
Hashem Al-Sheikh
Department of Biology, College of Science, King Faisal University, AlHassa, Saudi Arabia
Hani M.A. Abdelzaher
Department of Biology, College of Science, Al-Jouf University, Sakaka, Saudi Arabia
Research Journal of Environmental Sciences
Year: 2012 | Volume: 6 | Issue: 6 | Page No.: 196-209







ABSTRACT
Pythium species are considered to be an aquatic fungus which occurs much abundantly in aquatic media. There are no sufficient efforts in studying the presence of them in water as compared with study in soil. For this reason, these fungi were studied in the waters of Dawmat Al-Jandal Lake which is essentially not yet attained their roles of biological and environmental studies in this country. In this study, Pythium biota has been investigated in Dawmat Al-Jandal lake, Saudi Arabia in order to clarify a diversity of this important fungus. Pythium species were isolated from water by baiting method with grass blades using Pythium selective medium. Sequencing of the internal transcribed spacer regions of ribosomal DNA (rDNA-ITS) including the 5.8 SrDNA of isolated pythia confirmed identification based on morphological characteristics. Fifty-nine isolates of Pythium spp. were obtained, of these, a total of three species and one group could be identified. Species isolated in the water of the lake were P. aphanidermatum, P. diclinum, sterile P. dissotocum and Pythium 'group P'. P. aphanidermatum was highly pathogenic to tomato seeds causing 100% damping-off but P. diclinum and the sterile isolate of P. dissotocum were moderately pathogenic, while Pythium "group P" was non-pathogenic to tomato seeds causing 60 and 0% damping-off, respectively. It is concluded that Pythium species is present in Dawmat Al-Jandal lake and some of these fungi appeared their virulence to tomato seedlings. It is possible that these pythia represent a risk factor in dissimilation of such microorganisms to the surrounded areas.
PDF Abstract XML References Citation
Received: October 15, 2012;
Accepted: January 09, 2013;
Published: February 16, 2013
How to cite this article
Hashem Al-Sheikh and Hani M.A. Abdelzaher, 2012. Occurrence, Identification and Pathogenicity of Pythium aphanidermatum, P. diclinum, P. dissotocum and Pythium "Group P" Isolated from Dawmat Al-Jandal Lake, Saudi Arabia. Research Journal of Environmental Sciences, 6: 196-209.
DOI: 10.3923/rjes.2012.196.209
URL: https://scialert.net/abstract/?doi=rjes.2012.196.209
DOI: 10.3923/rjes.2012.196.209
URL: https://scialert.net/abstract/?doi=rjes.2012.196.209
INTRODUCTION
Dawmat Al-Jandal Lake is located near Dawmat Al-Jandal city, northern part of Saudi Arabia (29°51’56″N, 39°51’20″E) and it has been formed during the eighth decade of the last century. The Lake formed due to accumulation of agricultural drainage water and rain water into the region. Formation of Dawmat Al-Jandal Lake has led to a number of changes in the existing microhabitats that a large number of different species of biota, flora and fauna became reestablished in such new ecological setting. The area thus opened several possibilities for various forms of development in tourism and recreational activities, fisheries and limited agricultural or soil reclaimatory activities. Before this study, no studies on the occurrence of Pythium species in the water Dawmat Al-Jandal lake has been done.
One of the most important aquatic fungus is the genus Pythium which was established by Pringsheim 1858 and placed in the family Saprolegniaceae. The genus was subjected to several taxonomical alternation through more than 14 decades (Berlese and De Toni, 1888; Butler, 1907). Now-a-days, the genus belongs to the Kingdom Stramenopile, excluded from the traditional “true fungi” of the Kingdom Myceteae. Aquatic media is necessary for production of zoospores and thus, Pythium spp. have been referred to commonly as belonging to aquatic molds. Consequently, standing water in fields, ditches, ponds, lakes and hydroponic systems are places that Pythium spp. can exist, multiply to great numbers and spread. Zoospores can swim for a short time (approximately 1/2 h) to a limited distance of three inches on the thin film of water around soil particles. Without free water they die rapidly or sometimes they can encyst by forming a thick wall and may survive up to 7 days in soil. Zoospores swim towards all directions and may be reached to new roots during irrigation. The zoospores are then attracted by chemicals to root tips where infection typically occurs. Natural or artificial movement of water infested with Pythium spp. provides a method by which rapid spread of these plant pathogens can occur over greater distances (Kucharek and Mitchell, 2000).
