
A. Arghavani
Department of Agronomy and Plant Breeding, Faculty of Agriculture, University of Mohaghegh Ardabili, Ardabil, Iran
A. Asghari
Department of Agronomy and Plant Breeding, Faculty of Agriculture, University of Mohaghegh Ardabili, Ardabil, Iran
M. Shokrpour
Department of Agronomy and Plant Breeding, Faculty of Agriculture, University of Mohaghegh Ardabili, Ardabil, Iran
Mohammaddost Chmanabad
Department of Agronomy and Plant Breeding, Faculty of Agriculture, University of Mohaghegh Ardabili, Ardabil, Iran
Research Journal of Environmental Sciences
Year: 2010 | Volume: 4 | Issue: 1 | Page No.: 50-56







ABSTRACT
This study conducted to analyze genetic diversity in two Agropyron species, A. pectiniforme and A. elongatum, by RAPD analysis. Random Amplified Polymorphic DNA (RAPD) analysis using 12 primers produced 142 polymorphic bands with lengths ranging 564 to 2,000 bp. On the basis of Nei's gene index, the genetic diversity within ecotypes varied from 0.1014 to 0.178. The highest and lowest of this index were obtained in ecotype of 6951 (from A. pectiniforme; 0.178) and ecotype of 225 (from A. elongatum; 0.1014), respectively. The results showed high variation within ecotypes. The average gene diversity within (HS), total (HT) and coefficient of gene differentiation (GST) were 0.13, 0.27 and 0.47, respectively. Analysis of molecular variance (AMOVA) displayed significant variance within and among Agropyron ecotypes. The variance within ecotypes (61.08) was about two times higher than between ecotypes (31.24). Cluster analysis based on RAPD data using Nei's genetic distance categorizes the entries into four clusters. Using principle coordinate analysis, the first three coordinates accounted for the 52.84% of the total variation. Classifying the ecotypes by the two coordinates verified the results of cluster analysis.
PDF Abstract XML References Citation
How to cite this article
A. Arghavani, A. Asghari, M. Shokrpour and Mohammaddost Chmanabad, 2010. Genetic Diversity in Ecotypes of Two Agropyron Species using RAPD Markers. Research Journal of Environmental Sciences, 4: 50-56.
DOI: 10.3923/rjes.2010.50.56
URL: https://scialert.net/abstract/?doi=rjes.2010.50.56
DOI: 10.3923/rjes.2010.50.56
URL: https://scialert.net/abstract/?doi=rjes.2010.50.56
INTRODUCTION
Agropyron, as a range plant grows at the most of the rangeland of Iran. In world, Agropyron genus composed of 150 species, a 100 species existing over Asia. About 19 species of herbaceous and perennial plants have been found in North, Northwest and Central regions of Iran (Bor, 1970). The nearest relatives to Agropyron are Aegilops and Triticum (Refoufi et al., 2001) belonging to Triticeae family and can be crossed with each other. Today, plant geneticists are making many attempts to identify Agropyron species and transfer their useful genes for improving and developing new varieties for increasing disease resistance potential (Baden, 1991). Yellow dwarf disease resistance genes in Agropyron species have been introduced to wheat and barley (Bor, 1970). Three wheat lines has been crossed with A. elongatum and observed chromosome pairing and banding to produce a substitution disomic line, CI5321. The chromosome D1 of wheat is changed with chromosome 1 of Agropyron and become resistant to mosaic virus (Jiang et al., 1993). Also, there are many attempts to transfer the target genes from Agropyron particularly elongatum species to bread wheat (Liu et al., 2008, 2009; Cui et al., 2009).
Agropyron species have wide adaptation and grow in different climates. Therefore, gene pool conservation and its accurate application can be used in plant breeding programs in order to improvement rangeland and increasing forage production. Since, there is high variation within and among different species of Agropyron, so selection response for improving important traits is high and the gene pool can be more successfully used in breeding projects. In a breeding program, knowing of genome and ploidy level is very important. Therefore, such studies have been made on some Agropyron species (Assadi, 1995). Karyology of four Agropyron species has been investigated and tetraploid state with 7 chromosomes in genome base (2n = 4x = 28) has been demonstrated and differed from other attributes (Asghari et al., 2007).
