
Emad El-Zayat
Department of Zoology, Faculty of Science, Cairo University, Giza, Egypt
Sayed Rawi
Department of Zoology, Faculty of Science, Cairo University, Giza, Egypt
Nashwa Ismail
National Organization for Drug Control and Research (NODCAR), Giza, Egypt
Research Journal of Environmental Toxicology
Year: 2008 | Volume: 2 | Issue: 1 | Page No.: 35-52







ABSTRACT
The present study was designed to investigate the therapeutic effect of selenium on delayed deltamethrin (DM) neurotoxicity. Animals under investigation were treated with a single acute dose of deltamethrin (¼ LD50 = 111.25 mg/100 g body weight). The therapeutic effect of selenium has been evaluated by administration of a daily dose either individually or in antioxidant preparation (Antox) for a period of three weeks. Acute deltamethrin intoxication induced marked alterations in the brain and blood levels of monoamines and free amino acids as well as acetylcholinesterase activity. Selenium supplementation, either individually (DMS) or in antioxidant preparation (DMA), showed a considerable ability to attenuate some but not all signs of delayed deltamethrin neurotoxicity. Among the two Se-supplemented groups, it was difficult to see a clear therapeutic effect on all measured parameters. DMS showed the best results in some cases but DMA showed the best results in others reflecting different mechanisms of action. These therapeutic effects seemed to be time-dependent.
PDF Abstract XML References Citation
How to cite this article
Emad El-Zayat, Sayed Rawi and Nashwa Ismail, 2008. Delayed Effects of Acute Deltamethrin Toxicity on Brain and Blood Monoamines and Free Amino Acids: Therapeutic Role of Selenium-Supplementation. Research Journal of Environmental Toxicology, 2: 35-52.
DOI: 10.3923/rjet.2008.35.52
URL: https://scialert.net/abstract/?doi=rjet.2008.35.52
DOI: 10.3923/rjet.2008.35.52
URL: https://scialert.net/abstract/?doi=rjet.2008.35.52
INTRODUCTION
Humans are constantly exposed to free radicals created by internal cellular metabolism or by exposure to environmental pollution (Gilgun-Sherki et al., 2002). The recent wide spread use of pesticides in agriculture and public health programs increased the risk of exposure to pesticides and in turn to more reactive oxygen species. Antioxidants have recently attracted attention because of their ability to quench free radical formation before it damages cells or tissues. According to Khedr et al. (1999), the more free radicals present, the greater the need for antioxidants in our daily life.
Previous studies have accumulated data suggesting that pyrethroids; widely used insecticides; produce oxidative stress through the generation of free oxygen radicals (Kale et al., 1999a, b; Sayeed et al., 2003; Parvez and Raisuddin, 2006; Rehman et al., 2006; Li et al., 2007) and play a role in the neuro-degenerative process leading to some neural diseases (Theophilidis et al., 1997; He, 2000; Karen et al., 2001; Soderlund et al., 2002).
Antioxidants like vitamin E, zinc and copper have shown to play a protective role against the pyrethroid-induced oxidative stress (Kale et al., 1999a, b; Aldana et al., 2001; El-Zayat et al., 2005; Parvez and Raisuddin, 2006). On the other hand, selenium has demonstrated a great potential in maintenance of human health and proved to be important in the biotransformation and detoxification of many toxic materials (Shoka et al., 1994). Kumar et al. (2001) reported normal appearance of animals within 2 weeks, after receiving selenium and vitamin E, when they beard paralysis as a side effect of Butox treatment (deltamethrin commercial preparation for veterinary use). Previous reports have also revealed the protective role of selenium against toxicity with carbosulfan pesticide (El-Manakhly, 1996; El-Zarkouny et al., 1999).
Alterations of the neurotransmitters and their metabolites have been used as a measure of neurotoxicity. Several lines of evidence indicate that type II pyrethroids are strong inducers for release of catecholamines (Cremer and Seville, 1982; Bradburry et al., 1983; Ray, 1991). In addition, preliminary reports of Rawi (1984) and Chugh et al. (1992) indicated that pyrethroid insecticides affect some amino acid-neurotransmitters, suggesting that the excitatory amino acids are the target for pyrethroid neurotoxicity.
The present research aimed to investigate the therapeutic effect of selenium in single or combined form on delayed neurotoxic effects due to single acute administration of deltamethrin; a type II pyrethroid; as manifested by alterations in monoamines and some excitatory and inhibitory free amino acids in both brain and blood of adult male rats.
MATERIALS AND METHODS
Experimental Animals
One hundred twenty adult male albino rats (140-180 g) were assigned for the present study. Animals were obtained from local suppliers, housed in cages with natural temperature and illumination. They had free access to standard diet and tape water. Excitement and chance of fight between animals were minimized as possible as could be.
Chemicals
Deltamethrin (DM): (99.0% pure) was donated by Agrochemical Company-Egypt.
Selenium
Sodium selenite contains 45% as selenium was donated by Arab Company for Pharmaceutical and Medical Plant Mepaco-Egypt.
Antox
Antioxidant mixture was donated by Arab Company for Pharmaceutical and Medical Plant Mepaco-Egypt. It contains the three main antioxidant vitamins (A, C and E) together with selenium. Each tablet contains: Selenium (50 μg), Ascorbic acid (100 mg), Vitamin E (30 mg), Vitamin A acetate (5.54 mg) and medical yeast (105 μg).
Animal Groups and Experimental Design
After one week of acclimatization period, animals were randomly divided into four groups, 30 rats each.
Control Group C
Inoculated intraperitonially only once with 0.5 mL of corn oil and a daily volume of distilled water (1 mL/100 g body weight) given orally via a gastric tube over a total period of three weeks.
Deltamethrin Treated Group (DM)
Inoculated intraperitonially with a single dose of deltamethrin (¼ LD50) suspended in 0.5 mL of corn oil.
Selenium Treated Group (DMS)
Inoculated intrapertonially with a single dose of deltamethrin (¼ LD50) and an oral dose of sodium selenite (0.45 μg/100 g body weight) dissolved in distilled water given daily over a total period of three weeks.
Antox Treated Group (DMA)
Inoculated intraperitonially with a single dose of deltamethrin (¼ LD50) and an oral dose of Antox equivalent to 0.45 μg/100 g body weight of selenium dissolved in distilled water given daily over a total period of three weeks.
Blood and Brain Samples
Animals were sacrificed by decapitation 1, 2 and 3 weeks post inoculation. Serum, plasma and brain homogenates (1/10 wt/v in 70% ethanol) were prepared and kept frozen at -20 °C for subsequent analysis. Plasma samples and brain homogenates were used for estimation of the four monoamines: serotonine (5-HT), dopamine (DA), norepinephrine (NE) and epinephrine (E). Serum samples and brain homogenates were used for estimation of five free amino acids, namely glutamic acid, aspartic acid, taurine, GABA and glycine and the enzyme activity of acetylcholinesterse (AChE). The enzymatic activity of aspartate aminotransferase (AST) and alanine aminotransferase (ALT), as well as total protein and glucose concentrations were estimated in serum samples.
