
A.C.C. Egbuonu
Nutrition and Toxicological Biochemistry Unit, Department of Biochemistry, University of Nigeria, Nsukka, Nigeria
L.U.S. Ezeanyika
Nutrition and Toxicological Biochemistry Unit, Department of Biochemistry, University of Nigeria, Nsukka, Nigeria
P.M. Ejikeme
Industrial Chemistry Unit, Department of Pure and Industrial Chemistry,University of Nigeria, Nsukka, Nigeria
O. Obidoa
Nutrition and Toxicological Biochemistry Unit, Department of Biochemistry, University of Nigeria, Nsukka, Nigeria
Research Journal of Environmental Toxicology
Year: 2010 | Volume: 4 | Issue: 4 | Page No.: 205-213







ABSTRACT
Adult male Wistar rats were fed with arginine (ARG) (60 mg kg¯1 b.wt.), glutamate (GLU) (90 mg kg¯1 b.wt.), monosodium glutamate (MSG) (15 mg kg¯1 b.wt.), ARG+GLU (60:90 mg kg¯1 b.wt.) or ARG+MSG (60:15 mg kg¯1 b.wt.). The feeding was by oral intubation and was daily for 28 days. The aim was to investigate possible alterations in the serum biochemistry and liver histology induced by ARG, GLU or MSG either alone or in such combinations as ARG+GLU or ARG+MSG. After 28 days oral treatment, rats treated with ARG, GLU or MSG alone significantly (p<0.05) increased the serum alkaline aminotransferase (ALT) activity, whereas rats co-treated with ARG and either GLU or MSG decreased (p<0.05) the serum ALT activity. In comparison with the control, the other treatment groups elicited a significant quantitative increase in the serum aspartate aminotransferase (AST) activity and the computed AST:ALT ratio but, the observed increase in the ARG-treated group was consistently lower relative to the other groups. Liver sections of rats from the various treatment groups showed varying degrees of focally diffuse (random) hepatocellular necrosis. On the whole, the study suggests that sub-acute daily oral treatment of male Wistar rats with ARG, GLU or MSG alone or ARG together with GLU or MSG could possibly impair their physiological functions as evidenced in the significant alterations in the serum biochemistry and liver histology of this study. The results are considerably significant going by the possible inadvertent abuse of ARG, GLU and MSG alone or together in diets and drugs.
PDF Abstract XML References Citation
Received: April 09, 2010;
Accepted: May 31, 2010;
Published: August 05, 2010
How to cite this article
A.C.C. Egbuonu, L.U.S. Ezeanyika, P.M. Ejikeme and O. Obidoa, 2010. Histomorphologic Alterations in the Liver of Male Wistar Rats Treated with L-Arginine Glutamate and Monosodium Glutamate. Research Journal of Environmental Toxicology, 4: 205-213.
DOI: 10.3923/rjet.2010.205.213
URL: https://scialert.net/abstract/?doi=rjet.2010.205.213
DOI: 10.3923/rjet.2010.205.213
URL: https://scialert.net/abstract/?doi=rjet.2010.205.213
INTRODUCTION
The inter-conversion of amino acids and also the metabolism of most amino acids occur to a significant extent in the liver (Mayes and Bender, 2003). Thus, it is conceivable that any untoward consequence of amino acids intake may also manifest in the form of liver damage.
Arginine, a semi essential amino acid, is an important component in the human diet that is abundant in natural foods including wheat flour, diary products, pork, nuts and seeds. The ARG may have therapeutic effect through its major metabolite nitric oxide, No. Also, GLU is an amino acid. Like ARG, GLU is abundant in common foods and could play a role in the synthesis of NO (Moncada et al., 1991). Despite the promising therapeutic potentials of ARG through its metabolite, NO, results from recent studies have implicated ARG for causing significant weight loss (Sepehri et al., 2006), increasing systemic blood pressure (at 4 mg mL-1 in drinking water) (Nematbakhsh et al., 2008) and exacerbating (at 20 mg kg-1) the MSG-induced adverse effects (Egbuonu et al., 2010) in rats. Furthermore, although NO plays an important role in the host defense mechanism, excess or deficiency of NO could lead to pathological conditions, including hypertension, diabetes, tissue damage and inflammation (Lokhande et al., 2006).