Keys for identification of Pythium species are based only on morphological basis (Middleton, 1943; Waterhouse, 1968; Van der Plaats-Niterink, 1981; Dick, 1990). Recently, morphological characteristics of a species are increasingly supported by molecular characteristics (Matsumoto et al., 1999; Levesque and de Cock, 2004; Kageyama et al., 2005; Tsukiboshi et al., 2007). Amplification of ribosomal genes by Polymerase Chain Reaction (PCR) is used for the genetic identification of many fungi (Masih and Paul, 2003). Additionally, The ribosomal nuclear DNA consists of transcribed and non-transcribed regions [ITS1 (internal transcribed spacer) and ITS2] can be amplified with the PCR method by using the universal primers ITS1 and ITS4 (Chen et al., 1992). Sequences of these parts are used to assist in identification of these species within the genus Pythium.
Species of Pythium species are cosmopolitan. They may present as saprophytes, muturalists and parasites and distributed in aquatic and terrestrial places (Van der Plaats-Niterink, 1981; Ichitani and Goto, 1982; Al-Sheikh and Abdelzaher, 2010a, b). Recently, some species of Pythium caused diseases to fishes, animals (Kitancharoen and Hatai, 1998; Helman and Oliver, 1999). Pythium diseases to human (Pythiosis) occurs in animals and human who contacting aquatic habits that contain Pythium insidiosum de Cock (Mendoza et al., 1987; Shenep et al., 1998).
Pythium spp. is commonly occurred in agricultured and non-agricultured soils of the world (Al-Sheikh and Abdelzaher, 2012). They present most abundantly in agricultured zones (Van der Plaats-Niterink, 1975), less commonly in non-agricultured or acid soils where the genus Trichoderma is the reason for their absence (Barton, 1958). Pythium species have also been isolated from different soil types of arable, pastures, forests, nurseries, marshes, swamps and water (Senda and Kageyama, 2006). Deserts, dry forests and salt marshes are generally poor in this taxon (Moharam, 2010). Species of Pythium dormans in soil as saprophyte and by the formation of resistant sexual structures (oospores) (Elnaghy et al., 2009). They germinate when humidity increase and by any other soil factor that may give it a competitive advantage against other soil microorganisms. They also become pathogenic and enter plant tissues; it spreads rapidly but can soon be overcome by competitors. Sparrow (1968) and Abdelzaher et al. (1995) postulated that "the ecology of fresh-water Pythium spp. has not received the same attention reached by that of soil pythia. Studies have been made on Pythium spp. associated with aquatic basins (Sanchez and Gallego, 2001). Until recently, studies on the occurrence of aquatic pythia in Saudi Arabia are scarce.
In Saudi Arabia, researching on this genus has begun recently. Little investigations were performed concerning some virulent Pythium spp. (Sunboul, 2001, 2006). International Migration Institute (IMI) (records for geographical unit Arabian Peninsula) has an isolate of P. aphanidermatum which isolated from Al-Madinah city and was parasitic on tomato plants in 1983 (Herb IMI 291421). Some researchers isolated some species of Pythium from the diseased and of healthy plants cultivated in Saudi Arabia, others have also isolated two species of Pythium from accumulated rainfall water in the Riyadh region, Saudi Arabia (El-Nagdy and Nasser, 2000) but descriptions and illustrations of the pure cultures of the isolated Pythium spp. were not obtained as well as identification based on morphological and molecular basis were not also supplied (Sunboul, 2001; Molan, 2009).
Occurrence of Pythium, especially in soil of near different plants, have been carried out by several researchers all over the world. In Saudi Arabia, scare studies has been performed for isolation and identification of this economic fungus. Except for few studies performed in certain places, no comprehensive study has been done concerning this line. Therefore, studying this unique fungus in Saudi Arabia is of importance and more researches should be done. In a series on study of Pythium in Saudi Arabia, Al-Sheikh and Abdelzaher (2012) have isolated some species of Pythium from rhizosphere soil of different cultivated plants. Generally, no data were available on isolation of Pythium from aquatic habitats in Saudi Arabia.
To receive more information, this study was done to get information about the presence of Pythium species in Dawmat Al-Jandal lake during winter of December of 2012. A complete description of the isolated species based on morphological and molecular criteria was provided. Pathogenicity of the isolated Pythium species using tomato was also tested. This study is one of a series of research on the unique fungus of Pythium in Saudi Arabia.
MATERIALS AND METHODS
Survey points: Collection were made from Dawmat Al-Jandal lake in Al-Jouf governorate, Saudi Arabia (29°51’56″N, 39°51’20″E), during December of 2012.
Surface water was collected from a depth of 30 cm at a distance of about 100 cm from the bank in 1 L sterilized plastic bottles and hermetically sealed.