Methods
The LD50 of DM was determined according to Litchfield and Wilcoxon (1949) and was found to be 445 mg/100 g body weight after 24 h. Monoamines were analyzed by high performance liquid chromatography (HPLC) technique according to the method of Pagel et al. (2000). Amino acids were also detected by HPLC using the precolumn PTC derivatization technique according to Heinrikson and Meredith (1984). Estimation of acetylcholinesterase (AChE) activity in serum samples and brain homogenates was according to the method described by Gorun et al. (1978). Total serum protein was measured by the classical Biuret method according to Gornall et al. (1949) using the ready-made kits of Stanbio, Texas, USA. Glucose concentration was determined according to Trinder (1969) using the ready-made reagent kits of bioMérieux, France. Enzymatic activities of aspartate aminotransferase (AST) and alanine aminotransferase (ALT) were determined by the colorimetric method of Reitman and Frankel (1957) using the ready-made reagent kits of QCA-Spain.
Statistical Analysis
Data were analyzed by the one-way analysis of variance (ANOVA) followed by Duncan test according to Winter et al. (1991) using the SPSS-PC computer software package version 10.
RESULTS
[I] Brain and Blood Monoamines Serotonin (5-HT) Levels
As shown in Table 1, brain serotonin levels showed significant elevations in deltamethrin-intoxicated (DM) group versus the control (C) group during the whole experimental period (p < 0.05). DMS and DMA groups followed almost the same trend. Despite these elevations, brain serotonin levels showed a continuous reduction in all treated groups from the 1st to the 3rd weeks as compared to the control values. A significant rise was demonstrated in DMS group versus DM group at the 2nd and 3rd weeks of the experiment, while a significant reduction was demonstrated in DMA group versus DM group at the 2nd week. Comparing both DMS and DMA groups demonstrated significantly lower
| Table 1: | Effect of selenium or Antox supplementation on changes in brain and blood monoamine concentrations as well as acetylcholinesterase activity induced by acute deltamethrin intoxication |
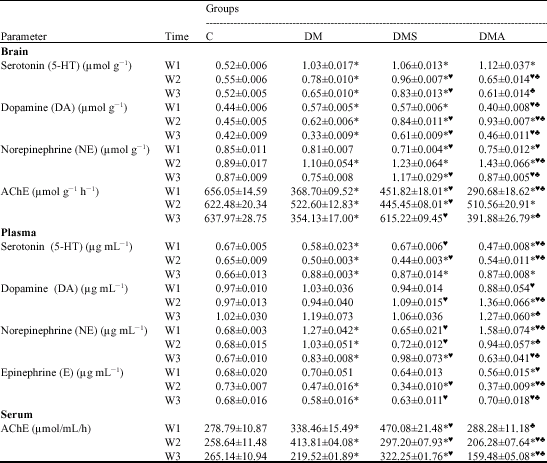 | |
| *= Significant versus C at p < 0.05;♥ = Significant versus DM at p < 0.05; ♠ = Significant versus DMS at p < 0.05 | |
brain serotonin levels in the latter versus the former group at the 2nd and 3rd weeks. DMA group showed almost normal brain 5-HT levels at the end of the experiment, indicating a good sign of therapeutic effect.
In opposition, plasma 5-HT levels revealed a general trend of reduction in all DM-treated groups during the 1st and 2nd week versus the control group except DMS group which exhibited an insignificant change versus C group at the 1st week (Table 1). In contrast, a significant rise was demonstrated in all DM-treated groups versus the control group at the end of the experiment. Significant differences have also been recorded in DMS and DMA groups versus DM group and among DMS and DMA groups at 1st and 2nd weeks. Signs of therapeutic effect could only be demonstrated in DMS group at the 1st week only.
Dopamine (DA) Levels
As compared to control rats, acute DM toxicity induced a significant increase in brain DA levels at 1st and 2nd week followed by a significant reduction at the 3rd week (Table 1). On the other hand, DMS group showed a significant rise versus the control group during the whole experimental period, while DMA group exhibited a significant rise versus the control group at the 2nd week only. At the 1st and 3rd weeks, there was a significant reduction in DMA group versus the DM and DMS groups. Signs of therapeutic effect could only be demonstrated in DMA group at the 1st and 3rd weeks of the experiment.
As regards plasma DA level, DM and DMS groups revealed insignificant changes versus the control group throughout the whole experimental period (Table 1). On the other hand, DMA group exhibited a significant rise versus C, DM and DMS groups at 2nd week and versus C and DMS at 3rd week. These findings exposed a negative impact of Se-supplementation in DMA group over serum DA level at the 2nd and 3rd weeks.
Norepinephrine (NE) Level
As shown in Table 1, DM-treated group exhibited a significant increase in brain NE levels as compared to control value at the 2nd week only. Levels were within the control values at early and last stages of the experiment. On the other hand, DMS group exhibited a significant reduction at the 1st week but a significant elevation at the 2nd and 3rd weeks as compared to the control group. DMA group exhibited the same trend seen in DMS group at the 1st and 2nd weeks but an insignificant change at the 3rd week versus the control values. Statistical analysis has also revealed significant variations between both DMS and DMA groups at the 2nd and 3rd weeks. These findings revealed a negative impact of Se-supplementation in either individual or combined form over brain NE levels throughout most of experimental period. The only exception was seen in DMA at the end of the experiment.
On the other side, plasma NE levels showed a significant elevation in DM group versus the control group throughout the whole experimental period. The values showed gradual reduction from the 1st to the 3rd week (Table 1). DMS group exhibited an insignificant change at the 1st and 2nd weeks but a significant rise at the 3rd week versus the control values. In opposition, DMA group exhibited a significant rise at the 1st and 2nd weeks but insignificant change at the 3rd week versus the control values. Significant variations were also detected in between DMS and DMA groups at different time intervals. These findings exposed a good sign of therapeutic effect of either individual or combined Se-supplementation over serum NE levels as affected by acute deltamethrin intoxication. In DMS group, the therapeutic effect appeared during the 1st and 2nd weeks, but in DMA group it appeared only at the end of the experiment.
Epinephrine (E) Level
Significant reduction in plasma epinephrine (E) levels was recorded in DM group versus the control group at the 2nd and 3rd weeks (Table 1). On the other hand, DMS group exhibited a significant reduction versus C group at the 2nd week only. Meanwhile, DMA group exposed a significant reduction versus C group at the 1st and 2nd weeks. At the end of the experiment, almost normal plasma epinephrine levels have been expressed in DMS and DMA groups indicating a good sign of therapeutic effect. Statistical analysis has also revealed significant variations between the three DM-treated groups at different time intervals.
Brain and Blood Acetylcholinesterase (AChE) Activity
As seen in Table 1, brain AChE activity revealed a general trend of significant reduction among all treated groups versus the control group throughout the whole experimental period. The only exception was seen in DMS group which exhibited almost normal value at the 3rd week. These findings indicated a good sign of therapeutic effect in DMS but not in DMA group over brain AChE activity at the end of the experiment.