Genetic distance based on genetic structure of biological population, may be stated by genotypic frequencies (genotypic distance) and or by different allele frequencies in loci (gene distance). Gene distance has positive association with heterosis (Falconer, 1996). Many researches have been made on measuring of genetic variation in Agropyron using protein and DNA markers (Che and Li, 2007; Refoufi and Esnault, 2008). Generally, determination of genetic variation is made by morphological, biochemical and molecular traits. Morphological traits are measured at field or greenhouse so they are cost consuming and are affected by environmental factors. Isozyme and storage proteins may be affected by environments, tissues and plant developmental stages (Kato and Yokoyama, 1992). Markers based on PCR e.g., RAPD is a useful way to overcome above difficulties (Virk et al., 1996). The RAPD markers are random and unlimited for number. There is no need to knowing of genome sequence to design RAPD primers. On the other hand, application of this markers are cost saving and their efficiency have been verified in genetic variation studies (Callow et al., 1997). Welsh et al. (1991) showed RAPD can be applied to study all organisms and is more valuable to identify various strains. RAPD analysis has been used widely to assess genetic variation in other plants (Huff, 1997; Garcia et al., 2002; Rajasekar et al., 2005; Narasimhan et al., 2006; Rout, 2006). In this study, RAPD analysis was applied to study genetic diversity among and within ecotypes of two Agropyron species.
MATERIALS AND METHODS
In this study, 10 various ecotypes from two species of Agropyron (A. pectiniforme and A. elongatum) were assessed during 2009 in molecular plant breeding laboratory of Agriculture Faculty, University of Mohaghegh Ardabili, Ardabil, Iran. Names and accession numbers of these ecotypes were shown in Table 1. These ecotypes were grown in greenhouse and leaf samples were harvested at rosette stage from seven individual plants of each ecotype for DNA extraction. The DNA of individual plants was extracted using the CTAB procedure according to Maroof et al. (1984). The quality and quantity of DNA samples were assessed using spectrophotometer (Techne, England) and 0.8% agarose gel electrophoresis. All of the DNA samples were diluted to 25 ng μL-1 and used in PCR reactions. Fifty RAPD primers (Metabione, Germany) were used to analyze polymorphism in the plants and polymorphic primers were used to genotyping. PCR reaction for RAPD analysis was performed in a volume of 15 μL contained 50 ng of DNA template, 2 mM MgCl2, 0.05 mM each dNTP, 0.132 μM primer, 1U Taq DNA polymerase and 1xPCR buffer. For RAPD primers the amplification profile consisted of a 5 min initial denaturation step at 94°C followed by 40 cycles of denaturing at 94°C for 30 sec, annealing at 37°C for 30 sec, extension step at 72°C for 2 min and a final extension step at 72°C for 10 min.
| Table 1: | Species, accession No. and origin of studied ecotypes of Agropyron |
 | |
The RAPD amplified products were analyzed using 1.5% agarose gels and ethidium bromide staining. Each polymorphic marker bands were scored as 1 (presence) and 0 (absence) and obtained data were analyzed using NTSYS pc 2.0, POPGENE 1.31 and Arlequin 3.11 software.
RESULTS AND DISCUSSION
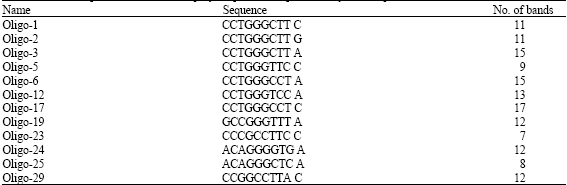
Results showed that from 50 studied primers, 12 primers were highly polymorphic and banding pattern resulted from these 12 primers were located in the intervals between 564-2000 bps and totally produced 142 polymorphic bands. The average band production for each polymorphic primer was 11.83. The name and sequence of the studied primers was outlined in Table 2. According to Table 2, the primers, Oligo-17 and Oligo-23, produced highest and lowest number of polymorphic bands, respectively. Also, ecotypes of 224 (A. elongatum) and ecotype of 225 (A. elongatum) had most and least amplified bands, respectively (not shown data). On the basis of Nei's gene index (Nei and Chakravarti, 1977), the genetic diversity within ecotypes varied from 0.1014 to 0.178. The highest and lowest of this index were obtained in ecotypes of 6951 (from A. pectiniforme; 0.178) and 3202 (from A. elongatum; 0.1014), respectively. Gene diversity in other studied ecotypes was in average level. The results showed high diversity within ecotypes. The amount of gene diversity of ecotypes was shown in Table 1. The estimated average gene diversity within (HS), total (HT) and coefficient of gene differentiation (GST) were 0.13, 0.27 and 0.47, respectively.
