
Homa Mahmoodzadeh
Department of Biology, Faculty of Sciences, Islamic Azad University, Mashad Branch, Iran
Forugh Abbasi
Department of Biology, Faculty of Sciences, Islamic Azad University, Mashad Branch, Iran
Yalda Ghotbzadeh
Department of Biology, Faculty of Sciences, Islamic Azad University, Mashad Branch, Iran
Research Journal of Environmental Sciences
Year: 2011 | Volume: 5 | Issue: 5 | Page No.: 486-492







ABSTRACT
To determine the allelopathic potential of root exudate from early developmental stage of rice (Oryza sativa L.), 6-days-old seedlings of two cultivars were grown with 3-days-old Hedgemustard seedlings in petri dishes under controlled condition. There were 4 treatments (2, 4, 8 and 10 days). Also rice leachate was used for germination test of the weed. Rice lichate caused pronounced inhibitory effect on seed germination and germination rate of receptor plants. Root exudates of rice cultivars also inhibited root and shoot growth and fresh weights of seedlings of receiver plants. Effectiveness of cv. kalat was more than cv. shomal. Hence, it could be concluded that the rice root exudates and rice leachate contain water-soluble allelochemicals which could inhibit the seed germination and reduce seedling growth of Hedgemustard.
PDF Abstract XML References Citation
Received: December 07, 2010;
Accepted: February 14, 2011;
Published: March 25, 2011
How to cite this article
Homa Mahmoodzadeh, Forugh Abbasi and Yalda Ghotbzadeh, 2011. Allelopathic Effects of Root Exudate and Leaching of Rice Seedlings on Hedgemustard (Sisybrium officinale). Research Journal of Environmental Sciences, 5: 486-492.
DOI: 10.3923/rjes.2011.486.492
URL: https://scialert.net/abstract/?doi=rjes.2011.486.492
DOI: 10.3923/rjes.2011.486.492
URL: https://scialert.net/abstract/?doi=rjes.2011.486.492
INTRODUCTION
Rice has been extensively studied with respect to its allelopathy as part of a strategy for sustainable weed management, such as breeding allelopathic rice strains (Olofsdotter, 2001; Takeuchi et al., 2001). A large number of rice varieties were found to inhibit the growth of several plant species when grown together under field and/or laboratory conditions (Azmi et al., 2000). These findings suggest that rice may produce and release allelochemical(s) into the neighboring environment, thus encouraging the exploration of allelochemicals in rice.
Rice residues and their aqueous extracts suppressed the growth of lettuce and Phalaris minor (Khan et al., 2001) and aqueous and organic solvent extracts of rice plants inhibited the growth of several plant species (Das and Goswami, 2001; Ebana et al., 2001; Mattice et al., 1998; Kato-Noguchi, 2002). Several phenolic acids, such as p-hydroxybenzoic acid, vanillic acid p-coumaric acid and ferulic acid were found in aqueous extracts of rice straws, roots and residues (Chung et al., 2001). Phenolic acids were also found in rice root exudates. Although phenolic acids are often mentioned as putative allelochemicals, it is unclear whether concentrations of phenolic acids measured in rice ecosystems are sufficient for causing growth inhibition of neighboring plant species (Olofsdotter, 2001).
Zhang et al. (2005) identified potent allelopathic rice germplasm for breeding the new rice varieties with allelopathic potential. Yiqing et al. (2005) focussed on the evaluation of allelopathic wild rice species germplasm resources for barnyardgrass control, to further facilitate research and exploitation of these breeding materials to improve rice yield, quality and resistance to weeds.