As regards serum AChE activity, DM group exhibited a significant increase at the 1st and 2nd weeks followed by significant reduction at the 3rd week as compared to the control values (Table 1). Meanwhile, DMS group exhibited a significant rise in serum AChE activity versus C group that extended for the whole experimental period. In opposition, DMA group showed almost normal value at the 1st week but a significant reduction at the 2nd and 3rd weeks versus the
| Table 2: | Effect of selenium and Antox on changes in brain and blood free amino acids levels induced by acute deltamethrin intoxication |
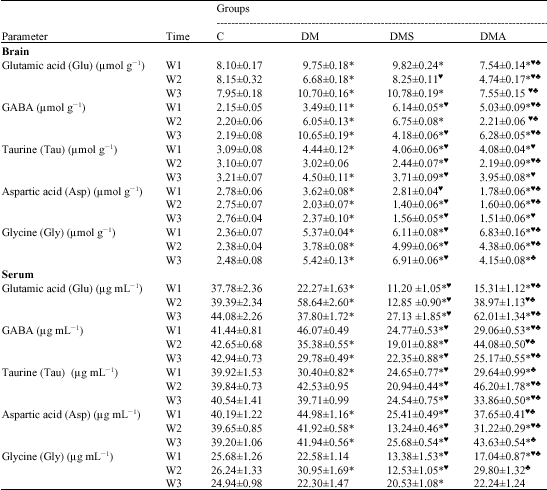 | |
| *= Significant versus C at p < 0.05;♥ = Significant versus DM at p < 0.05; ♠ = Significant versus DMS at p < 0.05 | |
control values. Among both Se-supplemented groups, DMA showed a significant reduction versus DMS through the whole experiment. These findings indicated a good therapeutic effect of Se-supplementation in combined but not in individual form over serum AChE activity at the early stage of the experiment only.
Brain and Blood Free Amino Acids
Glutamic Acid (Glu) Levels
As shown in Table 2, subacute DM intoxication induced a significant increase in brain Glu (excitatory amino acid) levels at the early and last stages of the experiment as compared to the control values. In contrast, a significant reduction was demonstrated in the same group at the 2nd week versus C group. The same trend of changes has been encountered in DMS group at the early and last stages of the experiment, while normal value has been expressed at the 2nd week. On the other hand, DMA group exhibited a significant reduction at the 1st and 2nd weeks of the experiment versus the control group. At the 3rd week almost normal value has been expressed in DMA. These findings indicated a good sign of therapeutic effect of Se-supplementation over brain Glu levels at the middle and last stages of the experiment in DMS and DMA groups, respectively.
As regards serum Glu levels, data shown in Table 2 revealed an inverted image to that demonstrated in brain in DM group. Meanwhile, a significant reduction was demonstrated in DMS group throughout the whole experiment versus both C and DM groups. On the other side, DMA group exhibited a significant reduction at the 1st week, but a significant elevation at the 3rd week as compared to control values. Statistical analysis has also revealed significant variations among the three treated groups (DM, DMS, DMA) at different time intervals. These findings indicated a good sign of therapeutic effect of Se-supplementation in the combined but not in the individual form over serum Glu levels at the middle stage of the experiment only.
GABA Levels
As demonstrated in Table 2, brain GABA (inhibitory amino acid) levels showed a marked and significant rise in all DM-treated groups throughout most of the experimental period as compared to the respective control values. The only exception was demonstrated in DMA group at the 2nd week, where almost normal value was expressed. In DM group, the GABA levels were increasing by time.
In opposition, serum GABA levels exhibited an inverted image to that demonstrated in the brain in all DM-treated groups for most of the experimental period in comparison with the control values. The only exception was seen in DM group at the 1st week, where an insignificant change versus the control group was demonstrated.
Among the two Se-supplemented groups, DMA group showed the minimal change in both brain and serum levels of GABA at the middle stage of the experiment with a good sign of therapeutic effect.
Taurine (Tau) Levels
As shown in Table 2, significant elevations were demonstrated in brain Tau (inhibitory amino acid) levels among all treated groups at the early and last stages of the experiment as compared to the control values. In opposition, a significant reduction has been detected in DMS and DMA groups at the 2nd week versus both C and DM groups. Both Se-supplemented groups seemed to slightly attenuate the stimulatory effect of acute deltamethrin intoxication on brain Tau levels at the 3rd week of the experiment. However, neither of them showed signs of complete recovery.
With respect to serum taurine levels, DM group showed a significant reduction versus the control group at the 1st week only. Meanwhile, DMS group exhibited a significant reduction versus both C and DM groups throughout the whole experimental period. On the other hand, DMA group revealed a significant reduction at early and last stages but a significant rise at the middle stage of the experiment versus the respective control values. At the same time, values expressed in DMA were significantly higher than those demonstrated in DMS group. No signs of therapeutic effect could be demonstrated in both DMS and DMA groups over serum Tau levels throughout the whole experimental period.
Aspartic Acid (Asp) Levels
Data presented in Table 2 revealed a significant reduction in brain Asp (excitatory amino acid) levels among all treated groups versus the control group at the 2nd and 3rd weeks. These reductions were more obvious in Se-supplemented groups. At the 1st week, a significant increase in DM group, but a significant decrease in DMA group has been demonstrated versus the control value.
As regards serum Asp levels, a significant rise was demonstrated in DM group versus the control group throughout the whole experimental period (Table 2). In contrast, DMS group exhibited a marked and significant reduction versus C and DM groups throughout the whole experimental period. In DMA group, fluctuations in serum Asp levels, with significant reduction at the 2nd week and significant elevation at 3rd week, could be detected in comparison with the control group. Statistical analysis has also revealed a significant rise in DMA versus DMS through the whole experiment. Signs of therapeutic effects were more represented in DMS group in case of the brain, but more represented in DMA group in case of the serum at the early stage of the experiment week only.
| Table 3: | Effect of selenium and Antox supplementation on changes in serum biochemical parameters induced by acute deltamethrin intoxication |
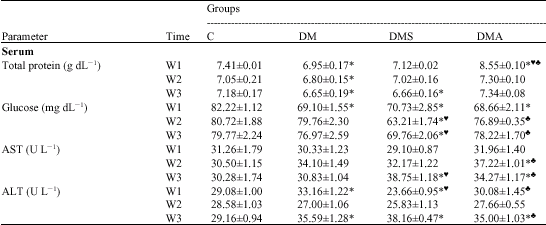 | |
| * = Significant versus C at p < 0.05;♥ = Significant versus DM at p < 0.05; ♠ = Significant versus DMS at p < 0.05 | |
Glycine (Gly) Levels
Data presented in Table 2 revealed a marked and significant rise in brain Gly (inhibitory amino acid) levels in all DM-treated groups versus the control group through the whole experimental period. Se-supplementation in both individual and combined forms seemed to exaggerate the effect of DM intoxication and failed to express any therapeutic effect on brain Gly level. Despite this tendency, DMA group showed the minimal changes, especially at the end of the experiment.
On the other hand, DM intoxication induced a significant elevation in serum Gly level as compared to the control value at the 2nd week only. In opposition, DMS group exhibited a significant reduction versus the control values at all experimental periods, while DMA group expressed a significant reduction at the 1st week only. DMA group showed a good sign of therapeutic effect on serum Gly levels for the rest of the experiment.
Biochemical Parameters
Serum Total Protein
After a single acute administration of DM, a significant reduction in total serum protein was recorded throughout the whole experimental period in comparison with the control group (Table 3). DMS group exhibited a considerable improvement in serum total protein levels at the early and middle stages of the experiment. On the other hand, DMA group exhibited a significant rise versus the control value at the 1st week only. For the rest of the experiment, normal values have been expressed in DMA group with good sign of therapeutic effect on total serum protein.