Samples of water were collected in clean dry plastic bottles and at the laboratory, 30 mL poured in sterilized glass Petri-dishes and baited with 4 autoclaved wheat leaf pieces (0.5x1.0 cm). After incubation at 25°C for 5-7 days, the baits then were removed, washed several times with sterilized distilled water and blotted dry with sterile tissue paper. Baits were then placed on the colonial edges of a Petri-dish containing Nystatin Ampicillin Rifampicin Miconazole (NARM) medium (Gamo et al., 2004) for selective isolating of Pythium species. Baits were then incubated at 20°C for 3 days or until colonial appearance.
Morphological identification: NARM selective medium was used for purification and then identification. Identification using morphological characteristics was performed with the aid of keys of Middleton (1943), Waterhouse (1968), Van der Plaats-Niterink (1981) and Dick (1990).
Temperature-growth relations: Minimum, optimum and maximum temperatures of one each fungus are concluded on cornmeal agar (CMA; 17 gl-1, BD-BBLTM) inoculated with 5 mm dam discs from stock cultures on CMA plates. All plates must be incubated at 25°C for 24 h before starting growth determinations. Cardinal temperatures were evaluated at 5, 10, 15, 20, 25, 30, 35 and 40°C.
Molecular identification
DNA extraction, amplification, cloning and sequencing: Adequate mycelia of the tested pythia were got from the growth in V8 agar medium at 25°C for 7 days. Mycelia from the edge of Pythium colony grown on the plate were placed in 200 μL of the Reagent (Prep Man Ultra Sample Preparation; Applid Biosystems, CN, USA) in a 2.0 mL microcentrifuge tube in order to obtain extract of total genomic DNA. DNA of each fungus were then whirled for 30 sec and heated at 100°C for 10 min. DNA samples were then wind for 30 min in a centrifuge at 15000 g. The remaining supernatants were subjected to PCR. Amplification of The nuclear rDNA region of the Internal Transcribed Spacer (ITS), including the 5.8S rDNA, with the universal primers ITS4 (5′TCCTCCGCTTATTGATATGC3′) and ITS5 (5′GGAAGTAAAAGTCGTAACAAGG3′) was done. Primers of ITS1 (5′TCCGTAGGTGAACCTGCGG3′) and ITS2 (5′GCTGCGTTCTTCATCGATGC3′) were, sometimes, used according to White et al. (1990) and Matsumoto et al. (1999). The amplicons were 700-900 bp long. In some Pythium species, 563 bp of the cox II gene was amplified with the primer pair FM66 (5′TAGGATTTCAAGATCCTGC3′) and FM58 (5′CCACAAATTTCACTACATTGA 3′) (Martin, 2000). Amplification of the target template of sequencing was done using Thermal Cycler with a profile of pre PCR at 94°C for 5 min, then denaturation at 94°C for 1 min, another one minute of primer annealing at 55°C for the internal transcript spacer, after the final cycle, 52°C for gene cox II and elongation at 72°C, further two min for 40 cycles, with a 7 min extension at 72°C. Five micro liters of the PCR reaction mixture was loaded in 2% L03 (Takara Bio) agarose gel, electrophoresed at 100 V, 20-30 min and stained with ethidium bromide to ensure presence of PCR product. Sequencing templates were purified with GenElute PCR Clean-up kit following the manufacturer’s instructions (Sigma Chemical Co., St Louis, Missouri, USA). Sequencing was performed with BigDye Terminator v3.1 Cycle Sequencing Reaction kit (Applied Biosystems) using the same primers in the initial PCR step. After purification of the sequencing reaction mixture by using ethanol precipitation, it was then run on ABI 3100 DNA Sequencer (Applied Biosystems). Some PCR products were cloned in the pT7Blue T-vector (Takara Bio) with the Ligation kit (Takara Bio) according to the manufacturer’s instruction. The cloned ITS region was amplified using M13M4 (5′GTTTTCCCAGTCACGAC3′) and M13Rv (5′CAGGAAACAGCTATGAC3′) primers. The PCR products were purified using the Gene Elute PCR cleaning kit (Sigma, Ronkonkoma, NY, USA). The BigDyeTM Terminator Cycle Sequencing Ready Reaction kit *Applied Biosystems) was used for cycle sequencing with primers M13M4 and M13Rv according to the manufacturer’s instructions. The sequencing reaction mixture was purified by ethanol precipitation and run on an ABI 3100 DNA sequencer (Applied Biosystems). The consensus sequences were generated based on the forward and reverse primer sequences.