He et al. (2006) detected the diethyl ether extracts from the root exudates of two rice accessions, allelopathic rice (PI312777) and non-allelopathic (rice Lemont). Seedlings and the individual compounds identified by comparing their mass spectra with those from NIST and Wiley Library of mass spectral database. Sixty-three compounds were detected in root exudates of PI312777 and 77 compounds in Lemont. The substances were terpenoids, phenols (quinones), aldehydes (ketones), heterocyclic alcohols, ethers, hydrocarbons etc. (He et al., 2006). Hiroshi (2008) examined the effect of husk extracts of wild rice spp. on root and shoot growth of lettuce, barnyard grass and Eclipta thermalis (false daisy). The results suggested that the husk extracts of O. glumaepatula contain water soluble allelochemicals that inhibit root growth of lettuce but promote shoot growth of false daisy and ethyl acetate soluble allelochemicals that inhibit root growth of barnyard grass. It is not clear, however, whether these compounds are released from roots of living rice plants. Olofsdotter et al. (1995) claimed that extraction of plant tissue by water might force abnormal allelopathic effects to occur.
This study described the assessment of the allelopathic potential of root exudates from rice plants during their early developmental stage and allelopathic potential of rice leaching by using Petri dish bioassay under laboratory condition.
MATERIALS AND METHODS
The experiment was conducted in Faculty of Sciences, Islamic Azad University, Mashad Branch, Iran in 2009.
Two cultivars of rice (Oryza sativa L.), cv. Kalat and Shomal were chosen for bioassay as donor plants. About 250 plants of rice were planted in plastic pots and irrigated with tap water at 3 days interval and leaching were collected through draining every week. These leaching were used to study their effect on seed germination.
Germination test: Seeds of Sisybrium officinale were supplied from Agricultural Research Center of Khorasan. The seeds were surface sterilized with 5% sodium hypochloride for 10 min then rinsed with the distilled water for several times to remove excess of chemical. Ten uniform and surface sterilized seeds (5% sodium hypochlorite for 15 min) of weed plant were kept for germination in sterilized petri-dishes lined double with blotting paper and moistened with 5 mL of different concentrations of leachating solutions of rice (25, 50 and 75%). Each treatment had three replicas (total number of test seeds: 10 x3 = 30). One treatment was run as control with distilled water only. The petri-dishes were maintained under laboratory conditions (room temperature 25°C at mid day and diffused light during day) for one week. Equal volume of distilled water was added in the dishes when moisture content of the blotting paper declined. Germination rate was determined by counting the number of germinated seeds at 24 h intervals over a 7 day period. Final germination percentage was recorded after 7 days.
Seedling growth: The rice seeds were sterilized in an aqueous solution of 25 mM sodium hypochlorite for 15 min and rinsed in distilled water four times. Then, the seeds were allowed to germinate on two sheets of moist filter paper at 25°C and 12 h photoperiod. After 6 days, uniform seedlings were selected and used for bioassay. It is difficult to use weed species as test plants under laboratory conditions, since their germination is at low rate and inconsistent (Inderjit and Olosfsdotter, 1998). Hedgemustard (Sisymbrium officinale L.) was chosen for bioassay as receiver plant because of its known germination behavior. Seeds of the plant were sterilized, rinsed and germinated as described above. After being kept in the daily cycles for 3 days, uniform seedlings were chosen for bioassay.
Rice seedlings were transferred, in groups of ten, to 9 cm Petri dishes which contained two sheets of filter paper (No. 2) moistened with 10 cm3 of 3 mM phosphate buffer (pH 6.0). Then, receiver plants (ten seedlings of Hedgemustard) was arranged on the same filter paper in the petri dishes and incubated under the same conditions. The phosphate buffer was added in 6 h intervals. After 2, 4, 8 and 10 days, the shoot and root lengths and fresh weight of the receiver plants were measured. Control seedlings were grown alone without rice seedlings. Root and shoot length of the germinated seed was measured using a millimeter ruler.
All experimental treatments were replicated three times in complete randomized block designs.
Statistical analysis: To detect the significance of differences of variables statistical analysis was performed employing one way ANOVA test using MINITAB software.