Among the two Se-supplemented groups, DMS exhibited the best therapeutic effect at the early and middle stages of the experiment, whereas DMA group expressed the best effect at the middle and last stages of the experiment.
Serum Glucose
Data presented in Table 3 revealed a significant reduction in serum glucose level in DM and DMA groups versus control group at the early stage of the experiment only. In DMS group, a significant reduction was demonstrated versus the control values through the whole experiment.
Among the two Se-supplemented groups, DMA exhibited the minimal changes as compared to control values. However, no sign of therapeutic effect could be demonstrated in DMA group versus DM group at the early stage of the experiment.
Serum AST Activity
As compared to the control group, an insignificant change in serum AST activity was demonstrated in DM group throughout the whole experimental period (Table 3). Meanwhile, a significant elevation was recorded in DMS group versus both C and DM groups at the 3rd week only and in DMA group versus both C and DM groups at the 2nd and 3rd weeks. Both Se-supplemented groups induced a significant rise in AST activity, but DMS showed the minor changes as compared to control values.
Serum ALT Activity
Data presented in Table 3 revealed a significant rise in serum ALT activity in DM group versus the control group at the early and last stages of the experimental period. Meanwhile, DMS group showed some fluctuations, with significant reduction at the early stage, but a significant rise at the last stage of the experiment versus the control group. DMA group, however, exhibited a significant rise at the end of the experiment only. Selenium supplementation, either in individual or combined forms seemed to counteract the delayed effect of acute DM intoxication on serum ALT activity at the early stage of the experiment only, but DMA showed the best therapeutic effect.
DISCUSSION
The present study was interesting to expose the therapeutic effect of selenium supplementation, either individually or in antioxidant combination over delayed neurotoxic effects of a single acute dose of deltamethrin (DM), a type II pyrethroid, on brain and blood monoamines and free amino acids.
The present data explored significant elevations in brain levels of the three neurotransmitters, 5-HT, DA and NE among all DM-treated groups during most of the experimental period with few exceptions. These elevations may be attributed to one or more of the following mechanisms: the stimulatory effect of deltamethrin on the activity of the protein kinase C associated with an increase in the intracellular concentration of inositol triphosphate (IP3) and free calcium, as well as an increase in overall and specific protein phosphorylation within the synapses which finally stimulates the transmitter release as cited by Enan and Matsumura (1993); the role of deltamethrin as calcineurin inhibitor that inhibits Ca2+ ionophore-induced dephosphorylation as reported by Carrera et al. (1996); the potent and stereoselective action of deltamethrin on the voltage-sensitive sodium channel that increases influx into synaptic terminals producing membrane depolarization and neurotransmitter release as recorded by Eells et al. (1992) and pyrethroid-induced inhibition in monoamine oxidase in different brain regions that leads to significant neurochemical changes which may be culminated in perturbed synaptic function as supported by the work of Rao and Rao (1993) and Husain et al. (1994, 1996) who reported an inhibition of monoamine oxidase in rat brains after pyrethroid exposure.
One more additional mechanism for the general enhancement of brain monoamine neurotransmitters may be related to the rise in brain GABA levels seen in the present study in all DM-treated groups throughout the whole experimental period. GABA can act persynaptically via a GABAB-like receptor to increase the release of neurotransmitters as supported by the previous report of Gutovitz et al. (2001).
The present data have also exposed a significant reduction in brain level of DA in DM group at the 3rd week and in DMA group at the 1st week which might be attributed to inhibited DA synthesis as supported by the recent work of Liu et al. (2006). According to the authors, the contents of DA metabolites DOPAC and HVA were increased in striatum after deltamethrin administration, indicating a stimulated dopamine turnover and inhibited DA synthesis. The authors claimed a novel aspect of deltamethrin neurotoxicity and suggested tyrosine hydroxylase as a molecular target of deltamethin on dopamine metabolism in the nigrostriatal pathway. The findings of Bloomquist et al. (2002) that oxidative damage induced by pyrethroid insecticides, especially deltamethrin, possess specific effects on striatal dopaminergic neurotransmission particularly the substantia nigra might support this idea.
From the other side, the present study has demonstrated no signs of therapeutic effect of individual selenium supplementation over all measured brain monoamines. These findings might be explained by the possible selenium-deltamethrin interaction for the extended period of the experiment which seemed to exaggerate the delayed neurotoxic effects of deltamethrin on brain monoamines. This postulation can be supported by comparing the brain monoamine levels in both DMS and DM groups throughout most of the experimental period, where significantly higher values were expressed in the former versus the later group. Nevertheless, it should be noticed that in case of brain 5-HT and DA, the levels were almost decreasing by time. It seemed that excessive production of free radicals, likely induced by acute deltamethrin in brain tissues, needs further time of compensation by selenium supplementation. This might also be a dose-dependent effect of individual selenium supplementation. Perhaps higher concentrations of selenium were necessary to achieve a therapeutic effect.
In contrast, the present data have exposed a good sign of improvement in brain levels of 5-HT and NE at the 3rd week by Se-supplementation in the combined form (Antox). The different levels of biotransformation of selenite in Antox drug inside the central nervous system might account for this improvement. It might also be the combination of Se with other antioxidants in Antox that causes these good therapeutic effects. This interpretation can be supported by the fact that vitamins E and C (the main constituents of Antox) are the main antioxidants in the brain (Siegel et al., 1999). Vitamin E supplementation has shown to potentiate the antioxidant system and to lower pyrethroid-induced oxidative stress in rat tissues as cited by Kale et al. (1999b). Moreover, Kumar et al. (2001) reported that animals appear normal within 2 weeks, after receiving vitamin E and selenium when they beard paralysis as a side effect of deltamethrin treatment.
On the other hand, vitamin C has shown to exert a powerful protection against degenerative changes in the brain associated with oxidative damage (Ghosh et al., 1996). Vitamin C is also known to act as free radical trap and as cofactor in the synthesis of biologically antioxidant materials such as glutathione (Tppel et al., 1968; Khedr et al., 1999). Dietary supplement of ascorbic acid at appropriate level appeared to be a good way to counteract deltamethrin toxicity in catfish as reported by Datta and Kaviraja (2003). In addition, Daoud et al. (2000) has shown that Antox inhibits free radical generation, decreases the inflammatory reaction and increases chance of the drug reaching its target during treatment of trichinosis.
With respect to selenium, it is known to stimulate Na, K-ATPase activity and inhibit lipid peroxidation. Since Na, K-ATPase activity is known to be inhibited by oxygen free radicals, likely formed by deltamethrin, selenium supplementation appears to exert its beneficial effect on the Na, K-ATPase activity by preventing free radical-induced damage (Siegel et al.,1999).
The present study has also revealed a significant reduction in plasma 5-HT levels at the early and middle stage but a significant elevation at the last stage of the experiment in all DM-treated groups. At the same time, there was a gradual reduction in brain levels of 5-HT seen in the three DM-treated groups throughout the whole experimental period. These time-dependent changes in plasma 5-HT in DM-treated groups seems to be correlated with changes in plasma membrane reuptake of this neurotransmitter and/or the rate of degradation of DM by hepatic cells.