Glutamate (GLU), a natural flavor enhancer, is also used in the form of its variant, monosodium glutamate (MSG). In particular, GLU in the form of MSG is used the world over as flavor enhancer. Recently, Praputpittaya and Wililak (2003) reported that MSG treated rats experienced dose-dependent deficit in visual performance which may reflect impairment of visual organs and brain function. Furthermore, Bergen et al. (1998) reported MSG-induced elevation in feed intake, which may lead to obesity. And obesity was associated with the development of diabetes and impaired glucose metabolism in male mice (Zhou and Sato, 2008). Reported cases of MSG-induced side effects as reviewed by Egbuonu et al. (2009a), include, but are not limited to, the alteration in the levels of carbohydrates, lipids and proteins in rats as well as the Chinese restaurant syndrome. Furthermore, possible MSG-induction of deleterious response on the prostate and other major organs with high metabolic activities, especially the liver was reported recently (Egbuonu et al., 2010) and GLU (as MSG) was shown to decrease the monoamines consequent upon MSG-induced neuro-toxic effect in rats (Abeer Waggas, 2009). Also, Eweka and Om'Iniabohs (2007) studied the histological effects of MSG alone on the cerebellum of adult Wistar rats. They found out that the treated section of the cerebellum showed disruption of the Purkinje and granular layers, sparse granular cell disruption cellular degenerative changes in the granular layer and inferred that MSG consumption may have some deleterious effects on the cerebellum of adult Wistar rats which may lead to tremor and ataxia.
The ARG, GLU and MSG are readily available either in natural foods or in packaged foods. Therefore, since ARG together with GLU or MSG may be present in the human diet and with the above highlighted possible adverse effects of these agents, it is necessary to study the possible effects of their single or combined intake on the liver. Thus, the effect of ARG (60 mg kg-1 b.wt), GLU (90 mg kg-1 b.wt.) or MSG (15 mg kg-1 b.wt.) alone or ARG together with either GLU or MSG on the serum activity of ALT (a specific bio-marker of hepatocellular damage), AST (a non-specific bio marker of hepatocelluar damage) as well as on the computed AST:ALT ratio and the histology of the liver sections of the rats were investigated in the present study. The choice of treatment dose was based on earlier reports (Olney, 1969; Barbul et al., 1981; Marshal, 1994; Alexander et al., 2004; Onyema et al., 2006).
MATERIALS AND METHODS
Chemicals
Ajinomoto brand of MSG was purchased from a regular foodstuff market at Nsukka. The ARG and GLU were obtained from Sigma Chemical Co., St. Louis, Mo. USA through the chemical store of Biochemistry Department, University of Nigeria, Nsukka. Other chemicals used were of certified analytical grade. The study was conducted between June, 2005 and June, 2006.
Animals and Treatment
Adult male albino rats procured from the animal house of the Faculty of Biological Sciences, University of Nigeria, Nsukka were used in this study. All the animals received humane care in accordance with the guidelines of the National Institute of Health, USA for ethical treatment of laboratory animals.
Twenty four adult male albino rats having mean Body Weight (BW) of 93±0.5 g were normalized for 1 week and then randomly assigned to six groups of four rats each. Groups II, III and IV rats were fed ARG (60 mg kg-1 b.wt.), GLU (90 mg kg-1 b.wt.) and MSG (15 mg kg-1 b.wt.), respectively while group V rats were fed ARG with GLU (60:90 mg kg-1 b.wt.) whereas group VI rats were fed ARG with MSG (60:15 mg kg-1 b.wt.). Group I rats were given distilled water (1 mL kg-1 of BW). Treatment was by daily oral incubations and lasted for 28 days. The rats were housed in stainless steel cages at room temperature (28±2°C) and exposed to a normal daylight/dark cycle under humid tropical condition. They were supplied with enough rat feed and drinking (tap) water ad libitum throughout the duration of the experiment.
Sample Collection and Preparation
The animals were sacrificed 24 h after the 28 days oral treatment. Their blood samples were collected individually with sterile capillary tubes as already described (Egbuonu et al., 2009a). The liver specimens were promptly excised from the sacrificed rats and fixed in 10% formaldehyde buffered saline (formal saline).