Pathogenicity test: Inocula of tested species were developed to be suitable for the present study. Five gram of grass blade leaf segments and 2 g glucose were moistened by adding distilled water (10 mL) each in 250 mL conical flask. After sterilization, each conical flask containing the above mentioned mixture was supplemented with 3 colonized agar disks (7 mm dam) by Pythium spp. and incubated at 25°C for 10 days. Inoculums concentration of 2.5% was prepared by mixing one gram of colonized grass leaf segments in the conical flask with 50 g of dried clay sandy soil using a sterilized mortar and pestle. The 2.5 g of this mixture were then mixed with 97.5 g of sterilized clay loam soil. Tomato seeds (Solanum lycopersicum L.) were surface sterilized by using sodium hypochlorite 2% for 3 min and then washing well by sterilized H2O followed by 1 min in 70% ethanol and finally three times using sterilized distilled water. Pre germination test was done to select viable seeds for the pathogenicity test. Twenty tomato seeds were planted in each pot to perform the pathogenicity test. The experiments were tested in a growth chamber at 25°C with illumination of 12 h photoperiod (91 μmol m-2 S-1). Damping-off percentage was determined by the difference between tomato seeds emergence of the infested pot and the control one.
RESULTS
Occurrence and distribution of Pythium spp.: All baits produced Fungi. Grass leaf blades baits were very useful for isolating different species of Pythium. Pythium aphanidermatum, P. diclinum, sterile P. dissotocum and Pythium “group P” were isolated using this bait.
Morphological identification: Fifty-nine isolates of Pythium spp. were obtained from the studied area in Dawmat Al-Jandal lake, Al-Jouf governorate, Saudi Arabia (Table 1). Of these, a total of six species could be identified on the basis of morphological and molecular criteria. The remaining isolates, which lacked sexual structures, have filamentous sporangia and one of them was identified molecularly as P. dissotocum others which lacked sexuality and have proliferated sporangia were identified as Pythium “group P”.
Pythium aphanidermatum (Edson) Fitzp.(JU00010) (Fig. 1): Mycelial growth of this fungus showed heavy with cottony appearance especially on rich medium such as Potato Dextrose Agar (PDA). Main hyphae were up to 10 μm wide. Zoosporangia consisted of terminal complexes of swollen hyphal branches of varying length and up to 22 μm wide. Zoospores were formed at 15-35°C. Encysted zoospores had a diameter of 12 μm. Oogonia were terminal, globose, smooth and of (20-) 22-25(-26) μm (av. 24 μm) dam. Antheridia were mostly intercalary, sometimes terminal, broadly sac-shaped, 11-15 μm long and 9-15 μm wide, 1(-2) per oogonium and monoclinous or diclinous. Oospores were not filling oogonia (aplerotic) (19-)20-24 μm (av. 22 μm) in dam and their walls are 1-2 μm thick. This fungus was deposited in Department of Biology, College of Science, Al-Jouf University, Saudi Arabia with number JU00010.
Pythium diclinum Tokunaga (JU00020) (Fig. 2): Mycelial growth of this fungus showed submerged growth on poor medium such as CMA and with little aerial mycelia on the rich medium. Main hyphae was up to 7 μm wide. Sporangia were filamentous or non-inflated, sometimes branched. Zoospores formed at 20-25°C; vesicles are varying from very small containing 2 zoospores to big containing many zoospores. Encysted zoospores were about 7 μm dam. Oogonia were globose, to subglobose and sometimes ovoid, smooth, mostly terminal or subterminal, occasionally intercalary, 18-24 μm (av. 22 μm) dam. Antheridia were typically diclinous, 1-2 per oogonium, about 12x15 μm. Antheridial stalks not branched. Oospores were single, aplerotic; 16-20 μm (av. 19) dam, wall up to 3 μm thick.
| Table 1: | Linear growth of Pythium spp. isolated from water of Dawmat Al-Jandal lake at various temperatures |
 | |
| *Growth is the mean of 5 replicates and no significant difference in diameter within replicates of each measurement was observed | |
 | |
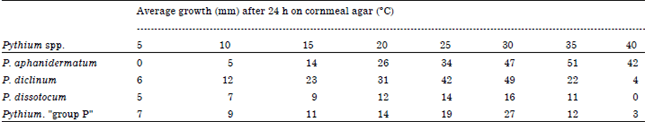
| Fig. 1(a-f): | Morphology of Pythium aphanidermatum (JU00010), (a) A toruloid zoosporangium with a vesicle (arrow), (b) A vesicle contains differentiated zoospores, (c) Young oogonium and young intercalary diclinum antheridium, (d) Intercalary antheridia (arrows) and (e, f) Aplerotic oospores (arrows). Bar on photo (f) is equal 20 μm and is applicable to the rest photos |
This fungus was deposited in Department of Biology, College of Science, Al-Jouf University, Saudi Arabia with number JU00020.
