNA-protein cross linking (El-Kodary et al., 1990; El-Ghamery et al., 2000; Kovaleva, 2008). Gaulden (1987) postulated that sticky chromosomes may result from the defective functioning of one or two types of specific non-histone proteins involved in chromosome organization which are needed for chromatid separation and segregation. The altered functions of these proteins leading to stickiness may be caused by mutations in the structural genes coding for them (hereditary stickiness) or by the action of mutagens on the proteins (induced stickiness). According to Kumar and Rai (2007), it seems most probable that the heavy metals may have caused some kinds of gene mutations which led to incorrect coding of some non-histone proteins involved in chromosome organization.
In the present study, residual sewage sludge reduced the percent of chromosomes not able to congregate in the metaphase plate, remaining scattered in the cytoplasm. Absence or defective spindle fibers or defects in kinetochore that impair spindle fiber attachment could be responsible for such abnormality. A mutation involving absence of spindle fibers and causing inability of bivalent congression at the metaphase plate was reported in maize by Daniela et al. (2005) and might be inhibited by residual sewage sludge.
Many factors of both cytoplasmic and nuclear origin could influence spindle formation and chromosome alignment. Dysfunctional cytoplasm could lead to formation of abnormal meiotic spindles and induce chromosomal mal-segregation (Battaglia et al., 1996; Gaulden, 1992; Lin and Keefe, 2004). Mutations controlling chromosome segregation have been described in maize, among them, the dv (Clark, 1940), ms43 (Golubovskaya and Sitnikova, 1980), ms28 (Golubovskaya and Distanova, 1986) and ms17 (Albertsen and Phillips, 1981) genes. The dv, ms28 and ms43 mutants cause failure of the spindle or impair disjunction mechanisms (Golubovskaya, 1979). Under the action of the dv gene (Clark, 1940; Staiger and Cande, 1990), the spindle fibers lie parallel or diverge instead of converging toward the poles (Caetano-Pereira and Pagliarini, 2001). This may lead to abnormal spindle orientation which might be inhibited by residual sewage sludge.
Staiger and Cande (1991) stated that all anomalies of chromosome segregation and cytokinesis are dependent on a delay of spindle fiber depolymerization (which in normal meiosis begins after anaphase I) affected by the ms28 gene and consequently leading to abnormal chromosome segregation (i.e., karyokinesis or abnormal chromosome movement) in division II.
Golubovskaya and Distanova (1986) suggested that the dv mutant affecting meiosis after metaphase I mainly disturbs the orientation of the two spindles relative to each other at meiosis II, leading to abnormal orientation of spindle fibers within the spindle, ending with abnormal spindle orientation.
The forces required for spindle assembly and movements of the spindle and chromosomes is initiated at Microtubule Organizing Centres (MTOCs) and have been attributed to microtubule dynamics, together with microtubule motors (Endow, 1999). In higher plant meiocytes, microtubules initially appear around prometaphase chromosomes, indicating a chromatin-mediated spindle assembly mechanism (Chan and Cande, 1998), however Mei mutations was found to disrupt the structure and function of spindle (Staiger and Cande, 1990; Staiger and Cande, 1991; Golubovskaya et al., 1992; Shamina et al., 1994; Mendes-Bonato et al., 2006).
Macieira-Coelho (2007) stated that disturbing the polarity of cells affects the orientation of the cell division plane. Accordingly, asymmetric division or overlapping cells in division II could occur. The presence of telophase cells in meiosis I and II with a protrusion at the position of cell plate may be due to the effect of the Preprophase Band (PPB) at the plane in which a cell will divide (PPB: microtubules in the outer cytoplasm become concentrated into a ring). These cortical microtubules oriented in the same plane as the future PPB, indicate cell division plane (Fig. 3). Division planes in the mutants appear random from the earliest stages of embryogenesis onward and hence the orientation of cell division is severely affected. The PPB is not the single decisive cue for orientation of the spindle; the spindle axis may rotate during division due to space constraints. So, a new division site may be chosen, resulting in a change of the orientation of cell division.
Abnormal cell contour and/or cell wall protrusion were detected in high percentages in meiocytes after 20 T/F of two residual sewage sludge treatment. This might be attributed to the drastic changes in deposition of uniform cell wall components leaving weak areas where protrusions occur and abnormal cell contour (Meijer et al., 1988).