The blood samples thus collected were allowed to clot and centrifuged at 3000 rpm for 10 min. The resultant sera were collected individually in stoppered polystyrene tubes and stored in a deep freezer for the determination of the serum alanine aminotransferase (ALT) and aspartate aminotransferase (AST) activities.
The liver specimens were dehydrated in graded levels of alcohol (about 70-100%) in ascending order to remove the water content. After the dehydration, the tissues were cleared in xylene impregnated with paraffin wax and sectioned at 5 microns thickness using rotary microtone. The sections were floated on a water bath maintained at a temperature of 2-3°C below melting point of the paraffin wax after which the sections were dried on a hot plate maintained at a temperature of 2-3°C above the melting point of the paraffin wax used. After proper drying, the sections were stained and mounted using haematoxylin and eosin.
Determination of the Serum Alkaline Aminotransferase (ALT) Activity
The serum alkaline aminotransferase (ALT) activity was assayed by the method of Reitman and Frankel (1957). The method is based on the principle that pyruvate (pyruvic acid) formed from the alanine aminotransferase catalysed reaction between α–ketoglutarate (oxoglutarate) and L-alanine is coupled with chromogen solution (2, 4-dinitrophenyl hydrazine) in alkaline medium to form colored hydrazone the concentration of which is proportional to the alanine aminotransferase activity as measured with a colorimeter.
To 0.05 mL of each serum sample in a test tube was added 0.25 mL of buffer/substrate solution. This was incubated at 37°C for 30 min in a water bath followed by the addition of 0.25 mL of chromogen solution. The content was mixed and allowed to stand for 20 min at room temperature. Then 2.5 mL of sodium hydroxide (0.4N) was added and mixed. The absorbance was read after 5 min against the blank at 540 nm. The blanks were treated as the samples but without the addition of chromogen solution used to stop all the enzymatic reactions. ALT activity in IU L-1 was read off from the standard curve.
Determination of the Serum Aspartate Aminotransferase (AST) Activity
The serum aspartate aminotransferase (AST) activity was assayed by the method of Reitman and Frankel (1957). The method is based on the principle that oxaloacetate (oxaloacetic acid) formed from the aspartate aminotransferase catalysed reaction between alpha–ketoglutarate and L-aspartate is coupled with chromogen solution (2, 4-dinitrophenyl hydrazine) in alkaline medium to form colored hydrazone and that the concentration of which is proportional to the alanine aminotransferase activity as measured with a colorimeter.
To 0.05 mL of each serum sample in a test tube was added 0.25 mL of buffer/substrate solution. This was incubated at 37°C for 30 min in a water bath followed by the addition of 0.25 mL of chromogen solution. The content was mixed and allowed to stand for 20 min at room temperature. Then 2.5 mL of sodium hydroxide (0.4 N) was added and mixed. The absorbance was read after 5 min against the blank at 540 nm. The blanks were treated as the samples but without the addition of chromogen solution used to stop all the enzymatic reactions. The ALT activity in IU L-1 was read off from the standard curve.
Computation of the Serum Aspartate Aminotransferase to Alanine Aminotransferase (AST:ALT) Ratio
We computed the serum AST:ALT ratio from the results of the serum AST and ALT activities of the present study.
Statistical Analysis
The data obtained from the present study were expressed as Mean±SEM. They were analyzed by one-way analysis of variance (ANOVA) as earlier described (Egbuonu et al., 2009a). In brief, multiple comparisons of means were made using the Least Significant Difference (LSD) test with the Statistical Package for Social Sciences (SPSS) for Windows version 11.0 package. Differences were considered significant at p<0.05 level of significance.
RESULTS
Serum ALT Activity
The determination of the serum ALT activity as shown in Table 1 revealed that rats that were fed either ARG, GLU or MSG alone significantly (p<0.05) increased the serum ALT activity whereas the rats that were fed ARG together with either GLU or MSG decreased (p<0.05) the serum ALT activity. Furthermore, the serum ALT activity increased (p<0.05) most (27.94% or 103.21±0.14 IU L-1) in the MSG-treated rats (Group IV) but decreased (p<0.05) most (93.19 % or 5.50±0.19 IU L-1) in the rats given a combination of ARG and GLU (Group V).