Pythium dissotocum Drechsler (JU00030) (Fig. 3): Colonies on corn meal agar were submerged, without a special pattern and with little aerial mycelia. Main hyphae was up to 6 μm wide.
 | |
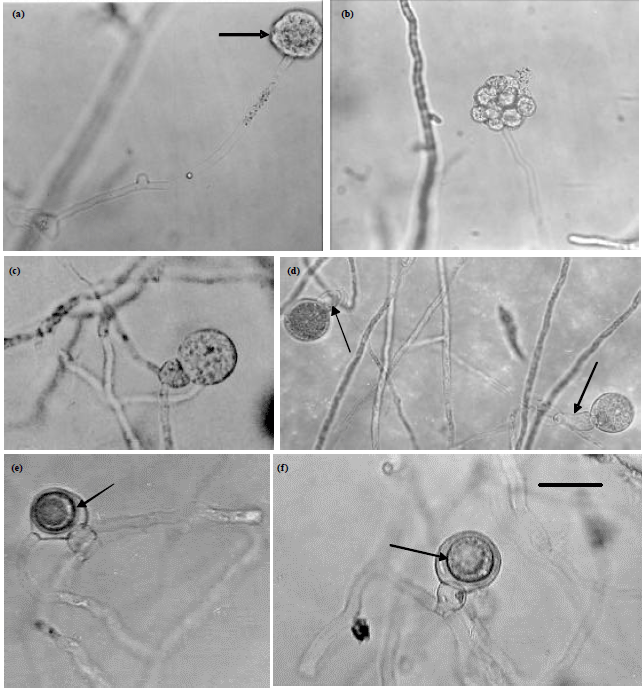
| Fig. 2(a-f): | Morphology of Pythium diclinum (JU00020), (a) A filamentous zoosporangium with a vesicle (arrow), (b) Intercalary oogonium and diclinous antheridium, (c, d) Terminal oogonia and diclinum antheridia (arrows) and (e) An aplerotic oospore (arrows). Bar on photo (4) is equal 20 μm and is applicable to photos 1, 2, 3 whereas bar on photo 5 is equal 20 μm and it is for its photo only |
Sporangia were filamentous forming very little inflated dendroid structures. Zoospores formed at 20°C. Encysted zoospores were up to 8 μm dam. This isolate failed to produce sexual structures and it was deposited in Department of Biology, College of Science, Al-Jouf University, Saudi Arabia with number JU00030.
Pythium “group P” Plates Niterink (JU00040) (Fig. 4): Colonies on corn meal agar were submerged, with radiate pattern and with some aerial mycelia. Main hyphae was up to 8 μm wide.
 | |
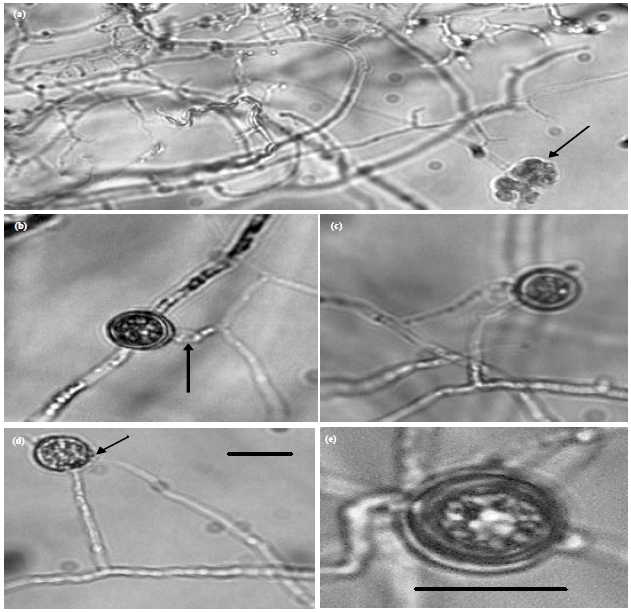
| Fig. 3(a-c): | Morphology of a sterile isolate of Pythium dissotocum (JU00030), (a) Terminal hyphae, (b) A young vesicle (arrow), (c) Young and old vesicles contains differentiated zoospores (arrow), (d) Vesicles and evacuation tube after zoospores liberated from the vesicle (arrow) and (e) Liberation of zoospores from the vesicle. Bar on photo (a) is equal 20 μm and is applicable to the rest photos |
Sporangia were proliferating internally by forming a new sporangium inside the old one, often 20-30 μm dam, 25 μm on average, mostly forming short evacuation tubes, sometimes germinated by 1 or 2 germ tubes. Zoospores produced at 20-25°C. Encysted zoospores were up to 12 μm dam. No sexual reproduction observed. This fungus was deposited in Department of Biology, College of Science, Al-Jouf University, Saudi Arabia with number JU00040.