RESULTS AND DISCUSSION
The allelopathic effect of rice (Oryza sativa) on the germination percentage of Hedgemustard is shown in Fig. 1. It is obvious that the leaching of rice inhibited the germination of plants. The maximum seed germination percentage was shown in the control where no leaching was used, which was 50% for Hedgemustard. The highest inhibitory effect was found in Hedgemustard at 50% treatment of Kalat cultivar which was 12%. Also, germination rate was inhibited by leaching of two rice cultivars (Fig. 2, 3).
According to the test solution of (Weidenhamer et al., 1987), 3 mM phosphate buffer (pH 6.0) was chosen as the test solution for the experiment 2. It was pointed out that research in allelopathy should design to eliminate the effects of the competitive interference (He et al., 2006). In our experiments, each cultivar of 6-days-old rice seedlings (2 cultivars, donor plants) was grown with 3-days-old Hedgemustard seedlings (receiver plant) in petri dishes without interspecies competition for light and nutrients as well as water. All rice cultivars inhibited the root and shoot growth and fresh weight of receiver plant. However, the inhibition rate of both root and shoot growth and fresh weight differed with combinations of rice cultivars. The root length of Hedgemustard, in treatments of 2, 4, 8 and ten days of cv. Shomal was 95.8, 56.6, 76.5 and 94.5% those of the control plants, respectively (Fig. 5) and the shoot length of Hedgemustard seedlings was 44.1, 41.2, 64.1 and 89.2% those of the control plants, respectively (Fig. 4). For cv.Kalat treatments, root length was 89.4, 63.3,80.8 and 100% (Fig. 5) and shoot length was 42.9, 54, 57.8 and 100%, respectively (Fig. 4). Also treatments of 2, 4, 8 and 10 days of rice cultivars (Fig. 6) reduced the fresh weight of seedlings of Hedgemustard. Thus, the growth of roots and shoots and fresh weights of receiver plant was inhibited by the presence of rice seedlings of all cultivars tested. Although there was difference in the sensitively between roots and shoots in the receiver plant. Inhibition indexes (Table 1) were calculated from average inhibition rate of root length, shoot length and fresh weight.
It has not always been well accepted that chemical compounds produced by plants can escape into the environment and adversely affect other plants. This phenomenon termed “allelopathy” usually occurs in plant communities and is thought to be a possible way for plants to influence the growth and development of competing plants. In this study we evaluated the allelopathic potential of the rice using a weed (Hedgemustard).
Several genes because of the involvement of several chemicals in combination with the production and release of these chemicals may control allelopathic ability. In fact, the allelopathic activity was reported in various rice of both traditional and improved cultivars from different countries (Dilday et al., 1994).
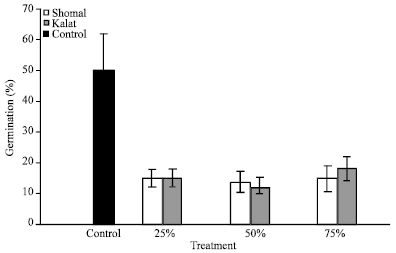 | |
| Fig. 1: | Seed germination percentage of Sisybrium officinale under different treatments of rice leachating solutions |
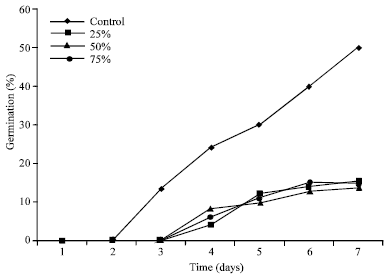 | |
| Fig. 2: | Seed germination rate of Sisybrium officinale under different treatments of rice leaching solutions (shomal cultivar) |
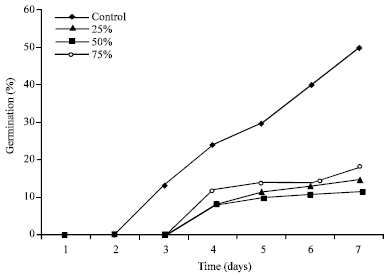 | |
| Fig. 3: | Seed germination rate of Sisybrium officinale under different treatments of rice leachating solutions (Kalat cultivar) |
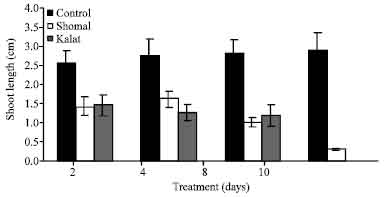 | |
| Fig. 4: | Effects of rice seedlings on shoot growth of Sisybrium officinale seedlings |
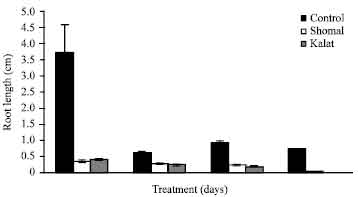 | |
| Fig. 5: | Effects of rice seedlings on root growth of Sisybrium officinale seedlings |