| Table 1: | The effect of distilled water, arginine (ARG), glutamate (GLU), monosodium glutamate (MSG), arginine plus glutamate (ARG+GLU) and arginine plus monosodium glutamate (ARG+MSG) on the serum alanine aminotransferase (ALT) activity |
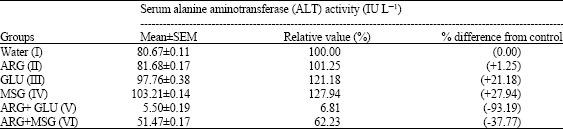 | |
| The results are Mean±SEM for four rats in each group and are significantly different from control and between groups (p<0.05) value in bracket is the percentage increase (+) or decrease (-) relative to the control | |
| Table 2: | The effect of distilled water, arginine (ARG), glutamate (GLU), monosodium glutamate (MSG), arginine plus glutamate (ARG+GLU) and arginine plus monosodium glutamate (ARG+MSG) on the serum aspartate aminotransferase (AST) activity |
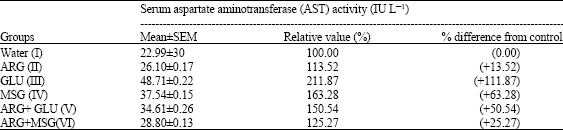 | |
| The results are Mean±SEM for four rats in each group and are significantly different from control and between groups (p<0.05) value in bracket is the percentage increase (+) relative to the control | |
| Table 3: | The effect of distilled water, arginine (ARG), glutamate (GLU), monosodium glutamate (MSG), arginine plus glutamate (ARG+GLU) and arginine plus monosodium glutamate (ARG+MSG) on the serum aspartate aminotransferase to alanine aminotransferase (AST:ALT) ratio |
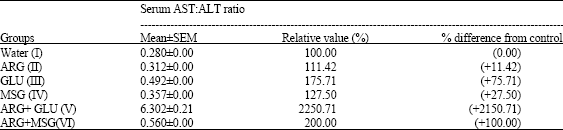 | |
| The results are mean ± SEM for four rats in each group and are significantly different from control and between groups (p<0.05) value in bracket is the percentage increase (+) relative to the control | |
Serum AST Activity
The results shown in Table 2 revealed a significant elevation in the serum AST activity of rats in the various treatment groups relative to rats in the control group. The highest quantitative rise (above one-fold) in the serum AST activity was noted in the GLU-fed rats (Group III) followed by MSG-treated group of rats. Data indicated the least increase in the serum AST activity in the group of rats that were fed ARG alone.
Serum AST:ALT Ratio
When compared with the control group of rats, the serum AST:ALT ratio (computed from the results of the AST and ALT activities obtained from the present study) showed a significant increase in rats from the various treatment groups. The least increase in the serum AST:ALT ratio was observed in the ARG-treated group of rats (Group II) whereas a marked increase in the serum AST:ALT (that is above 1) was recorded in the rats that were ingested simultaneously with ARG and GLU (Group V) (Table 3).
Histopathological Changes in the Liver Sections
The histomorphological changes in the liver section of the different groups of treated rats were characterized by degenerative/necrotic and inflammatory changes as evidenced by lesions (Fig. 2). Liver sections of the control (Group I) group of rats showed typical lobular histology, with hepatocytes radially arranged around a central vein (Fig. 1). Sections collected from rats treated with ARG (Group II) were mildly hyperaemic with moderate sinusoidal oedema. There was also mild mononuclear leucocytes infiltration of the portal areas.
Rats treated with GLU (Group III) showed moderate to severe vacuolar infiltration of ductular epithelium lining cells with mild to moderate infiltration of mononuclear leucocytes into the portal area and mild to moderate focally diffuse (random) hepatocellular necrosis; (apoptotic) (Fig. 2).