Temperature growth relations: P. aphanidermatum showed temperature maximum for growth higher than that of the other Pythia studied.
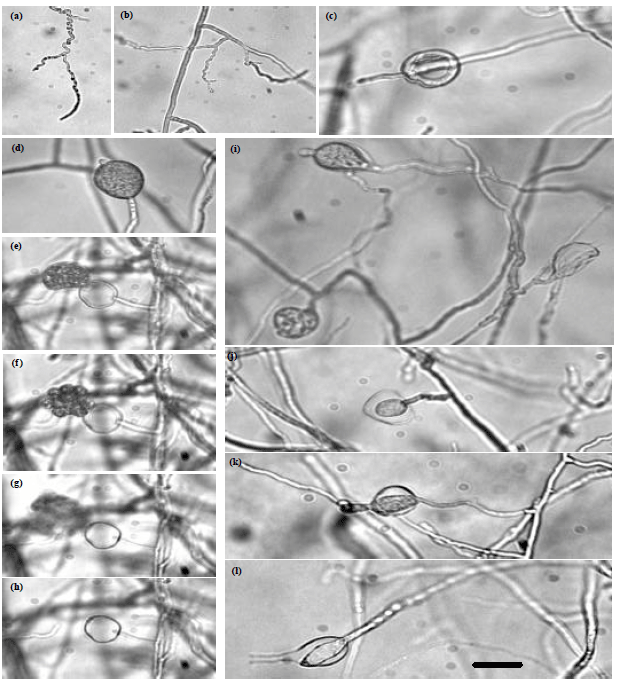 | |
| Fig. 4(a-h): | Morphology of Pythium "group P" (JU00040), (a, b) Hyphae, (c) Proliferated zoosporangium, (d) Young zoosporangium, (e-h): Stages of vesicle formation and zoospores liberation and (i-l): Internal proliferation of zoosporangia. Bar on photo (h) is equal 20 μm and is applicable to rest photos |
The optimum temperatures for all the other Pythium spp. investigated lied between (25 and 30°C) and the minimum temperature supporting mycelial growth of these spp. was around 5°C except P. aphanidermatum which started to grow at 10°C (Table 1).
Molecular identification: The sequence of JU00010 was closely related with that of P. aphanidermatum (AY598622.1) with 100% similarity. The sequence (with the aid of TA-cloning) of JU00020 was closely related with that of P. diclinum (EF153676) with 100% similarity.
 | |
| Fig. 5: | Nuclear rDNA region of the internal transcribed spacer (ITS), including the 5.8S rDNA, of P. aphanidermatum, P. diclinum and P. dissotocum |
The sequence of JU00030 was closely related with that of P. dissotocum (AY598631) with 99% similarity. Sequencing of Pythium “group P” was though to be Pythium helicoides, therefore, sequencing of cox II gene was done. It was confirmed that this isolate of group P was not P. helicoides.
Pathogenicity test: Pre-emergence damping-off test was performed in pots using 2.5% inoculum concentration of the tested Pythia added to soil and incubated for 2-3 weeks before cultivated tomato seeds. P. aphanidermatum was highly pathogenic to tomato seeds causing 100% damping-off. P. diclinum and P. dissotocum were moderately pathogenic producing 65% damping-off while Pythium "group P" was non-pathogenic to tomato seeds (Fig. 5, 6).
DISCUSSION
This is the first study dealt with aquatic pythia in Dawmat Al-Jandal lake, Dawmat Al-Jandal city, Al-Jouf governorate, Saudi Arabia. The species obtained in this study were; P. aphanidermatum, P. diclinum, sterile P. dissotocum and Pythium “group P”. P. diclinum and the sterile P. dissotocum have filamentous sporangia, while P. aphanidermatum has inflated lobulate sporangia and Pythium “group P” forms proliferated sporangia. P. aphanidermatum was isolated several times from aquatic habitats such as irrigation water (Abdelzaher, 1999; Moustafa et al., 2010). P. diclinum was originally isolated from a paddy field in Japan (Van der Plaats-Niterink, 1981). P. dissotocum has also been isolated from water in the formal USSR and the Netherlands (Van der Plaats-Niterink, 1981). Pythium “group P” was isolated many times from aquatic habitats (Van der Plaats-Niterink, 1981; Abdelzaher, 1999; Abdelzaher et al., 1995). Therefore, result of isolation of such species from water here is in harmony with the previous findings.