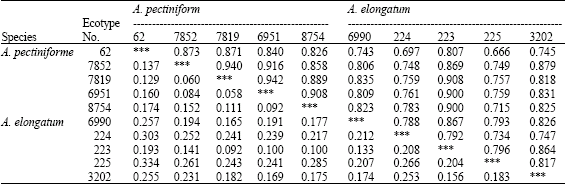
Cluster analysis based on RAPD data using Nei (1972) genetic distance (Table 3) categorized the entries into 4 clusters (Fig. 1). First and second groups composed of 6 and 2 ecotypes, respectively and the remained entries located on two different clusters. The first cluster encompassed the ecotypes belonged to A. pectiniforme together with one ecotype of A. elongatum and second cluster included two ecotypes of A. elongatum.
| Table 2: | Name, sequence and number of polymorphic bands produced by studied primers |
 | |
| Table 3: | Coefficient of similarity (above diagonal) and distance (below diagonal) between studied ecotypes based on Nei (1972) genetic distance method |
 | |
| ***p = 0.01 | |
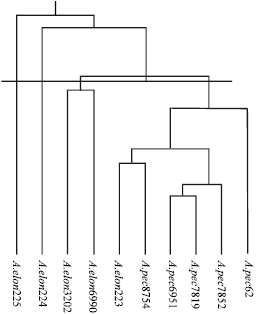 | |
| Fig. 1: | Dendrogram obtained from cluster analysis using UPGMA method based on Nei’s (1972) genetic distance |
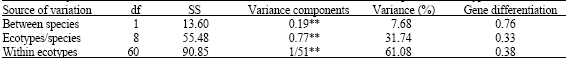
| Table 4: | Analysis of molecular variance (AMOVA) based of RAPD data for studied species and ecotypes |
 | |
| **p = 0.01 | |
Other ecotypes of A. elongatum located in two different clusters. In order to study of primer efficiency that used in grouping, each primer was separately analyzed. Primers of Oligo-1 and Oligo-29 were more efficient than others to distinguish ecotypes (Fig. 3, 4). These may be applied to distinguish the two species and ecotypes.
Analysis of molecular variance (Table 4) displayed significant variance within and among Agropyron ecotypes. The variance within ecotypes (61.08%) was about two times of between ecotypes (31.24) and only 7.68% of totally estimated variance belonged to between species. According to these results it was concluded that some of studied ecotypes may be presented as potential gene resources for using in further breeding aims. Separately analysis of variance for each of the two species showed significant differences between and within ecotypes. Estimated variance between and within ecotypes of A. elongatum were 47.07 and 52.93% with coefficient of gene differentiation of 0.47. Also, it was resulted that within ecotype variance of A. pectiniforme was higher than A. elongatum and the estimated average gene diversity between and within ecotypes and coefficient of gene differentiation were 30.45, 69.55 and 0.30%, respectively.
The estimated genetic distance of 10 ecotypes based on Nei (1972) genetic distance were ranged between 0.060 to 0.334% and the highest estimated values belonged to A. pectiniforme (ecotype No. 62) and A. elongatum (ecotype No. 225) (Table 3). Because of higher genetic difference between A. elongatum 225 and A. pectiniforme 62, it was recommended to use these ecotypes in order to make crossing for design further breeding efforts and benefiting heterosis in hybridization.
