
M. Faravani
Institute of Biological Sciences, University of Malaya, 509603, Kuala Lumpur, Malaysia
B.B. Baki
Institute of Biological Sciences, University of Malaya, 509603, Kuala Lumpur, Malaysia
Shogo Kato
Institute of Statistical Mathematics, 4-6-7
Minami-Azabu, Minato-Ku, Tokyo 106-8569, Japan
K. Shimizu
Department of Mathematics, Keio University, 3-14-1 Hiyoshi, Kohoku-ku,Yokohama 223-8522, Japan
C.H. Sim
Institute of Mathematical Sciences, University of Malaya, 509603, Kuala Lumpur, Malaysia
Research Journal of Environmental Sciences
Year: 2009 | Volume: 3 | Issue: 4 | Page No.: 427-438







ABSTRACT
The ensuing plant branching patterns were studied at three planting densities 1, 2 and 3 plants per wooden box, raised until maturity. Circular statistics were used to represent the evolving spatial distribution of branches along density gradients of the translation with certain assumptions. The position of each branch was characterized by three parameters: horizontal rotation (φ), vertical rotation (θ) and translation. The values of φ were measured within 45° angle using a circular protractor, divided into 8 angular sectors oriented clockwise, while φ was measured from the geographical north direction (0°). The translation was registered from 1 to 5 categories. The distribution φ was uniform, but the distribution θ was non uniform. The former was distributed symmetrically in plants at densities 1, 2 and 3 with respective mean direction values of 52.3°, 47.8° and 41.8°. The mean direction of the angle θ was different between plants at different densities and translations, whereby branches became progressively more erect with increasing density regimes from 1 to 3 and from the base to the top of the canopy. The registered mean direction of each translation from 1 up to 5 from the base were 60.2°, 49.7°, 40.1°, 30.5° and 22.5°, respectively.
PDF Abstract XML References Citation
How to cite this article
M. Faravani, B.B. Baki, Shogo Kato, K. Shimizu and C.H. Sim, 2009. Effects of Intra-Plant Competition on the Ensuing Spatial Branching Patterns of Straits Rhododenron. Research Journal of Environmental Sciences, 3: 427-438.
DOI: 10.3923/rjes.2009.427.438
URL: https://scialert.net/abstract/?doi=rjes.2009.427.438
DOI: 10.3923/rjes.2009.427.438
URL: https://scialert.net/abstract/?doi=rjes.2009.427.438
INTRODUCTION
Melastoma malabathricum L., otherwise known locally as the Straits Rhododenron, Indian Rhododenron or Singaporean Rhododenron, is a common pioneer shrub species in arable lands, abandoned farmlands, secondary forest openings and derelict areas in Malaysia and elsewhere, especially in tropical and sub-tropical forests of India, Southeast Asia and Australia (Meyer, 2001; Baki, 2004). The species is a common scourge in young rubber, oil palm, coconut plantations and fruit orchards and open places in Malaysia.
Spatial analysis of tree trunks and development of biological branching structures has become an established method to infer tree population dynamics such as self-thinning or gap recruitment in forest communities (Caplat et al., 2008; Getzin and Wiegand, 2007). Inter- and intra-plant competition models illustrating density-mediated effects on yield and biomass in plants have been the subject of vigorous academic pursuits by several workers (Kropff and Struik, 2002; Cornelissen and Stiling, 2008). Surveys on asymmetric growth are very useful in plant competition and are essential in practical management for improving timber quality and stand leaf area index or wind resistance of stands (Rudnicki et al., 2001; Rock et al., 2004; Getzin and Wiegand, 2007). Under field conditions without water stress, branches do not converge into a universal movement to a favoured mature azimuth. On the contrary, there is a trend toward azimuthal dispersion. Azimuthal movement of branches is sensitive to initial clumping of foliage, being triggered initially and more importantly, when the initial clumping is strong (Chelle, 2006; Saudreau et al., 2007). Some plant species have a random distribution of horizontal rotation angle φ of leaf, independent of planting pattern while other plant species can modify leaf orientation according to the conditions. Plant canopy behaviour has a significant effect on light interception (Maddonnia et al., 2001). Drouet and Moulia (1997) reported that uniform distribution of leaf azimuth was effected by light interception and crop architecture. Each leaf was characterized by three parameters: leaf horizontal rotation angle, leaf vertical rotation angle and leaf height (translation). The horizontal rotation angle has received much less attention.