The different kinds of aberrant meiotic end products (Fig. 2) were divided into three categories according to the number of microspores resulting from one meiocyte (diad and triad), arrangement of microspores within meiocytes wall (overlapped, decussate, linear, Apple, T, J, X, V -Shaped and abnormal cell orientation) and structural malformations as meiocytes having appendages and cytoplasmic protrusions. The percentage of aberrant meiotic end products increased by residual one than two additions (Table 2) and its value were more than thirty and eleven times than the control. When the meiotic end product aberrations were calculated from total end products only (normal tetrad stage) (Table 3), it was found that its percentage decreased by increasing residual sludge rates, this means that, 10 T/F, residual one addition caused the highest value.
According to Albertsen and Phillips (1981), the ms17 gene presents variable expression that affects spindle formation causing a variety of quartet abnormalities, including coenocytic quartets and structures with more or less than the normal four microspores. As a consequence, synchronous behaviour leads to the formation of a wide range of meiotic products, from diad, triads and tetrads with uneven-sized microspores (Risso-Pascotto et al., 2003; Reginaboldrini et al., 2006). Also, the total lack of a spindle may have been due to the absence of microtubules which are needed for spindle-fiber formation (Caetano-Pereira and Pagliarini, 2001).
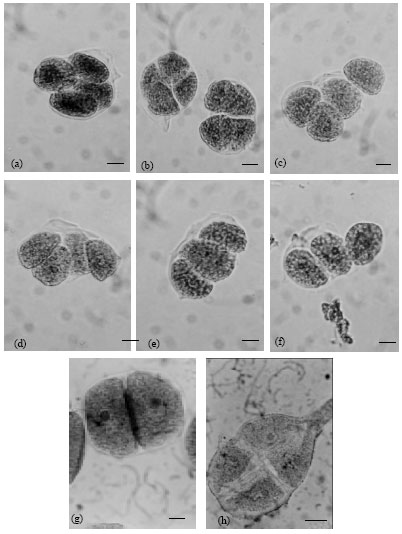 | |
| Fig. 2(a-h): | Abnormal meiotic end product of M2 Meiocytes of Zea mays. (a and b):different degrees of overlapping tetrads, (c): abnormal T-shape orientation, (d and e): different degrees of Linear tetrad, (f): Linear Triad, (g): Diad and (h): Tetrad with cytoplasmic protrusion |
In the mean time, the two cytoskeletal elements of spindle, microtubules and microfilaments, were found by Staiger and Cande (1991) to interact in a cell cycle-specific and site-specific fashion during higher plant meiosis leading to severa1 meiotic mutants known to have stage-specific disruptions in their microtubule arrays. Herein, abnormal cell contour, cell wall appendage, cytoplasmic protrusions and abnormal meiotic end products could be the result of a similar mutation. In addition, conditions of polyspory have been observed to depend on the direction of the formation and the number of cell plates resulting from several spindles (Caetano-Pereira and Pagliarini, 2001). Also, Golubovskaya and Distanova (1986) suggested that the presence of "dv" mutant mainly disturbs the orientation of the two spindles relative to each other at meiosis II leading to abnormal meiotic end product arrangements. This also may lead to the formation of abnormal arrangements of microspores within meiocyte wall as decussate, linear, T- J- X- Z-Apple- shape, overlapped and other abnormal cell orientation found in the present study.
Due to the effect of residual sewage sludge for the maturation, the pollen grains formation was inhibited after 10, 30 and 40T/F residual one addition and 10T/F residual two addition in the time of immature male tassels collection (~about eleven weeks old Zea).
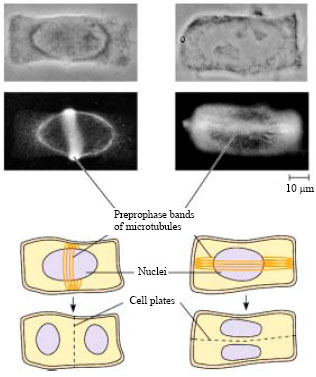 | |
| Fig. 3: | PPB predicts the orientation of cell division selected |
| Table 3: | The percentage of aberrant end products and the different kinds of aberrant M2 microspores meiocytes |
 | |
Pollen grain sterility indicated that the treatments of residual second additions ranged from 9-13%. However, no pollen grains were detected in the residual one addition except 6.23% in 20% T/F (Table 1). All previous values ranged from 3-8 times that of the control. The trend of the occurrence of aberrant meiotic end products was approximately similar to the percentage of pollen grain sterility. Inspite of that, the productivity, represented as kernel index, of these treatments were not significant difference when compared with the control (Amin et al., 2009a).
Generally, single metal or combination of metals have differential mode of action in biological system, since Metal-metal interaction plays a vital role in health and toxicity. Therefore, it is important to discuss mode of action of heavy metal(s) that is taking part in generating the genotoxicity in the present study.