Using Principle Coordinate Analysis (PCoA), the first three coordinates accounted for the 52.84% of the total variation. First, second and third components determined in turn 29.28, 19.28 and 14.28% of total variance and it was concluded that the adequate coverage of selected primers and suitable sampling were existed (Fig. 2). Therefore, the grouping based on the first two components confirmed cluster analysis results.
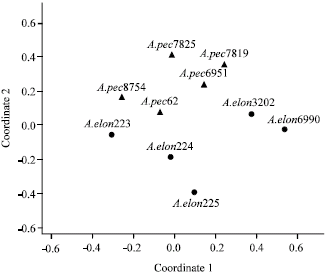 | |
| Fig. 2: | Grouping of studied ecotypes using the first and second coordinates |
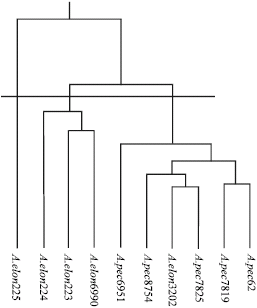 | |
| Fig. 3: | Dendrogram obtained from cluster analysis using UPGMA method based on Oligo-1 markers data |
This study suggested RAPDs is a good technique to analyze Agropyron diversity in respect to its time and cost saving. Nevertheless, the most important disadvantage of RAPD is dominant nature of the amplified markers. The results displayed high genetic variability in Agropyron. Analysis of molecular variance (AMOVA) indicated that high levels of variation existed in within ecotypes. This result with low gene differentiation is expected in cross-pollinated species (Hamrick and Godt, 1996). Several reports about genetic analysis in grasses using molecular markers showed variation within was greater than between population (Che and Li, 2007; Refoufi and Esnault, 2008). Since, the half of the ecotypes in this study was open pollinated, the obtained results were expected. Separately analyses of molecular variance for the two species showed elongatum species, a self and some cross pollinated, had same variation of within and among ecotypes. While in pectiniforme species, a cross pollinated, variance of within was about twice of between. These are corresponded to expected genetic structure of self and cross pollinated species, respectively.
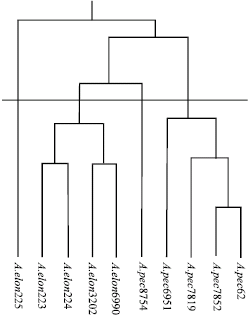 | |
| Fig. 4: | Dendrogram obtained from cluster analysis using UPGMA method based on Oligo-29 (b) markers data |
The UPGMA clustering of Agropyron ecotypes and PCoA established by RAPDs, had not direct relationships with geographical pattern. Moreover, some genotypes from the two species were clustered together. Since, the two species have close relativeness, RAPDS could unravel genetic similarity at noncoding regions of genome. That is two species may be different morphologically but have too similarities at genome level. RAPDS proved to be a suitable technique to analyze genetic variation between and within species in Agropyron.
In conclusion, high genetic variability found within Agropyron ecotypes, specialy in Pectiniforme species, which is a cross pollinated species. This variability can be used in plant breeding programs, in order to improvement of range plants and increasing of forage production. Also, the ecotypes of each species with high genetic distance can be used in crossing programs in order to producing mapping populations and utilization of heterosis.
REFERENCES
- Asghari, A., Y. Agayev and S.A.A. Fathi, 2007. Karyological study of four species of wheat grass (Agropyron sp.). Pak. J. Biol. Sci., 10: 1093-1097.
CrossRefPubMedDirect Link - Assadi, M., 1995. Meiotic configuration and chromosome number in some Iranian species of Elymus L. and Agropyron Gartner. Bot. J. Linnean Soc. London, 117: 159-168.
Direct Link - Baden, C., 1991. A taxonomic revision of Psathyrostachys (Poaceae). Nordic J. Bot., 11: 3-26.
Direct Link - Che, Y.H. and L.H. Li, 2007. Genetic diversity of prolamines in Agropyron mongolicum Keng indigenous to Northen China. Genet. Res. Crop Evol., 54: 1145-1151.
CrossRefDirect Link - Cui, H., Z. Yu, J. Deng, X. Gao, Y. Sun and G. Xia, 2009. Introgression of bread wheat chromatin into tall wheatgrass via somatic hybridization. Planta, 229: 323-330.