Studies on the leaf angle (θ, φ) have shown that the effects of initial plant orientation on θ of leaf and height were less spectacular. Initial plant orientation and density had a small effect on leaf vertical rotation angle. An effect of density was observed only on leaf translation, but no intercalation of leaves between adjacent plants was noticed in development of maize (Girardin and Tollenaar, 1994; Maddonnia et al., 2001). There is a remarkable correlation between plant architecture and the bio-geographic distribution of species in different places and this correlation can be used to assess plant distribution in ecological science. Other reports indicated that a species may show different orientations according to the environment in which it grows and its behaviour is relatively fixed at the species level (Ezcurra et al., 1991).
In this study, the system of primary and secondary branches of M. malabathricum as a function of translation (plant height), their horizontal rotation angles, vertical rotation angle and translation of branches over different densities were checked for possible adaptive re-positioning of successive branches during 161 days of their growth from the transplanting date. The spatial scales for branching patterns were analyzed along plant height, as well as the directional preferences of growing branches in relation to different plant densities and light regimes at the stand level. The hypothesis was that branch orientation is a selected trait that permits changes in the vertical and horizontal angle to achieve adequate light interception.
MATERIALS AND METHODS
One hundred young uniform seedlings of M. malabathricum of the same cohorts were collected in May 2006 from the campus of the University of Malaya, Kuala Lumpur (3° 8’ N; 101° 42’ E) Malaysia and raised in wooden boxes previously filled with garden soil of Malacca series in an insect-proof house with 12 h of natural sunlight (mean midday radiation of 622 and 125 μmole photon/m2/sec outdoor and inside in insect-proof house respectively) and mean ambient temperatures of 33 ± 2 °C (day) and 25 ± 2 °C (night) at Rimba Ilmu, University of Malaya. The most uniform plants were selected randomly and transplanted into wooden boxes, each measuring 72x72x30 cm at the density of 1(D1), 2(D2) and 3(D3) plants box-1, each with 3 replications The plants were transplanted into the centre of the box for the single plants, while for 2 or 3 plants per box the plants were spaced equidistant at 10 cm from each other. The plants were watered once daily, in the morning from above with a fine rose. The boxes were arranged in a completely randomized design.
For the branching pattern studies, each plant was progressively divided into 50 cm heights from the soil surface and denoted accordingly. A circle of transparent plastic sheet divided into 8 sections, each with a 45° degree angle, was prepared (Girardin and Tollenaar, 1994; Drouet and Moulia, 1997). Overlays were prepared to assess branching patterns of M. malabathricum with this plastic sheet. The direction and branch angles (primary branches with respect to the mother plant) in each section were recorded with the aid of a compass oriented clockwise. The vertical rotation angles were measured with a clinometer.
A botanical numbering system were applied to plant branches, counting the branch order(s) outward from the main stem (Borchert and Slade, 1981).The mean angles of orientation of primary branches with respect to the main stem and other branches were measured, so were the angles of secondary branches with respect to primaries (Fig. 1), viz. the horizontal rotation angle (φ), the vertical rotation angle (θ), translation (H) for each 50 cm throughout the plant height, based on the main stem for each translation of 50, 100, 150, 200 and 250 cm (hereinafter referred as translation 1, 2, 3, 4 and 5, respectively). The horizontal rotation angle of the branch was measured within sections the 45 degree, using a home-made circular protractor, with clockwise orientation from the north direction (0°).