Peijnenburg and Vijer (2007) recorded a variety of mechanisms exist for the detoxification of metals once they have entered an organism, precluding simple predictions of metal-induced toxicity on the basis of metal quotas, as occur with organic contaminants. Metals inside organisms can be present in the free ionic form or as soluble salt, or bound as follows:
| • | In the active center of functional proteins and low molecular weight peptides (as Zn finger proteins) |
| • | In the active center of enzymes (as cytochrome) |
| • | To transport proteins (as ferritin) |
| • | To metallothionine (MT) or other sequestration proteins |
| • | To cellular constituents causing dysfunction (as ion channels, DNA) |
The interactions among metals and between metals and plant are numerous and occur commonly during the processes of absorption and sequestration (Peijnenburg and Vijer, 2007).
Mode of action and toxicity: Most trace metals mediate gene expression by activation of single transduction pathways. Signal transduction is a progression by which information from an extracellular indication is transmitted into the cell through the plasma membrane and along an intracellular sequence of signaling molecules to excite a cellular response. The response may be the commencement of a gene transcription that occurs through various regulatory proteins that bind to specific DNA sequences in a gene. The net result of this binding is usually transcription of that gene and is referred to as transcriptional activation (Shanker, 2008).
Mode of action of Zn: The cytotoxic effects of zinc on plants are elucidated by Rout and Das (2003) as the major change was seen in the nucleus. If two metals bind to the same site of toxic action (such as competition of Zn2+ and Cd2+ for Ca2+ transport sites), it is possible to model the total metal bound to that site and hence, to predict metal toxicity using a mechanistic receptor binding approach in a concentration additive model. Alternatively, if two metals do not complete for the same binding site (such as binding of Zn2+ to Ca2+ transport sites vs. simultaneous binding of Cu2+ to Na2+ transport sites) then these models may provide more reliable estimates of individual metal bioavailability which then can be combined in more accurate response additive models (Peijnenburg and Vijer, 2007).
| • | Mode of action of Cd: Cell cycle progression, DNA replication and repair differentiation, proliferation and apoptotic pathway are altered and affected by cadmium. By activation cellular signals, it regulates cell cycle progression and inhibition of DNA methylation and/or interference with cell adhesion. Although, it enhances DNA synthesis and cell proliferation at lower concentration than 1 μM, Cd exposure above this concentration inhibits DNA synthesis and cell division (Yang et al., 2004). Zn-dependent proteins and Zn-binding molecules are “candidate” targets of Cd+2 toxicity. Cd2+ can replace Zn and so interfere with some of many reliant processes (Weber et al., 2006) |
| • | Mode of action of Lead: Possible mechanism for lead toxicity includes completion with and substitution for calcium, disturbance of calcium homeostasis. Calcium blocks the uptake of lead because it is a strong blocker of calcium channels. Disturbance of tissue oxidant/antioxidant balance and substitution or zinc in various Zn-mediated processes are some of the metabolic repercussions of lead toxicity (Ahmed et al., 2007). Therefore, lead can alter the regulation of genetic transcription through sequence-specific DNA-binding zinc sites in receptor channels (Langman and Kapu, 2006) |
| • | Mode of action of Copper: Copper can induce oxidative stress. Toxicity associated with Cu may be due to oxidative damage to biological macromolecules (Peijnenburg and Vijer, 2007) |
Therefore, due to sewage sludge production is continuously increasing in Egypt, the main current pressing needs are to find and develop more efficient sustainable technologies to allow a safe and suitable reuse of sewage sludge in agriculture. Furthermore, the legislations should be more adapted to Egyptian conditions as well as improvement of the institutional capacity to guarantee the enforcement of their application.
CONCLUSION
In sludge rule, it is stated that limits of the annual and cumulative loading of trace elements are adequate to protect against phytotoxicity and to prevent the accumulation of these elements in crops at levels harmful to consumers if it is applied in quantities commensurate with crop needs. Nevertheless, continued monitoring of trace elements in soils over longer time periods must be carried out. However, heavy metals and toxic compounds could be reached to higher levels in the course of time by repeated applications of sludge at agronomic rates and thus it could be inevitable to meet the irreversible problems of cumulative loading of trace elements and in any type of soil in long-term effect.
This work provides a bioassay to investigate the mutagenic properties of soil amended with sewage sludge. In the present study, the total percentage of aberrant pollen mother cells increased significantly with increasing sewage sludge concentration of residual one or two additions. A large number of genes acting from pre-meiosis to the post-meiotic mitosis could be affected, inducing mutations of these genes, may cause anomalies that impair plant fertility.
Therefore, to keep soils clean for future generations it is absolutely necessary to lower heavy metal limits in soils and organic materials.