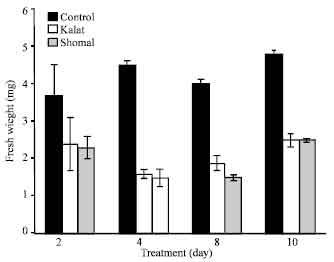 | |
| Fig. 6: | Effects of rice seedlings on fresh weight of Sisybrium officinale seedlings |
| Table 1: | Inhibition index of rice seedlings on Sisybrium officinale seedlings. The inhibition indexes were calculated on average of inhibition rate of root length , shoot length and fresh weight. Inhibition rate of root length , shoot length and fresh weight, respectively was scored so that those of control plants were 100% |
 | |
Dekker and Meggitt (1983) have found that most allelochemicals were released during germination and early developmental stage of crop plants when the crops were most sensitive for competition. During the period, weeds also establish and create the basis for later major weed problems. Thus, early developmental stage of crop plants might determine the possible crop yield at the end of the season. Using laboratory Petri dish bioassay, 6-days-old rice seedlings were shown to affect the growth of roots, shoots and fresh weights of Hedgemustard seedlings. These results suggest that the 6 to 9-days-old rice seedlings may produce and release alleochemical(s) into the environment and inhibit growth of neighbouring plants. Inderjit and Olosfsdotter (1998) proposed that laboratory bioassay is one of the most important parts of research in rice allelopathy because bioassays under controlled environments permit distinctions to be made between allelopathic effects and interference by other means. Yiqing et al. (2005) showed Two wild rice accessions of S46 (Oryza barthii) and S72 (O. rufipogon) significantly reduced the plant height and dry weight of barnyardgrass. While, a wild rice accession of S37 (O. longistaminata) reduced the barnyardgrass germination rate significantly. The inhibitory effects of wild rice on the weed were greater during the rice elongation stage than at the reviving stage Yiqing et al. (2005). Farooq et al. (2008) studied the allelopathic potential of different plant parts of rice against wheat, oat, barley and berseem. In Lab bioassay, rice stem extract reduced the germination, energy of germination, seedling length and seedling dry weight of test crops. The number of roots was decreased in all crops except wheat. Roots and leaves extract also inhibited the seedling germination and growth. Nonetheless they also had promotive effects, suggesting the presence of growth promoting allelochemicals Farooq et al. (2008). The stability of 9 allelopathic rice accessions against barnyard grass was investigated with different sowing-dates approach in paddy field. The results showed that the inhibitory rates of rice on barnyard grass varied significantly with test rice accessions and barnyard grass sowing-dates. The inhibitory effects of 3 hybrid rice accessions on barnyard grass showed higher stability than other 7 inbred rice accessions Hu et al. (2008). Fudou et al. (2008) investigated the effects of 5 accessions of Oryza Longistaminata and Oryza sativa [PI 312777, RD 23, O. longistaminata, O. rufipogon and F1 (O. longistaminataxRD23)] at different leaf stages on the growth of barnyardgrass (Echinochloa crus-galli (L.) Beauv.). The rice accessions suppressed the weed growth by allelopathic activity more than competition for growth resources Fudou et al. (2008). Understanding of the chemical basis of the allelopathic system in rice plants as well as the field experiment of the allelochemicals are also essential (Inderjit and Dakshini, 1994). These studies are in support to our results. To clarify the chemical basis of such allelopathic system, purification and identification of the allelochemical(s) released from cv. Kalat and Shomal which possess greatest allelopathic potential is now underway.