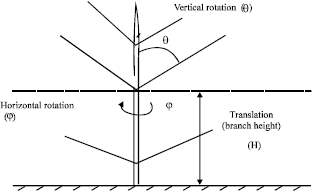 | |
| Fig. 1: | The three parameters of the horizontal rotation angle φ, the vertical rotation angle θ and translation (H) measured from the north direction during plant growth to describe the development of the aerial structure of Melastoma malabathricum L. |
The branch-base height above the soil was measured using a rule tape. Growth parameters, viz., plant height; lengths of primary and secondary branches and reproductive traits (time and duration of flowering, number of flowers branch-1 or flowers plant-1) were recorded.
Data Analysis
Directional data commonly occur in environmental studies when measurements are taken from an orientation or cyclical timing application. The mean direction is denoted by mean vector values, defined as follows: Let θ1 ... θn be the angular observations expressed in the form of radians [0, 2θ]. Then, the mean direction, 6, is defined by:
(1) |
where, i is the imaginary unit, i.e., i2 = -1
Two frequently-used families of distributions for circular data include the von Mises and the Uniform distribution (Fisher, 1993; Gatto and Jammalamadaka, 2007).
The sample mean direction is a common choice for moderately large samples, because when combined with a measure of sample dispersion, it acts as a summary of the data suitable for comparison and amalgamation with other such information. The sample mean is obtained by treating the data as vectors of length as one unit and using the direction of their resultant vector r that it lies in the range (0, 1). Resultant vector r is a measure of concentrations 1-r is a measure of dispersion. Lack of dispersion 1-r = 0 and maximum dispersion is 1-r = 0. The angular variance is defined as s2 = 2(1-r). Note that r = 0 does not necessarily indicate a uniform distribution (Zar, 1998). The Rayleigh’s test is used to test whether the population from which the sample is drawn differs significantly from randomness (Fisher, 1993; Zar, 1998; Mardia and Jupp, 2000). In order to compare and test whether the mean directions of two or more samples will differ significantly from each other, a widely used statistical test is the Watson-Williams test or approximate Analysis of Variance (ANOVA) (Batschelet, 1981; Mardia and Jupp, 2000; Jammalamadaka and Sengupta, 2001).
The hypothesis was that there is a correlation between a circular variable (horizontal or vertical rotation angle) and linear variable (branch length or translation). The hypothesis entailed the calculation of the circular-linear correlation coefficient (Fisher, 1993; Zar, 1998; Mardia and Jupp, 2000; Jammalamadaka and Sengupta, 2001). This correlation coefficient r ranges from 0 to 1. Suppose that a linear-circular correlation (Mardia and Jupp, 2000), which is a measure of correlation between a linear variable x and an angular variable θ, is defined by:
(2) |
Where:
| rxc | = | Corr (x, cosθ) |
| rxs | = | Corr (x, sinθ) |
| rcs | = | Corr (cosθ, sinθ) |
The hypothesis of no circular-linear association is rejected if r2 is too large. The data were processed and displayed with the software ORIANA and the R Project for statistical computing of the circular data, median, circular mean and concentration parameter.
RESULTS AND DISCUSSION
Melastoma malabathricum has terminal branch flowers and after flowering, commonly old branches at the base and inside of canopy die. This plant displays both terminal- and axillary-flowers. The branching pattern of M. malabathricum illustrates that of Leeuwenberg’s model. This model consists of equivalent orthotropic modules, each of which is determined in its growth by virtue of ultimate production of a terminal inflorescence. Branching is three-dimensional, producing the several equivalent modules and is correlated with flowering except in a few examples with branched sterile juvenile axes (Halle et al., 1978; Judd, 1986).
Horizontal Rotation of Plant Branches
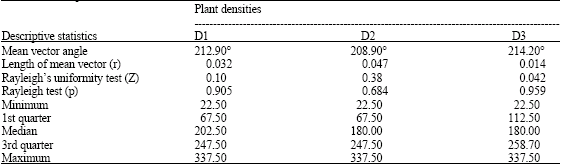
The Rayleigh’s test showed the distribution of φ was centrally symmetrical in different plant populations and the computed lengths of mean vector (r) were 0.032, 0.047 and 0.014 for the density regimes of D1, D2 and D3, respectively (Table 1). Figure 2 shows box-plots of (φ) for each density of the branches.