The present investigation has also explored a significant elevation in plasma norepinehrine (NE) levels in DM-intoxicated group during the whole experimental period. In contrast, plasma epinephrine (E) levels have shown a significant reduction during the middle and last stages of the experiment. The present elevation in plasma NE during the whole experimental period might be due to the stimulatory effect of deltamethrin on adrenal medullary chromaffin cells that spontaneously secrete catecholamines as reported by Bickmeyer et al. (1994). Several lines of evidence have also indicated that type II pyrethroids are strong inducers for catecholamine release (Ray and Cremer, 1979; Cremer and Seville, 1982; Bradburry et al., 1983; Ray, 1991). The present findings run partly in parallel with those reported by Santoni et al. (1999) who showed marked and long lasting increase in both epinephrine and norepinephrine plasma concentrations after exposure to cypermethrin. Deltamethrin can also elicit vigorous autonomic and neuro-endocrine responses due to an indirect action on the hypothalamic-pituitary axis leading to increase in plasma concentrations of corticosteriods and catecholamines as cited by De Boer et al. (1988) and Queiroz (1993). Moreover, type II pyrethroids have shown to enhance mesenteric blood vessel and cardiac contractility via circulating and locally released catecholamines (Forshaw and Bradbury, 1983; Berlin et al., 1984).
On the other hand, the present reduction in plasma E levels shown in DM-treated groups seems to be correlated with a selective inhibitory effect of DM on phenylethanolamine-N-methyltarsnferase (PNMT) which catalyzes the conversion of norepinephrine to epinephrine However, this contradictory effect of DM on plasma NE and E needs further investigation.
The present data have also exposed no signs of therapeutic effect on plasma 5-HT levels but some good signs of therapeutic effect on both plasma NE and E levels due to Se-supplementation at different time intervals. In case of plasma NE and E levels, DMS showed the best therapeutic effect, which indicates a specific and time-dependent therapeutic effect of individual Se supplementation at the dose used on plasma levels of these catecholamines. This effect seems be correlated with the antioxidant properties of selenium.
In the present research data collected on brain AChE activity proved that DM is an effective anti-cholinesterase agent since the enzyme activity was inhibited in brain tissue in all DM-treated groups throughout most of the experimental period. This finding runs in parallel with the previous studies on pyrethroid insecticides (Aldridge et al., 1978; Rawi, 1984; Mohamed and El-Sheamy, 1988; Eells et al., 1992). Several investigators have attributed this inhibition to attachment of pyrethroids to the ends of cholinergic nerve fibers that inhibit release of acetylcholineterase (Yamamoto, 1970; Narahashi, 1971; Mohamed, 1988; Mohamed and El-Sheamy, 1988; Eells et al., 1992). This effect might also be due to inhibited AChE synthesis and/or increased degradation in brain tissues by acute deltamethrin intoxication. The present data have also shown that DMS group only was successful to induce some good signs of therapeutic effect on brain AChE levels at the end of the experiment. This effect might be explained by delayed antagonistic effect of individual, but not combined selenium supplementation over synthetic pathways and/or rate of degradation of AChE in brain tissues as affected by DM intoxication.
On the other hand, the present study recorded a significant increase in serum AChE at the early and middle stages and a significant reduction at the last stage of the experiment in the DM group. In this context, Shaker et al. (1988) claimed that chronic toxicity by dimethoate and deltamethrin in rabbits has shown to induce primary elevation in plasma cholinesterase activity followed by 40% inhibition in comparison with the control animals. Present results also run in agreement with the findings of Novogorodskaya et al. (1971), Miyata et al. (1973) and Hussein et al. (1991) using other pesticides.
The present finding that brain AChE levels were reduced while serum levels were elevated in DM group seems to be correlated with brain tissue damage induced by free radical likely formed by acute deltamethrin intoxication, which might end with increased release of the enzyme in serum. Brain tissue damage due to acute DM intoxication has been documented in our laboratory by the work of Ismail (2005).
The present study has also shown that individual and combined selenium supplementation exhibited opposite effects on plasma cholinesterase activity under our experimental conditions. In the former case, selenium supplementation appeared to be synergistic, but in the latter case Antox supplementation seemed to be antagonistic to DM, reflecting different mechanism of action. In the latter case, however, some good signs of therapeutic effect have been demonstrated at the early stages of the experiment.
The present data have also exposed a significant elevation in brain levels of Glu and GABA in DM group, which run in parallel with the recent studies of Symington et al. (2007) who claimed that DM stimulates Ca2+-dependent release of L-glutamate from isolated brain synaptosomes. Nicholson et al. (1987) also claimed that deltamethrin stimulates the spontaneous release of [3H] GABA from mammalian brain synaptosomes superfused in vivo with non-depolarizing saline buffer. According to Wang et al. (2000), disturbance of glutamate and GABA metabolism showed to play an important role in the development of pyrethroid neurotoxicity.
This elevation in brain levels of Glu and GABA can be explained by one or more of the following mechanisms: neural damage induced by DM as previously reported by Groose et al. (2002); neurological signs and symptoms that characterize moderate hypoglycemia seen in the present study in DM group (Table 3) and as confirmed by previous studies using other insecticides (Srivastava and Singh, 1981; Enan et al., 1982; Agrawal et al., 1987; Rajini and Krishnakumari, 1988; Bendahou et al., 1999); the utilization of amino acids like glutamate and glutamine as alternative energy sources in moderate hypoglycemia as supported by preliminary reports of Butterworth (1983) and Butcher et al. (1987), who studied the adverse effect of hypoglycemia on the synthesis of the amino acid neurotransmitters, GABA and glutamate. They revealed that hypoglycemia produces a substantial increase in extracellular concentrations of glutamate and GABA. It has also been suggested that pyrethroid, especially deltamethrin, can disturb the function of brain tissue in high-affinity-glutamate uptake with their alpha-cyano-group which could play an important role in cerebral stimulation in mammals (Zhao et al., 1997). Recently, Wu and Liu (2003) reported that, deltamethrin induces neurodegeneration via a glutamate dependent pathway.
On the other hand, the signs of improvement in brain levels of both Glu and GABA seen in DMS and DMA groups at different time intervals of the experiment seem to be correlated with the antioxidant effect of selenium either in individual or combined supplementation. The combined supplementation appeared to be more effective in counteracting the effects of DM. However, the contradictory effects of DMS and DMA groups over brain levels of GABA reflect different mechanisms of action which might include the rate of uptake by brain tissues and/or rate of metabolism of this amino acid in the brain.
The present data have also revealed a general trend of reduction in serum levels of Glu and GABA in all DM-treated groups during most of the experimental period, which was almost an inverted image to that described in the brain. This reduction in both serum levels of Glu and GABA seems to be correlated with the stimulated uptake of these two amino acids by the brain tissues under acute DM intoxication as described above. Signs of therapeutic effects could only be demonstrated in DMA group at the middle of the experiment which might be attributed to the combined antioxidant effect of Antox. However, the contradictory effect seen in both DMS and DMA groups on serum levels of Glu might be attributed to time-dependent changes in the rate of uptake by the brain and/or the rate of metabolism in the blood for this amino acid under the experimental conditions.