Results appear that grass leaves segments was very useful as baits for isolation of Pythium spp. It is worth to mention that, the results (Fig. 7) of the rDNA-ITS and cox II sequence analysis of the Pythium isolates agreed with the morphological features of each isolate and were of great helpful in confirming identification and overcome hesitating and doubtfulness.
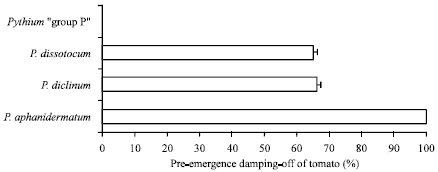 | |
| Fig. 6: | Pre-emergence damping-off of tomato seedlings grown in clay sand soil infested with tested Pythium spp. Bars indicate standard errors of 100 measurements |
 | |
| Fig. 7: | Pre-emergence damping-off of tomato seedlings grown in clay sandy soil in plastic pots infested with Pythium aphanidermatum (2), P. diclinum (3), P. dissotocum (4). Control pot (1) does not contain any Pythium inocula |
Results here appeared that three out of four pythia isolated from water of Dawmat Al-Jandal lake proved ferocity against tomato seedlings (Fig. 5, 6). As shown in results, P. aphanidermatum was highly pathogenic causing 100% damping-off, P. diclinum and P. dissotocum caused 65% damping-off to tomato germinating seeds. Therefore, the presence of such fungi that cause some diseases of crop plants is a source of danger, especially if the lake water is used for irrigation. Care must be taken of pollution of the agricultural fields by using water of the lake. In previous studies, the pathogenicity of the isolated Pythium spp. in this study was proven. P. aphanidermatum can cause root rot, damping-off, stalk and rhizome rot, soft rot, fruit rot or cottony blight of many economic plants (Van der Plaats-Niterink, 1981). P. diclinum caused severe damping-off of wheat in Egypt (Abdelzaher, 2004). P. dissotocum was pathogenic to sugar cane, pecan and peach trees (Van der Plaats-Niterink, 1981). Therefore, above mentioned previous studies support results of the present investigation.
It is worth mentioning that Dawmat Al-Jandal Lake is an artificial lake created by nearly 30 years in a desert area of continental climate. Explanation of occurrence of Pythium spp. which possess zoospores floating in the lake is of important from the environmental perspective. Where this fungus is not air borne, it could be explained that entry of it to the lake by either droppings of migratory birds or wash of agricultural soil adjacent to the lake.
CONCLUSION
Experimental scientific explanation for this phenomenon has not carried out in this study and needs further investigation.
ACKNOWLEDGMENTS
This study was partially supported by King Faisal University (small grant No. 131001). We are grateful for the partial support of this project by Eid and Otto Internationale, S.A. P.O. Box 793 Belize City, Belize, C.A.
REFERENCES
- Abdelzaher, H.M.A., 2004. Occurrence of damping-off of wheat caused by Pythium diclinum tokunaga in El-Minia, Egypt and its possible control by Gliocladium roseum and Trichoderma harzianum. Archiv. Phytopathol. Plant Prot., 37: 147-159.
CrossRef - Abdelzaher, H.M.A., T. Ichitani, M.A. Elnaghy, S.K.M. Hassan and E.M. Fadl-Alla, 1995. Materials for Pythium flora of Japan. X. Occurrence, identification and seasonality of Pythium spp. in three pond waters and mud soils in Osaka. Mycoscience, 36: 71-85.
Direct Link - Al-Sheikh, H. and H.M.A. Abdelzaher, 2010. Differentiation between two isolates of Pythium ultimum var. ultimum isolated from diseased plants in two different continents. J. Biol. Sci., 10: 306-315.
CrossRefDirect Link - Al-Sheikh, H. and H.M.A. Abdelzaher, 2012. Materials for Pythium flora of Saudi Arabia (I) Occurrence, pathogenicity and physiology of reproduction of Pythium aphanidermatum (Edson) Fitzp. isolated from north and east regions of Saudi Arabia. Res. J. Microbiol., 7: 82-100.
CrossRefDirect Link - Al-Sheikh, H. and H.M.A. Abdelzaher, 2010. Isolation of Aspergillus sulphureus, Penicillium islandicum and Paecilomyces variotii from agricultural soil and their biological activity against Pythium spinosum, the damping-off organism of soybean. J. Biol. Sci., 10: 178-189.