Such association is also given in Table 1, which is a summary of the horizontal rotation angle for each of the plant densities. The angular observations were calculated by Eq. 1. Figure 2 seems to imply that (φ) values are not dependent on, or influenced by, the density of branches. In this regard, we applied the circular ANOVA to test if density influences the mean direction of (φ).
| Table 1: | Directions of symmetric growth for the horizontal rotation angle of Melastoma malabathricum as analyzed by descriptive statistics |
 | |
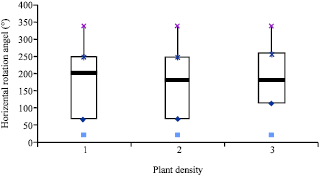 | |
| Fig. 2: | Box-plots of the horizontal rotation angles of branches (φ) as a function of planting density in Melastoma malabathricum |
| Table 2: | Approximate ANOVA values for testing the influence of plant density of Melastoma malabathricum on horizontal rotation angle of branches (φ) and the hypothesis: (H0: v1 = v2 = v3 against H1: v1≠ v2 or v1 ≠ v3 or v2 ≠ v3) |
 | |
| ns: No significant difference at p < 0.0.5 | |
The results of the circular ANOVA test applied to the horizontal rotation angles are shown in Table 2. With F-value of 0.0203 and because p (F>0.0203) ≈ 0.980>0.05, the null hypothesis is accepted. This implies that the mean directions for all three densities are not different from each other for each planting density of M. malabathricum.
Similar patterns of symmetrical distributions were registered in the different translations, irrespective of density regimes to which the plants of M. malabathricum were subjected. The registered horizontal rotation angle (mean direction) for the translations 1-5 were 249.292°, 175.935°, 230.576°, 75.832° and 90°, respectively. The means of algebraic values of the horizontal rotation angles between two successive branches were not significantly different, irrespective of the plant density, with a mean vector of μφ = 212.890 after 161 days of growth from seedling time to the successive date of measurement.
In addition, the observations do not seem to follow the uni-modal distribution (Fig. 3). Therefore, any given one branch had an equal probability of the alternate position (Drouet and Moulia, 1997). Similar observations were recorded for Larrea ameghinoi (Ezcurra et al., 1991):
Influence of Plant Density on the Vertical Rotation Angle
It is too natural that the vertical rotation angle in the range from 0 to 180 degrees and it was also observed in the Rayleigh’s uniformity test that the vertical rotation angles were not distributed uniformly in the different plant densities (Table 3). The ANOVA applied to assess the effects of density on the length of branches of M. malabathricum. Since P (F>12.702) = 5.80x10-5 < 0.05, the hypothesis was rejected that the mean directions for all densities are considered the same (Table 4). Given these results, a natural question is: “AWhen subjected to plant density, which mean direction of the branches will take a different value from the others θ. Here, the comparisons for three pairs of densities were made, viz., Densities 1 and 2, Densities 1 and 3 and Densities 2 and 3. Table 5 shows approximate ANOVA table values for Densities 1 and 2, Densities 1 and 3 and Densities 2 and 3. From Table 5, it can be seen that the F-value is 3.68 in Densities 1 and 2 and here, the null hypothesis was accepted, i.e., μ1 = μ2, since P (F>3.68) ≈ 0.058 > 0.05. On the other hand, F-values are 26.97 and 10.12 Table 5 in Densities 1 and 3 and Denesties 2 and 3; because of this, the null hypotheses can be rejected, μ1 = μ3 and μ2 = μ3, respectively. The mean direction is defined in the same manner, as shown in Equation 1. Invariably, there is a tendency that the angles for Density 3 generally take smaller values than those for Densities 1 and 2. As for the comparison between Density 1 and 2, it seems that Fig. 4 and Table 3 show that distributions of the angles for Density 1 and 2 are similar. However, from Table 5, the mean direction of the angles for Density 2 is less than that for Density 1.