CONCLUSION
We studied the allelopathic potential of two cultivars of rice against Hedgemustard. In Lab bioassay, rice root exudates reduced the rate and percentage of germination seedling length and seedling fresh weight of test plant. Inhibition index showed that cv. Kalat was more effectiveness on reduced growth of the weed. Our results suggested that rice allelopathy is involved in retarding the germination and growth of Sisybrium officinale.
REFERENCES
- Azmi, M., M.Z. Abdullah and Y. Fuzii, 2000. Exploratory study on allelopathic effect of selected Malaysian rice varieties and rice field weed species. J. Trop. Agric. Food Sci., 28: 39-54.
Direct Link - Dekker, J. and W.F. Meggitt, 1983. Interference between velvetleaf (Abutilon theophrasti Medic.) and soybean (Glycine max (L.) Merr.). I. Growth. Weed Res., 23: 91-101.
CrossRef - Dilday, R.H., J. Lin and W. Yan, 1994. Identification of allelopathy in the USDA-ARS rice germplasm collection. Aust. J. Exp. Agric., 34: 907-910.
CrossRefDirect Link - Ebana, K., W. Yan, R.H. Dilday, H. Namai and K. Okuno, 2001. Variation in the allelopathic effect of rice with water soluble extracts. Agron. J., 93: 12-16.
Direct Link - Fudou, Z., L. Tianlin, S. Qinli, G. Yiqing, X. Peng, H. Fengyi and T. Dayun, 2008. Weed-suppression ability of Oryza longistaminata and Oryza sativa. Allelopathy J., 22: 345-352.
Direct Link - Hiroshi, N., 2008. Effects of husk extracts of wild rice spp. on seedling growth of lettuce, barnyard grass and Eclipta thermalis. Allelopathy J., 22: 391-396.
Direct Link - Hu, F., D. Wang, X.H. Chen and G.L. Ding, 2008. Allelopathic potential of rice accessions against barnyard grass in paddy field. Allelopathy J., 22: 379-384.
Direct Link - Inderjit, A. and K.M.M. Dakshini, 1994. Allelopathic effect of Pluchea lanceolata Asteraceae) on characteristics of four soils and tomato and mustard growth. Am. J. Bot., 81: 799-804.
Direct Link - Kato-Noguchi, H., 2002. Isolation of allelopathic substances in rice seedlings. Plant Prod. Sci., 5: 8-10.
Direct Link - Olofsdotter, M., D. Navarez and K. Moody, 1995. Allelopathic potential in rice (Oryza sativa L.) germplasm. Ann. Applied Biol., 127: 543-560.
CrossRef - Takeuchi, Y., S. Kawaguchi and K. Yoneyama, 2001. Inhibitory and promotive allelopathy in rice (Oryza sativa L.). Weed Biol. Manage., 1: 147-156.
CrossRef - Weidenhamer, J.D., T.C. Morton and J.T. Romeo, 1987. Solution volume and seed number: Often overlooked factors in allelopathic bioassays. J. Chem. Ecol., 13: 1481-1491.
CrossRef - Zhang, Z., Y. Zhou, Y. Lu, D. Li and L. Yu, 2005. Identification of allelopathic potential of Chinese rice (Oryza sativa L.) germplasm. Allelopathy J., 15: 111-118.
Direct Link