The present investigation has also shown significant alterations in both brain and serum Tau levels in all DM-treated groups at different time intervals of the experiment. The effect was almost stimulatory in case of the brain but almost inhibitory in case of serum. In general, taurine is an inhibitory neurotransmitter that initiates membrane polarization, which results in decreased neuronal firing (Oja and Kornto, 1983). Taurine is also known to decrease K+ stimulated release of epinephrine and acetylcholine, which in turn support the present finding. According to Trachtman et al. (1988) and Siegel et al. (1999) taurine is an important cerebral osmoprotective molecule, where it constitutes 50% of the adaptive intracellular osmol pool.
Rise of taurine levels in the brain seems to be correlated with brain damage induced by DM. It might also be correlated with stimulatory effect of DM on the synthesis and/or release of this amino acid neurotransmitter. On the other hand, lowered serum values seemed to be correlated with inhibited synthesis in sites other than the brain. It is worth to mention here that this reduction was only temporary in DM group but was permanent in DMS indicating that Se-supplementation was synergistic to DM. The fact that no signs of modulation could be demonstrated in both brain and serum levels by individual or combined Se-supplementation under our experimental conditions reflect the highly specific adverse effects of DM on brain and serum taurine levels.
The present data has also indicated an inhibitory effect on brain Asp levels in all DM-treated groups during most of the experimental period. On the other hand, serum levels have exposed an opposite stimulatory effect in DM group but an inhibitory effect in DMS group. Individual or combined Se-supplementation seemed to be synergistic with DM as regards brain aspartic acid levels, but antagonistic as regards serum levels reflecting different mechanism of action in different tissues. In both cases it showed no signs of therapeutic effect under our experimental conditions. The reducing effect of DM on brain Asp levels together with the increasing effect on serum levels seemed to be correlated with inhibited rate of synthesis by the brain tissue and/or stimulated release of this neurotransmitter into serum. The increased serum levels might also be correlated with initiated synthesis by tissues other than the brain.
The present data also explored remarkable increase in brain levels as well as alteration in serum levels of Gly in all DM-treated groups. Glycine is an inhibitory glucogenic nonessential amino acid that can be synthesized from therionine and serine by a reversible reaction catalyzed by the enzyme serine-transhydroxy methylase. Serine itself can be formed in the nervous tissue from glucose via the intermediates 3-phosphoglycerate and 3-phosphoserine (Ahmed et al., 1992). The high brain levels of Gly seem to be correlated with stimulated synthetic pathway and/or release of this neurotransmitter amino acid by acute administration of DM. Individual and combined Se-supplementation showed a synergistic effect in case of the brain, but an antagonistic action in case of serum in DMA group only reflecting different mechanism of action in different tissues. The sings of improvement seen in case of serum in DMA group at the middle and end of the experiment might be associated with the scavenging effect of the antioxidants mixture in Antox preparation against free radicals likely formed by DM in different tissues.
As regards reduced total serum protein content encountered in DM group during the whole experiment, it might be attributed to one or more of the following reasons: disturbance of amino acid levels as supported by Felig et al. (1970), renal dysfunction as supported by Bishop et al. (1992), blood loss in open wounds or internal bleeding as mentioned by Bishop et al. (1992) or impaired protein synthesis in liver, which in turn affect the protein balance in serum. Both individual and combined Se-supplementation seem to compensate for this reduction during most of the experimental period, reflecting the potency of these antioxidants in scavenging free radicals likely formed by DM in different tissues.
The significant decrease in serum glucose level encountered in all treated groups at early stages of the experiment might be coupled with the signs of reduced food intake and diarrhea seen in the three deltamethrin-treated groups. Early signs of DM acute toxicity as starvation, distended stomach and intestine together with malabsorption might also end with a type of hypoglycemia. Several reports have also reported hypoglycemia in animals treated with other pesticides (Srivastava and Singh, 1981; Enan et al., 1982; Agrawal et al., 1987; Rajini and Krishnakumari, 1988; Bendahou et al., 1999).
On the other hand, the significant increases in the activity of serum ALT and AST encountered in all DM-treated groups at different time intervals of the experiment seem to be associated with hepatic cellular damage. However, serum ALT levels showed minor changes in DM but moderate changes in both DMS and DMA groups. These findings run in agreement with those reported by El- Banhawy et al. (1984). Signs of therapeutic effect by individual or combined Se-supplementation could not be seen under our experimental conditions except in DMA at early stages of the experiment, reflecting different mechanisms of action.
The present study revealed the adverse neurotoxic effects of acute DM toxicity in different treated groups which might be a primary response to deltamethrin exposure per se or secondary to free radicals likely induced by this pyrethroid. Previous studies have accumulated data suggesting that pyrethroids produce oxidative stress through the generation of free oxygen radicals and that antioxidant defense systems are disturbed by pyrehroids, especially deltamethrin (Kale et al., 1999a, b; Sayeed et al., 2003; Parvez and Raisuddin, 2006; Rehman et al., 2006; Yousef et al., 2006; Li et al., 2007).
The present study also indicated that Se-supplementation either individually or in antioxidant preparation has a considerable ability to compensate for some but not all delayed effects of acute deltamethrin intoxication under the experimental conditions. These signs of therapeutic effects might be correlated with the scavenging effect of selenium either individually or in presence of other antioxidants like vitamin C and vitamin E (present in Antox) and/or the enhancing ability for the antioxidant defense system. These postulations are supported by the previous reports of Nakagawa et al. (1991), Ji et al. (1995), Stevenson et al. (1995), El-Manakhly (1996), Szarek et al. (1997), Blasiak and Kowalik (1999) and El-Zarkouny et al. (1999) who proved the protective role of selenium, vitamin E and vitamin C either individually or in mixture against oxidative stress effects induced by different pesticides including pyrethroids. On the other hand, selenium has demonstrated a great potential in the biotransformation and detoxification of many toxic materials (Shoka et al., 1994).
It is worth to mention here that among the two Se-supplemented groups, it was difficult to see a clear therapeutic effect on all measured parameters. DMS showed the best results in some cases but DMA showed the best results in others reflecting different mechanisms of action. It appeared that these therapeutic effects are time-dependent.
REFERENCES
- Agrawal, D.A., R.N. Khanna, M. Anand, G.S. Gupta and P.K. Ray, 1987. Lindane-induced changes in glucose and glutathione levels in cats. Toxicol. Lett., 38: 77-82.
CrossRefPubMedDirect Link - Aldana, L., V. Tsutsumi, A. Craigmill, M.I. Silveira and E.G. de Mejia, 2001. Alpha-tocopherol modulates liver toxicity of the pyrethroid cypermethrin. Toxicol. Lett., 125: 107-116.
PubMedDirect Link - Berlin, J.R., T. Akera, T.M. Brody and F. Matsumura, 1984. The inotropic effects of a synthetic pyrethroid decamethrin on isolated guinea-pig atrial muscle. Eur. J. Pharmacol., 98: 313-322.
PubMedDirect Link - Bickmeyer, U., F. Weinsberg and H. Wiegand, 1994. Effects of deltamethrin on catecholamine secretion of bovine chromaffin cells. Arch. Toxicol., 68: 532-534.
CrossRefDirect Link - Blasiak, J. and J. Kowalik, 1999. Effect of paraoxon methyl and parathion methyl on DNA in human lymphocytes and protective action of vitamin C. Pest. Sci., 55: 1182-1186.