REFERENCES
- Ahmed, M., M. Kaleem and J. Siddiqui, 2007. Environmental lead toxicity and nutritional factors. Clin. Nutr., 26: 400-408.
CrossRefDirect Link - Amin, A.W. and F.K. Sherif, 2001. Heavy metal contents in maize as affected by sewage sludge application morphological characters and yield. Pak. J. Biol. Sci., 4: 1451-1455.
CrossRefDirect Link - Amin, A.W., F.K. Sherif, H. El-Atar and H. Ez-Eldin, 2009. Effect of residual and accumulative sewage sludge on heavy metals bioaccumulation gene action and some yield parameters of Vicia faba. Res. J. Environ. Toxicol., 3: 60-75.
Direct Link - Amin, A.W., F.K. Sherif, H. El-Atar and H. Ez-Eldin, 2009. Residual effect of sewage sludge on soil and several yield parameters of Zea mays. Res. J. Environ. Toxicol., 3: 86-93.
CrossRef - Mendes-Bonato, A.B., M.S. Pagliarini1 and C. Borges do Valle, 2006. Abnormal spindle orientation during microsporogenesis in an interspecific Brachiaria (Gramineae) hybrid. Genet. Mol. Biol., 29: 122-125.
Direct Link - Banerjee, M.R., D.L. Burton and S. Depoe, 1997. Impact of sewage sludge application on soil biological characteristics. Agric. Ecosyst. Environ., 66: 241-249.
CrossRefDirect Link - Battaglia, D.E., P. Goodwin, N.A. Klein and M.R. Soules, 1996. Influence of maternal age on meiotic spindle assembly in oocytes from naturally cycling women. Hum. Reprod., 11: 2217-2222.
PubMed - Bhatt, A.M., C. Canales and H.G. Dickison, 2001. Plant meiosis: The means to 1N. Trends Plant Sci., 6: 114-121.
CrossRef - Caetano-Pereira, C.M. and M.S. Pagliarini, 2001. A new meiotic abnormality in Zea mays: Multiple spindles associated with abnormal cytokinesis in both divisions. Genome, 44: 865-871.
PubMed - Chan, A. and W.Z. Cande, 1998. Maize meiotic spindles assemble around chromatin and do not require paired chromosomes. J. Cell Sci., 111: 3507-3515.
Direct Link - Chaudhury, A.M., S. Craig, L. Farrell, K. Bloemer and E.S. Dennis, 1992. Genetic control of male-sterility in higher plants. Aust. J. Plant Physiol., 19: 419-425.
CrossRef - Clark, F.J., 1940. Cytogenetic studies of divergent meiotic spindle formation in Zea mays. Am. J. Bot., 27: 547-559.
CrossRef - Daniela, M.V., A.B.M. Bonato1, M.S. Pagliarini and C. Borges Do Valle, 2005. Abnormal meiotic behavior in Brachiaria brizantha (Poaceae) leading to microspore degeneration. Caryologia, 58: 396-402.
Direct Link - Diamond, G.L. and R.K. Zalups, 1998. Understanding renal toxicity of heavy metals. Toxicol. Pathol., 26: 92-103.
Direct Link - Endow, S.A., 1999. Microtubule motors in spindle and chromosome motility. Eur. J. Biochem., 262: 12-18.
PubMed - Fuerhacker, M. and T.D. Haile, 2011. Treatment and reuse of sludge, waste water treatment and reuse in the Mediterranean region. The Handbook Environ. Chem., 14: 63-92.
CrossRef - Gaulden, M.E., 1992. Maternal age effect: The enigma of Down syndrome and other trisomic conditions. Mutat. Res., 296: 69-88.
PubMed - Gaulden, M.E., 1987. Hypothesis: Some mutagens directly alter specific chromosomal proteins (DNA topoisomerase II and peripheral proteins) to produce chromosome stickiness, which causes chromosome aberrations. Mutagenesis, 2: 357-365.
PubMed - Golubovskaya, I., N.A. Avalkina and W.F. Sheridan, 1992. Effects of several meiotic mutations on female meiosis in maize. Develop. Genet., 13: 411-424.
CrossRef - Järup, L., 2003. Hazards of heavy metal contamination. Br. Med. Bull., 68: 167-182.
CrossRefDirect Link - Kaul, L.H. and T.G.K. Murthy, 1985. Mutant genes affecting higher plant meiosis. Theor. Applied Genet., 70: 449-466.
CrossRefDirect Link - Kizilkaya, R. and B. Bayrakli, 2005. Effects of N-enriched sewage sludge on soil enzyme activities. Applied Soil Ecol., 30: 192-202.