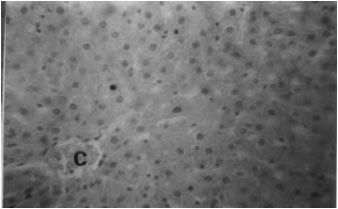 | |
| Fig. 1: | Liver section of untreated, control (Group I) rat showing central vein (C) surrounded by cords of hepatocytes. H and E stains, x400 |
 | |
| Fig. 2: | Liver section of rat treated with GLU (Group III) showing portal area with hepatic artery (A) and portal vein (P). Note the bile duct (B) with degeneration and necrosis of epithelia cells and apoptotic hepatocytes (arrow heads). H and E stains x400 |
Rats treated with MSG (Group IV) showed lesions, similar to, but less severe when compared to what was described above for Group III rats. Liver sections from rats treated with ARG+GLU (Group V) and ARG+MSG (Group VI) were moderately haemmorhagic and with moderate haemmosidinosis. There was mild to moderate necrosis of the hepatocytes in the periportal areas and mild fibroplasia with mononuclear leucocytic infiltration.
DISCUSSION
The ALT enzyme is abundant in the liver and is released into the blood stream following hepatocellular damage. Thus, it is a sensitive marker of liver damage (Al-Mamary, 2002) and the marked increase in the serum ALT activity observed in the MSG-fed rats followed by the GLU-fed rats in the present study might be indicating liver damage. The metabolism of most amino acids and their derivatives occur to a significant extent in the liver (Mayes and Bender, 2003) and essentially involves deamination to produce ammonium ion (NH4+) that could be toxic unless made less toxic via the reactions of the urea cycle. The sodium moiety in MSG could easily dissociate to yield free GLU. Thus, the possible ammonium ion overload that may occur with GLU or MSG intake could damage the liver, consequently releasing the ALT enzyme, hence its observed elevation in the serum. The result is basically comparable with that of previous works by Farombi and Onyema (2006) and Onyema et al. (2006). In particular, the result appears to be supported by the earlier report of Egbuonu et al. (2009b) that MSG treatment increased the serum ALT activity in male albino rats due to possible ammonium ion overload resulting from an increased level of glutamate.
The relatively low increase in the serum ALT activity (1.25%) observed in the ARG-fed rats appears to suggest an attempt by ARG to counteract possible liver damage. The marked decrease in the serum ALT activity observed in rats that were fed a combination of ARG with GLU (Group V) followed by those that were fed ARG together with MSG seems to suggest that ARG co-treatment with either GLU or MSG probably antagonized the possible hepatotoxic effects induced by either GLU or MSG alone. This may further suggest that a combination of ARG with either GLU or MSG could ameliorate the possible hepatotoxic effect of either GLU or MSG alone. The result, however, was at variance with the earlier report by Egbuonu et al. (2010) that ARG at 20 mgkg-1 b.wt. significantly exacerbated possible adverse effects induced by MSG (5 mg kg-1 b.wt.) in rats. Probably, the differing concentration used in the separate works accounted for this variance.
The significant and quantitative rise in the serum AST activity of rats in the various treatment groups could be reflective of possible treatment-induced damage to the metabolically active organs including the liver, brain, heart and lungs (Egbuonu et al., 2010) since AST enzyme, contrary to ALT enzyme, is not a specific marker of organ damage. The intake of GLU alone seems to be more potent in eliciting such damage followed by that of MSG alone and then ARG together with GLU.
Recently, Egbuonu et al. (2010) highlighted the biochemical/diagnostic importance of AST:ALT ratio by excluding liver damage as the likely cause for an increased serum AST activity when the AST:ALT ratio is greater than one and vice versa. Thus, the observed elevation in the computed serum AST:ALT ratio of the ARG plus GLU-treated rats by 6,302±0.21 appears to implicate damage to other high metabolic organs (apart from the liver) as the source of the marked increase in the AST activity following the concomitant ingestion of ARG and GLU in male rats. This is seemingly supported by the marked reduction by 93.19% in the ALT activity of the ARG plus GLU-fed rats reported in the present study.
Histological changes were the most consistent of the changes that could be identified due to toxic processes and, changes in the histology of organs were frequently used to collaborate physiological states of the organs as adduced from the results of the biochemical assays. Thus, the histology of the liver sections from rats in the different treatment groups that were characterized by degenerative/necrotic and inflammatory changes might be indicative of onset of toxic process/stimulus in the liver (Butler, 1996). The general picture was such that treating rats with GLU or MSG alone, or in combination with ARG caused variable degree of hepatocytes necrosis in the periportal areas which appeared to be related to ductular lesions that might have been directly or indirectly related to bile toxicity (histopathologist’s personal communication).