CrossRefDirect Link - Barton, R., 1958. Occurrence and establishment of Pythium in soils. Trans. Br. Mycol. Soc., 41: 207-222.
CrossRef - Chen, W., R.W. Schneider and J.W. Hoy, 1992. Taxonomic and phylogenetic analyses of ten Pythium species using isozyme polymorphism. Phytopathology, 82: 1234-1244.
CrossRef - El-Nagdy, M.A. and L.A. Nasser, 2000. Occurrence of zoosporic and terrestrial fungi in accumulated rainfall water in the Riyadh region (Saudi Arabia). Fungal Diversity, 5: 175-183.
Direct Link - Helman, R.G. and J. Oliver, 1999. Pythiosis of the digestive tract in dogs from Oklahoma. J. Am. Anim. Hosp. Assoc., 35: 111-114.
PubMed - Ichitani, T. and H. Goto, 1982. Distribution of Pythium zingiberum causing rhizome rot in ginger growing and its surrounding uncultivated soils. Ann. Phytopath. Soc. Japan, 48: 647-676.
Direct Link - Kageyama, K., A. Nakashima, Y. Kajihara, H. Suga and E.B. Nelson, 2005. Phylogenetic and morphological analyses of Pythium graminicola and related species. J. Gen. Plant Pathol., 71: 174-182.
CrossRefDirect Link - Kitancharoen, N. and K. Hatai, 1998. Some biochemical characteristics of fungi isolated from salmonid eggs. Mycoscience, 39: 249-255.
CrossRef - Levesque, C.A. and W.A. de Cock, 2004. Molecular phylogeny and taxonomy of the genus Pythium. Mycol. Res., 108: 1363-1383.
PubMedDirect Link - Martin, F.N., 2000. Phylogenetic relationships among some Pythium species inferred from sequence analysis of the mitochondrially encoded cytochrome oxidase II gene. Mycologia, 92: 711-727.
Direct Link - Masih, I. and B. Paul, 2003. Pythium regulare sp. Nov., isolated from the Canary Islands, its taxonomy, its region of rDNA and comparison with related species. Curr. Microbiol., 47: 309-313.
CrossRef - Matsumoto, C., K. Kageyama, H. Suga and M. Hyakumachi, 1999. Phylogenetic relationships of Pythium species based on ITS and 5.8S sequences of the ribosomal DNA. Mycoscience, 40: 321-331.
CrossRefDirect Link - Mendoza, L., L. Kaufman and P. Standard, 1987. Antigenic relationship between the animal and human pathogen Pythium insidiosum and nonpathogenic Pythium species. J. Clin. Microbiol., 25: 2159-2162.
PubMed - Middleton, J.T., 1943. The taxonomy, host range and geographic distribution of the genus Pythium. Memories Torrey Bot. Club, 20: 1-171.
Direct Link - Molan, Y.Y., 2009. Detection of presumptive mycoparasites in soil placed on host-colonized agar plates in Riyadh Region, Saudi Arabia. Asian J. Plant Pathol., 3: 22-26.
CrossRefDirect Link - Van der Plaats-Niterink, A.J., 1975. Species of Pythium in the Netherlands. Netherlands J. Plant Pathol., 81: 22-37.
CrossRef - Shenep, J.L., B.K. English, L. Kaufman, T.A. Pearson and J.W. Thompson et al., 1998. Successful medical therapy for deeply invasive facial infection due to Pythium insidiosum in a child. Clin. Infect. Dis., 27: 1388-1393.
PubMed - Sunboul, Y.H., 2006. Solarizaton of commercial peat in transparent polyethylene bags and its effect on survival of some plant pathogenic fungi. JKAU Sci., 18: 1-11.
Direct Link - Tsukiboshi, T., Y. Chikuo, Y. Ito, Y. Matsushita and K. Kageyama, 2007. Root and stem rot of chrysanthemum caused by five Pythium species in Japan. J. Gen. Plant Pathol., 73: 293-296.
CrossRef - White, T.J., T.D. Bruns, S.B. Lee and J.W. Taylor, 1990. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In: PCR Protocols: A Guide to Methods and Applications, Innis, M.A., D.H. Gelfand, J.J. Sninsky and T.J. White (Eds.), Academic Press, San Diego, CA, USA, ISBN-13: 9780123721808, pp: 315-322.
CrossRefDirect Link - Sanchez, J. and E. Gallego, 2001. Pythium spp. present in irrigation water in the Poniente region of Almeria (south-eastern Spain). Mycopathologia, 150: 29-38.
Direct Link