CrossRef - Knight, B.P., S.P. Mcgrath and A.M. Chaudri, 1997. Biomass carbon measurements and substrate utilization patterns of microbial populations from soils amended with cadmium, copper, or zinc. Applied Environ. Microbiol., 63: 39-43.
Direct Link - Kovaleva, O.A., 2008. Cytogenetic anomalies and causes for their occurrence in somatic cells. Cytol. Genet., 42: 48-59.
Direct Link - Kumar, G. and P. Rai, 2007. Comparative genotoxic potential of mercury and cadmium in soybean. Turk. J. Biol., 31: 13-18.
Direct Link - Lin, L. and D.L. Keefe, 2004. Nuclear origin of aging-associated meiotic defects in senescence-accelerated mice. Biol. Reprod., 71: 1724-1729.
CrossRef - McGill, M., S. Pathak and T.C. Hse, 1974. Effect of ethidium bromide on mitosis and chromosomes: A possible material basis for chromosome stickiness. Chromosoma, 47: 157-167.
CrossRef - Peijnenburg, W.J.G.M. and M.G. Vijer, 2007. Metal-specific interactions at the interface of chemistry and biology. Pure Applied Chem., 79: 2351-2366.
CrossRef - Reginaboldrini, K., M. Pagliarini and C. Dovalle, 2006. Abnormal timing of cytokinesis in microsporogenesis in Brachiariahumidicola (Poaceae: Paniceae). J. Genet., 85: 225-228.
Direct Link - Ricci, G.C.L., N. Silva, M.S. Pagliarini and C.A. Scapim, 2007. Microsporogenesis in inbred line of popcorn (Zea mays L.). Genet. Mol. Res., 6: 1013-1018.
Direct Link - Risso-Pascotto, C., M.S. Pagliarini and C. Borges do Valle, 2003. A mutation in the spindle checkpoint arresting meiosis II in Brachiaria ruziziensis. Genome, 46: 724-728.
CrossRef - Rout, G.R. and P. Das, 2003. Effect of metal toxicity on plant growth and metabolism: I. Zinc. Agronomie, 23: 3-11.
CrossRefDirect Link - Sastre, I., M.A. Vicente and M.C. Lobo, 1996. Influence of the application of sewage sludges on soil microbial activity. Bioresource Technol., 57: 19-23.
CrossRef - Sathawara, N.G., D.J. Parikh and Y.K. Agarwal, 2004. Essential heavy metals in environmental samples from Western India. Bull. Environ. Contam. Toxicol., 73: 756-761.
CrossRefPubMedDirect Link - Sharma, A. and G. Talukder, 1987. Effects of metals on chromosomes of higher organisms. Environ. Mol. Mutagen., 9: 191-226.
CrossRefDirect Link - Singh, R.P. and M. Agrawal, 2010. Effect of different sewage sludge applications on growth and yield of Vigna radiata L. field crop: Metal uptake by plant. Ecol. Eng., 36: 969-972.
CrossRef - De Lourdes Marzo Solano, M., P.L. de Lima, J.F. Luvizutto, P.R. Silva, G. de Aragao Umbuzeiro and J.L. de Camargo, 2009. In vivo genotoxicity evaluation of a treated urban sewage sludge sample. Mutat. Res., 676: 69-73.
PubMedDirect Link - Staiger, C.J. and W.Z. Cande, 1990. Microtubule distribution in dv, a maize meiotic mutant defective in the prophase to metaphase transition. Dev. Biol., 138: 231-242.
CrossRef - Staiger, C.J. and W.Z. Cande, 1991. Microfilament distribution in maize meiotic mutants correlates with microtubule organization. Plant Cell, 3: 637-644.
CrossRef - Tam, N.F.Y. and Y.S. Wong, 1990. The comparison of growth and nutrient removal efficiency of Chlorella pyrenoidosa in settled and activated sewages. Environ. Pollut., 65: 93-108.
CrossRef - Yang, P.M., S.J. Chiu, K.A. Lin and L.Y. Lin, 2004. Effect of cadmium on cell cycle progression in Chinese hamster ovary cells. Chem. Biol. Interact., 149: 125-136.
CrossRef - Sang, N. and G. Li, 2004. Genotoxicity of municipal landfill leachate on root tips of Vicia faba. Mutation Res., 560: 159-165.
CrossRef - Weber, M., A. Trampczynska and S. Clemens, 2006. Comparative transcriptome analysis of toxic metal responses in Arabidopsis thaliana and the Cd2+-hypertolerant facultative metallophyte Arabidopsis halleri. Plant Cell Environ., 29: 950-963.
CrossRef