In the present study, the recurring hepatic histology (though of varying degrees) of rats in the various treatment groups were random hepatocellular necrosis in the periportal areas and fibroplasia with mononuclear leucocytic infiltration. These were most likely the result of direct damage exerted by these agents on the hepatocytes. The observed necrosis might lead to hepatitis (Nwaopara et al., 2007) hence the infiltration of mononuclear leucocytes into the portal area was, probably, an adaptive cellular response to possible hepatitis resulting from the toxic effects of the test agents.
In particular, the MSG-induced lesions are in consonance with the reported adverse liver histology induced by MSG in mice (Nakanishi et al., 2008) and rats (Eweka and Om'Iniabohs, 2008). The mild hyperaemic and moderate sinusoidal oedema noted in the liver sections of ARG-fed rats apparently supports the very slight increase in their serum ALT and AST activities and AST:ALT ratio as observed in the present study. Further to this, it apparently agrees with earlier finding by Angele et al. (2002) that ARG prevented liver oedema in trauma-hemorrhage-induced liver injury in rats. However, the observation of a less severe lesion in the rats treated with MSG (Group IV) when compared to what was described for the GLU-fed rats (Group III) seems to be at variance with the increase in the serum ALT activity that was observed to be higher in the Group IV rats than in the Group III rats. The apparent significant deviation is partly accounted for by the serum AST activity that increased markedly in the GLU-fed than in the MSG-fed rats due, probably, to liver damage as adduced from the calculated AST:ALT ratio of less than one (Egbuonu et al., 2010).
On the whole, the study suggests that sub-acute daily oral treatment of male Wistar rats with ARG, GLU and MSG alone or ARG together with GLU or MSG could elicit significant biochemical and histological alterations in the liver, which might eventually lead to the impairment of liver functions. In addition, treating male rats with ARG and GLU together may have damaged other high metabolic organs apart from the liver. These results are of considerable significance going by the widespread usage and possible inadvertent abuse of ARG, GLU and MSG alone or together in diets and drugs. Research to evaluate the public health implication of these agents in the initiation, progression and management of liver damage is in progress.
ACKNOWLEDGMENT
The authors gratefully acknowledge Dr. S.V.O. Shoyinka of the Department of Veterinary Pathology, University of Nigeria, Nsukka for his kind assistance during the histopathologic studies.
REFERENCES
- Waggas, A.M., 2009. Neuroprotective evaluation of extract of ginger (Zingiber officinale) root in monosodium glutamate-induced toxicity in different brain areas of male albino rats. Pak. J. Biol. Sci., 12: 201-212.
CrossRefPubMedDirect Link - Alexander, B.T., M.T. Llinas, W.C. Kruckeberg and J.P. Granger, 2004. L-Arginine attenuates hypertension in pregnant rats with reduced uterine perfusion pressure. Hypertension, 43: 832-836.
CrossRefPubMedDirect Link - Al-Mamary, M., M. Al-Habori, A.M. Al-Aghbari and M.M. Baker, 2002. Investigation into the toxicological effects of Catha edulis leaves: A short term study in animals. Phytother. Res., 16: 127-132.
CrossRefPubMedDirect Link - Angele, M.K., F. Fitzal, N. Smail, M.W. Knoferl and M.G. Schwacha et al., 2002. L-arginine attenuates trauma-hemorrhage-induced liver injury. Crit. Care Med., 28: 3242-3248.
PubMed - Barbul, A., D.A. Sisto, H.L. Wasserkrug and G. Efron, 1981. Arginine stimulates lymphocytes immune response in healthy human. Surgery, 90: 244-251.
PubMed - Bergen, H.T., T.M. Mizuno, J. Taylor and C.V. Mobbs, 1998. Hyperphagia and weight gain after gold-thioglucose: Relation to hypothalamic neuropeptide Y and proopiomelanocortin. Endocrinology, 139: 4483-4488.
CrossRefDirect Link - Butler, W.H., 1996. A review of the hepatic tumors related to mixed-function oxidase induction in the mouse. Toxicol. Pathol., 24: 484-492.
CrossRef - Egbuonu, A.C.C., O. Obidoa, C.A. Ezeokonkwo, L.U. Ezeanyika and P.M. Ejikeme, 2009. Low dose oral administration of monosodium glutamate in male albino rats may be nephroprotective. Bio-Research, 7: 470-473.