CrossRefDirect Link - Bloomquist, J.R., R.L. Barlow, J.S. Gillette, W. Li and M.L. Kirby, 2002. Selective effects of insecticides on nigrostriatal dopaminergic nerve pathways. Neurotoxicology, 23: 537-544.
CrossRefPubMedDirect Link - Bradburry, J.E., P.J. Forshaw, A.J. Gray and D.E. Ray, 1983. The action of mephenesin and other agents on the effect produced by two neurotoxic pyrethroids in the intact and spinal rat. Neuropharmacology, 22: 907-914.
CrossRef - Butcher, S.P., M. Sandberg, H. Hagberg and A. Hamberger, 1987. Cellular origin of endogenous amino acids released into the extracellular fluid of the rat striatum during severe insulin-induced hypoglycemia. J. Neurochem., 48: 722-728.
CrossRefDirect Link - Carrera, A., J. Moos, X.P. Ning, G.L. Gerton, J. Tesarik, G.S. Kopf and S.B. Moos, 1996. Regulation of protein tyrosine phosphorylation in human sperm by a calcium/calmodulin-dependent mechanism: Identification of a kinase anchor proteins as major substrates for tyrosine phosphorylation. Dev. Voil., 180: 284-296.
CrossRefDirect Link - Chugh, Y., A. Sankaranarayanan and P.L. Sharma, 1992. MK-801 antagonizes the lethal action of centrally and peripherally administered cypermethrin in mice and rats. J. Pharm. Pharmacol., 44: 521-523.
PubMedDirect Link - Cremer, E.J. and P.M. Seville, 1982. Comparative effects of two pyrethroids, deltamethrin and cismethrin on plasma catecholamines and on blood glucose and lactate. Toxicol. Applied Pharmacol., 66: 124-133.
CrossRef - Daoud, A.A., A.E. Abdel Ghaffar, F.A. Deyab and T.M. Essa, 2000. The effect of antioxidant preparation (Antox) on the course and efficacy of treatment of trichinosis. J. Egypt. Soc. Parasitol., 30: 305-315.
PubMedDirect Link - Datta, M. and A. Kaviraj, 2003. Ascorbic acid supplementation of diet for reduction of deltamethrin induced stress in fresh water catfish Clarias gariepinus. Chemosphere, 53: 883-888.
CrossRef - De Boer, S.F., J. van der Gugten, J.L. Slangen and T.H. Hijzen, 1988. Changes in plasma corticosterone and catecholamine content induced by low doses of deltamethrin in rats. Toxicology, 49: 363-370.
CrossRef - Eells, J.T., P.A. Bandettini, P.A. Holman and J.M. Propp, 1992. Pyrethroid insecticide-induced alteration in mammalian synaptic membrane potential. J. Pharmacol. Exp. Ther., 262: 1173-1181.
Direct Link - El-Zarkouny, S.A., M.A. Ayoub, M.H.G. Ishak, F.D. El Nouty, G.A. Hassan, Z.R. El-Ezz and M.H. Salem, 1999. Effect of carbosulfan pesticide and selenium on some semen characteristics and serum testosterone in male rabbits. Int. J. Environ. Health Res., 9: 117-124.
CrossRefDirect Link - El-Zayat, E., M. El-Halwagy and M. Farid, 2005. Physiological and biochemical alterations induced by subacute administration of deltamethrin can be possibly corrected by zinc supplementation in male albino rat. J. Biological Sci., 5: 211-221.
CrossRefDirect Link - Enan, E., A.H. El-Sebae, O.H. Enan and S. El-Fiki, 1982. In vivo interaction of some organophosphorus insecticides with different biochemical targets in white rats. J. Environ. Sci. Health B., 17: 549-570.
CrossRefPubMedDirect Link - Enan, E. and F. Matsumura, 1993. Activation of phosphoinositide/protein kinase C pathway in rat brain tissue by pyrethroids. Biochem. Pharmacol., 45: 703-710.
CrossRef - Felig, P., E. Marliss, J.L. Ohman and C.F. Cahill Jr., 1970. Plasma amino acid levels in diabetic ketoacidosis. Diabetes, 19: 727-728.
PubMedDirect Link - Forshaw, P.J. and J.E. Bradbury, 1983. Pharmacological effects of pyrethroids on the cardiovascular system of the rat. Eur. J. Pharmacol., 91: 207-213.
CrossRef - Ghosh, M.K., D.J. Chattopadhyay and I.B. Chatterjee, 1996. Vitamin C prevents oxidative damage. Free Radic. Res., 25: 173-179.
Direct Link - Gilgun-Sherki, Y., Z. Rosenbaum, E. Melamed and D. Offen, 2002. Antioxidant therapy in acute central nervous system injury: Current state. Pharmacol. Rev., 54: 271-284.
CrossRefDirect Link - Gornall, A.G., C.J. Bardawill and M.M. David, 1949. Determination of serum proteins by means of the biuret reaction. J. Biol. Chem., 177: 751-766.
CrossRefPubMedDirect Link - Gorun, V., I. Proinov, V. Baltescu, G. Balaban and O. Barzu, 1978. Modified Ellman procedure for assay of cholinesterase in crude enzymatic preparation. Anal. Biochem., 86: 324-326.
PubMedDirect Link - Groose, G., T. Thiele, E. Heuckendorf, E. Schopp, S. Merder, G. Pickert and G. Ahnert-Hilger, 2002. Deltamethrin differentially affects neurnal subtyes in hippocampal primary culture. Neuroscience, 112: 233-241.
PubMedDirect Link - Gutovitz, S., J.T. Birmingham, J.A. Luther, D.J. Simon and E. Marder, 2001. GABA enhances transmission at an excitatory glutamatergic synapse. J. Neurosci., 21: 5935-5943.
PubMedDirect Link - He, F., 2000. Neurotoxic effects of insecticides-current and future research: A review. Neurotoxicology, 21: 829-835.
PubMedDirect Link - Husain, R., M. Malaviya, P.K. Seth and R. Husain, 1994. Effect of deltamethrin on regional brain polyamines and behaviour in young rats. Pharmacol. Toxicol., 74: 211-215.
CrossRefDirect Link - Husain, R., R. Husain, A.M. Adhami and P.K. Seth, 1996. Behavioral, neurochemical and neuromorphological effects of deltamethrin in adult rats. J. Toxicol. Environ. Health, 48: 515-516.
CrossRefDirect Link - Kale, M., N. Rathore, S. Jhon and D. Bhatnagar, 1999. Lipid peroxidation and antioxidant enzymes in rat tissue in pyrethroid toxicity: Possible involvement of reactive oxygen species. J. Nutr. Environ. Med., 9: 37-46.
Direct Link - Kale, M., N. Rathore, S. Jhon, D. Bhatnagar, S.S. Nayyar and V. Kothari, 1999. The protective effect of vitamin E in pyrethroid-induced oxidative stress in rat tissue. J. Nutr. Environ. Med., 9: 281-287.
Direct Link - Karen, D.J., W. Li, P.R. Harp, J.S. Gillette and J.R. Bloomquist, 2001. Striatal dopaminergic pathways as a target for the insecticide permethrin and chloropyrifos. Neurotoxicology, 22: 811-817.