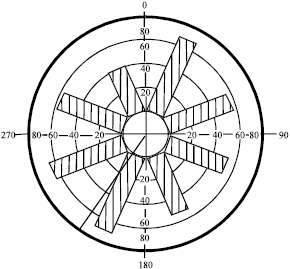 | |
| Fig. 3: | The mean vector (μ = 212.89°) of the horizontal rotation angle φ irrespective of plant density in the single plant spacing of Melastoma malabathricum |
| Table 3: | Descriptive statistics of the vertical angle θ of branches in Melastoma malabathricum |
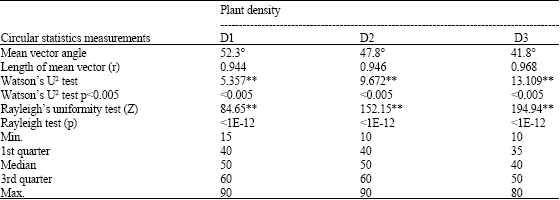 | |
| *Significant difference at p < 0.05 | |
| Table 4: | Approximate ANOVA table for testing the influence of density on the rotation angle of branches of Melastoma malabathricum |
 | |
| *Significant difference at p < 0.05 | |
When plants of M. malabathricum were subjected to density stress, the ensuing competition between branches led them to be more erect in stature as in D3, with measurably smaller vertical rotation angle θ values.
The same result was observed in that the branches of density-stressed plants had an inward curving of the branch planes (Ezcurra et al., 1991). When plant population increased, branches became progressively more erect with the mean angles decreasing from 52.27° to 47.8° and 41.81° (Table 4). Results were in agreement with other reports citing significantly smaller θ angles at higher densities.
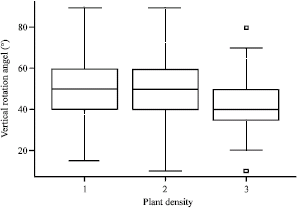 | |
| Fig. 4: | Box-plots of the vertical rotation angle of the branches for each plant density of Melastoma malabathricum |
| Table 5: | Approximate ANOVA for (a) Densities 1 and 2, (b) Densities 1 and 3 and (c) Densities 2 and 3 of Melastoma malabathricum |
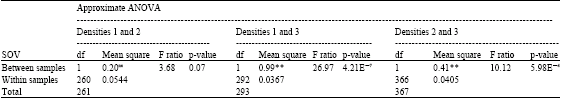 | |
| **Significantly difference at p < 0.01; ns, not significantly difference at p < 0.05 | |
Relationship among Vertical Rotation Angles and Translations
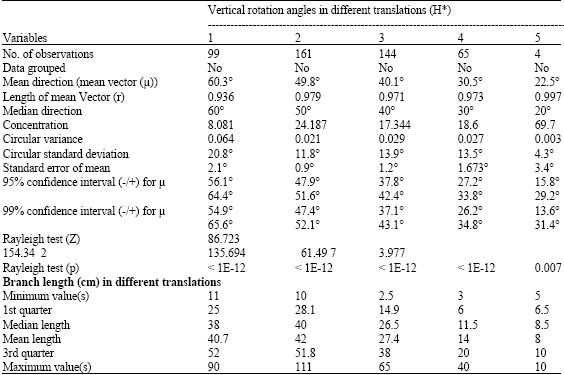
Statistical analysis of the circular data indicated that the lengths of mean vector increased throughout the translation from the base to the top of plant height in other words ,branches were more erect in the top of the plant canopy, arguably for optimization of light interception (Table 6).