CrossRefDirect Link - Falconer, D.S. and T.F.C. Mackay, 1996. Introduction to Quantitative Genetics. 4th Edn., Prentice Hall, Harlow, England, ISBN-13: 9780582243026, Pages: 464.
Direct Link - Garcia, P., J.V. Monte, C. Casanova and C. Soler, 2002. Genetic similarities among Spanisch populations of Agropyron, Elymus and Thinopyrum, using PCR-based markers. Genet. Res. Crop Evol., 49: 103-109.
Direct Link - Hamrick, J.L. and M.J.W. Godt, 1996. Effects of life history traits on genetic diversity in plant species. Philos. Trans. R. Soc. London B: Biol. Sci., 351: 1291-1298.
CrossRefDirect Link - Huff, D.R., 1997. RAPD characterization of heterogeneous perennial ryegrass cultivars. Crop Sci., 37: 557-564.
CrossRefDirect Link - Jiang, L., B. Friebe, H. Dhaliwal, Y.J. Martin and B.S. Gill, 1993. Molecular cytogenetic analysis of Agropyron elongatum chromatin in wheat germplasm specifying resistance to wheat streak virus. Theor. Applied Genet., 5: 89-111.
Direct Link - Kato, K. and M. Yokoyama, 1992. Geographical variation in heading characters among wheat landraces, Triticum aestivum L. and its implication for their adaptability. Theor. Applied Genet., 84: 259-265.
Direct Link - Liu, H., X. Gao and G. Xia, 2008. Characterizing HMW-GS alleles of decaploid Agropyron elongatum in relation to evolution and wheat breeding. Theor. Applied Genet., 116: 325-334.
Direct Link - Liu, H., S.H. Liu and G. Xia, 2009. Generation of high frequency of novel alleles of the high molecular weight glutenin in somatic hybridization between bread wheat and tall wheatgrass. Theor. Applied Genet., 118: 1193-1198.
Direct Link - Narasimhan, S., P. Padmesh and G.M. Nair, 2006. Assessment of genetic diversity in Coscinium fenestratum. Biol. Plant., 50: 111-113.
Direct Link - Nei, M. and A. Chakravarti, 1977. Drift variances of FST and GST statistics obtained from a finite number of isolated populations. Theor. Popul. Biol., 11: 307-325.
PubMed - Rajasekar, S., S.H. Fei and E.C. Nick, 2005. Analysis of genetic diversity in rough bluegrass determined by RAPD markers. Crop Sci., 46: 162-167.
CrossRef - Refoufi, A. and M.A. Esnault, 2008. Population genetic diversity in the polyploid complex wheatgrass using isoenzym and RAPD data. Biol. Plant., 52: 543-547.
CrossRefDirect Link - Refoufi, A., J. Jahier and M.A. Esnault, 2001. Genome analysis of a natural hybrid with 2n=63 chromosomes in the genus Elytrigia Desv. (Poaceae) using the GISH technique. Plant Biol., 3: 386-390.
Direct Link - Rout, G.R., 2006. Evaluation of genetic relationship in Typhonium species though random amplified polymorphic DNA markers. Biol. Plant., 50: 127-130.
CrossRef - Saghai-Maroof, M.A., K.M. Soliman, R.A. Jorgensen and R.W. Allard, 1984. Ribosomal DNA spacer-length polymorphisms in barly: Mendelian inheritance, chromosomal location, and population dynamics. Proc. Natl. Acad. Sci., 81: 8014-8018.
CrossRefDirect Link - Virk, D.S., B.V. Ford-Lyod, M.T. Jackson, H.S. Pooni, T.P. Clemeno and H.J. Newbury, 1996. Prediction of quantitative variation within rice germplasm using molecular markers. Heredity, 76: 296-304.
Direct Link - Welsh, J., C. Peterson and M. McClelland, 1991. Polymorphisms generated by arbitrarily primed PCR in the mouse application to strain identification and genetic mapping. Nucleic Acids Res., 19: 303-306.
Direct Link