CrossRef - Li, H.Y., Y.F. Zhong, S.Y. Wu and N. Shi, 2007. NF-E2 related factor 2 activation and heme oxygenase-1 induction by tert-butylhydroquinone protect against deltamethrin-mediated oxidative stress in PC12 cells. Chem. Res. Toxicol., 29: 1242-1251.
CrossRefPubMedDirect Link - Litchfield, Jr. J.T. and F. Wilcoxon, 1949. A simplified method of evaluating dose-effect experiments. J. Pharmacol. Exp. Ther., 96: 99-113.
PubMedDirect Link - Liu, G.P., Q. Ma and N. Shi, 2006. Tyrosine hydroxylase as a target for deltamethrin in the nigrostriatal dopaminergic pathway. Biomed. Environ. Sci., 19: 27-34.
PubMed - Mohamed, Z.A. and M.K. El-Sheamy, 1988. Aminotransferases, alkaline phosphatase and cholinesterase of hen tissues administered a single oral dose of alpha-cyano pyrethoids. Egypt. J. Food Sci., 16: 105-109.
Direct Link - Nakagawa, Y., I.A. Cotgreave and P. Moldeus, 1991. Relationships between ascorbic acid and alpha tocopherol during diquat-induced redox cycling in isolated rat hepatocytes. Biochem. Pharmacol., 42: 883-888.
PubMedDirect Link - Narahashi, T., 1971. Mode of action of pyrethroids. Bull. World Health Organ., 44: 337-345.
Direct Link - Pagel, P., J. Blome and H.U. Wolf, 2000. High performance liquid chromatographic separation and measurement of various biogenic compounds possibly involved in the pathomechanism of Parkinson's disease. J. Chromatogr. B., 746: 297-304.
CrossRef - Parvez, S. and S. Raisuddin, 2006. Copper modulates non-enzymatic antioxidants in the freshwater fish Channa punctata (Bloch) exposed to deltamethrin. Chemosphere, 62: 1324-1332.
CrossRef - Queiroz, M.L., 1993. Hematopoietic effects in mice exposed to deltamethrin and hydrocortisone. Int. J. Iummunopharmacol., 15: 301-307.
CrossRef - Rajini, P.S. and M.K. Krishnakumari, 1988. Toxicity of pirimiphos-methyl: II. Effect of dietary feeding on blood and urine constituents in albino rats. J. Environ. Sci. Health B, 23: 145-158.
CrossRefPubMedDirect Link - Rao, G.V. and K.S.J. Rao, 1993. Inhibition of monoamine oxidase-a of rat brain by pyrethroid-an in vitro kinetic study. Mol. Cell Biochem., 124: 107-114.
CrossRefPubMedDirect Link - Ray, D.E. and J.E. Cremer, 1979. The action of decamothrin (a synthetic pyrethroid) on the rat. Pest. Biochem. Physiol., 10: 333-340.
CrossRef - Rehman, H., M. Ali, F. Atif, M. Kaur, K. Bhatia and S. Raisuddin, 2006. The modulatory effect of deltamethrin on antioxidants in mice. Clin. Chim. Acta, 369: 61-65.
CrossRef - Reitman, S. and S. Frankel, 1957. A colorimetric method for the determination of serum glutamic oxalacetic and glutamic pyruvic transaminases. Am. J. Clin. Pathol., 28: 56-63.
CrossRefPubMedDirect Link - Santoni, G., F. Cantalamessa, O. Sagretti, M. Staffolani and M. Piccoli, 1999. Alterations of T cell distribution and functions in prenatally cypermethrin-exposed rats: Possible involvement of catecholamines. Toxicology, 138: 175-187.
CrossRef - Sayeed, I., S. Parvez, S. Pandey, B. Bin-Hafeez, R. Haque and S. Raisuddin, 2003. Oxidative stress biomarkers of exposure to deltamethrin in freshwater fish, Channa punctatus Bloch. Ecotoxicol. Environ. Saf., 56: 295-301.
CrossRefDirect Link - Shaker, N., G.A. Hassan, F.D. El-Nouty, Z. Abo-Elezz and G.A. Abd-Allah, 1988. In vivo chronic effect of dimethoate and deltamethrin on rabbits. J. Environ. Sci. Health B., 23: 387-399.
CrossRefDirect Link - Soderlund, D.M., J.M. Clark, L.P. Sheets, L.S. Mullin and V.J. Piccirillo et al., 2002. Mechanisms of pyrethroid neurotoxicity: Implications for cumulative risk assessment. Toxicology, 171: 3-59.
CrossRef - Srivastava, A.K. and N.N. Singh, 1981. Effect of alderin on carbohydrate metabolism in Indian catfish. Acta Pharmacol. Toxicol., 49: 266-269.
CrossRefDirect Link - Stevenson, D.E., J.P. Keher, K.L. Kolaja, E.F. Walborg and J.E. Klaunig, 1995. Effect of dietary antioxidants on dieldrin-induced hepatotoxicity in mice. Toxicol. Lett., 75: 177-183.
CrossRef - Symington, S.B., R.K. Frisbie, K.D. Lu and J.M. Clark, 2007. Action of cismethrin and deltamethrin on functional attributes of isolated presynaptic nerve terminals from rat brain. Pest. Biochem. Physiol., 87: 172-181.
CrossRef - Theophilidis, G., M. Benaki and E. Papadopoulou-Mourkidou, 1997. Neurotoxic action of six pyrethroid insecticides on the isolated sciatic nerve of frog (Rana ridibunda). Comp. Biochem. Physiol. C. Pharmacol. Toxicol. Endocrinol., 118: 97-103.
CrossRef - Tppel, A.L., 1968. Will antioxidant nutrients slow aging processes? Geriatrics, 23: 97-105.
PubMedDirect Link - Trinder, P., 1969. Determination of blood glucose using an oxidase-peroxidase system with a non-carcinogenic chromogen. J. Clin. Pathol., 22: 158-161.
CrossRefPubMedDirect Link - Wang, X., J. Wu, H. Xioa and Y. Shi, 2000. Effects of fenvalerate and phoxim insecticides on glutamate and gamma-aminobutyric acid immunoreactive cells in the central nervous system of rats. Wei Sheng Yan Jiu., 29: 1-3.
PubMedDirect Link - Wu, A. and Y. Liu, 2003. Prolonged expression of c-Fos and c-Jun in the cerebral cortex of rats after deltamethrin treatment. Brain Res. Mol. Brain Res., 110: 147-151.
CrossRef - Yamamoto, I., 1970. Mode of action of pyrethroids, nicotinoids and rotenoids. Annu. Rev. Entomol., 15: 257-272.
CrossRefDirect Link - Yousef, M.I., T.I. Awad and E.H. Mohamed, 2006. Deltamethrin-induced oxidative damage and biochemical alterations in rat and its attenuation by vitamin E. Toxicology, 227: 240-247.
CrossRefPubMedDirect Link - Zhao, X., X. Zhu and N. Shi, 1997. Effects of deltamethrin on the binding of glutamate receptor to rat cerebral cortical synaptosomal membrane. Zhonhua Yu Fan Yi Xue Za Zhi, 31: 157-159.
PubMedDirect Link