CrossRefDirect Link - Egbuonu, A.C.C., O. Obidoa, C.A. Ezeokonkwo, L.U.S. Ezeanyika and P.M. Ejikeme, 2009. Hepatotoxic effects of low dose oral administration of monosodium glutamate in male albino rats. Afr. J. Biotechnol., 8: 3031-3035.
Direct Link - Egbuonu, A.C.C., C.A. Ezeokonkwo, P.M. Ejikeme, O. Obidoa and L.U.S. Ezeanyika, 2010. Some biochemical effects of sub-acute oral administration of L-arginine on monosodium glutamate-fed wistar albino rats 2: Serum alkaline phosphatase, total acid phosphatase and aspartate aminotransferase activities. Asian J. Biochem., 5: 89-95.
CrossRefDirect Link - Eweka, A.O. and F.A.E. OmIniabohs, 2007. Histological studies of the effects of monosodium glutamate on the cerebellum of adult Wistar rats. Internet J. Neurol., 8: 68-72.
Direct Link - Eweka, A.O. and F.A.E. Om'Iniabohs, 2008. Histological studies of the effects of monosodium glutamate on the liver of adult Wistar rats. Internet J. Gastroenterol.
Direct Link - Lokhande, P.D., B.S. Kuchekar, A.R. Chabukswar and S.C. Jagdale, 2006. Nitric oxide: Role in biological system. Asian J. Biochem., 1: 1-17.
CrossRefDirect Link - Moncada, S., R.M.J. Palmer and E.A. Higgs, 1991. Nitric oxide: Physiology, pathophysiology and pharmacology. Pharmacol. Rev., 43: 109-142.
PubMedDirect Link - Nakanishi, Y., K. Tsuneyama, M. Fujimoto, T.L. Salunga and K. Nomoto et al., 2008. Monosodium Glutamate (MSG): A villain and promoter of liver inflammation and dysplasia. J. Autoimmun., 30: 42-50.
CrossRef - Nematbakhsh, M., Z. Heydarzadeh, L. Borjian and S. Haghjooyejavanmard, 2008. Low dose of l-arginine does not change endothelial permeability of aorta and coronary arteries in rat. Pak. J. Nutr., 7: 126-129.
CrossRefDirect Link - Nwaopara, A.O., M.A.C. Odike, U. Inegbenebor and M.I. Adoye, 2007. The combined effects of excessive consumption of ginger, clove, red pepper and black pepper on the histology of the liver. Pak. J. Nutr., 6: 524-527.
CrossRefDirect Link - Olney, J.W., 1969. Brain lesions, obesity and other disturbances in mice treated with monosodium glutamate. Science, 164: 719-721.
CrossRefDirect Link - Onyema, O.O., E.O. Farombi, G.O. Emerole, A.I. Ukoha and G.O. Onyeze, 2006. Effect of vitamin E on monosodium-glutamate induced hepatoxicity and oxidative stress in rats. Indian J. Biochem. Biophys., 43: 20-24.
PubMed - Praputpittaya, C. and A. Wililak, 2003. Visual performance in monosodium L-glutamate-treated rats. Nutr. Neurosci., 6: 301-307.
CrossRef - Reitman, S. and S. Frankel, 1957. A colorimetric method for the determination of serum glutamic oxalacetic and glutamic pyruvic transaminases. Am. J. Clin. Pathol., 28: 56-63.
CrossRefPubMedDirect Link - Sepehri, G., S. Vahid, B. Fariba and F. Rasoul, 2006. Effect of L-NAME/L-arginine microinjection into nucleus accumbens shell on morphine withdrawal signs in male rats. Int. J. Pharmacol., 2: 171-176.
CrossRefDirect Link - Zhou, W. and K. Sato, 2008. Physiological vulnerability to diet induced obesity in inbred alloxan-induced diabetes-susceptible mice. J. Boil. Sci., 8: 421-425.
CrossRefDirect Link - Farombi, E.O. and O.O. Onyema, 2006. Monosodium glutamate-induced oxidative damage and genotoxicity in the rat: Modulatory role of vitamin C, vitamin E and quercetin. Hum. Exp. Toxicol., 25: 251-259.
CrossRefPubMedDirect Link