The Watson-Williams test with multiple comparison showed there were significant differences in F = 50.157, (p < 0.01) for the vertical rotation angle in different translations. When Watson-Williams tests for paired comparisons of the vertical rotation angles for branches at different heights were done, results indicated that θ values decreased with increasing translations from 0 cm to 150 cm, with non-significant differences for ensuing translations beyond 150 cm (Table 6). However, if the angles were greater than 50°, it was apparent that branch lengths decrease slightly as the angle increased. These results are consistent with those obtained by others (Drouet and Moulia, 1997; Maddonnia et al., 2001), who studied the effect on developmental plasticity in crown asymmetry both by the directionality of solar radiation geometry and by local competition between individual plants and their modules (Ezcurra et al., 1991).
One report showed that the vertical and horizontal rotation of foliage clusters are non-random as are secondary leaf and branch orientations, which allow branches and leaves to intercept light without interfering with each other (Neufeld et al., 1988).
The difference in vertical rotation angles at different translations can easily be compared by plotting the lengths and translations of multiple box-plots. Figure 5 clearly shows the prevailing inverse relationship between translation and θ values in M. malabathricum in which the vertical rotation angle decreases when plant height increases. This conclusion is strengthened by the Watson-Williams F-tests for vertical rotation angles in different translations of M. malabathricum (Table 7).
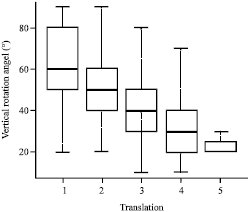 | |
| Fig. 5: | Box-plots of the vertical rotation angles (degree) for Melastoma malabathricum against the different translations: 1, 0-50 cm; 2, 50-100 cm; 3, 50-100 cm; 4, 150-200 cm; 5, 200-250 cm |
| Table 6: | Summary of vertical rotation angles and branch length at different translations of Melastoma malabathricum |
 | |
| *1, 0-50 cm; 2, 50-100 cm; 3, 100-150 cm; 4, 150-200 cm; 5, 200-250 cm | |
| Table 7: | Pair-wise comparison of vertical rotation angles at different translations of Melastoma malabathricum by Watson-Williams F-tests, where F and P are Fisher’s variance ratio and the probability associated with the null hypothesis, respectively that (H0: v1 = v2 against H1: v1θ v2) |
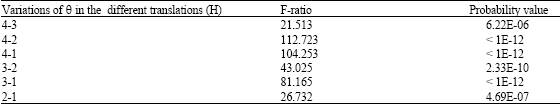 | |
| There is no comparison for translation 5 because this included less than 5 observations | |
| Table 8: | Summary of the Scheffe’s test on length of branches for each density of Melastoma malabathricum* |
 | |
| *Values with the same letter(s) have no significant difference at α = 0.05 | |
This is a suitable strategy for plants of M. malabathricum to achieve enough light for ensuring growth of leaves growing down inside the canopy, where self-shading leads to lower light availability than at the top of the plant canopy.
Table 6 Shows the relationship between translation and length of branches, this reveals that, as the height of branches increases, branch length decreases, with the exception that the median of translation 2 (50-100 cm) is slightly greater than that of translation 1 (0-50 cm). Formal comparisons with Scheffe’s test indicated that the lengths decreased significantly which had F = 44.43 and α < 0.01 from 41 to 8 cm at translations 1 and 4 respectively, but the difference in branch lengths between the translations 1 and 2 was not significant at α < 0.05 and no comparison was made for translation 5 because it included less than 5 observations for Scheffe’s test (Table 8). Table 8 summarizes branch lengths at each translation. The box-plots listed in Fig. 6 show that branch lengths of M. malabathricum have an inverse relationship with the plant translation. Branches in the top of the canopy were more erect than their counterparts at lower heights, arguably for optimization of light interception. Such orientation and distribution of branches allow the gradual warming of leaf surfaces during the morning, with maximum light interception (Ezcurra et al., 1991).
Evaluating the Influence of Plant Density on Branch Length
ANOVA for single-factor plant density regimes on the branch lengths indicated that with increasing plant density, branch length significantly decreased (p < 0.05) from D1: 40.4 cm, D2: 30.3 to D3:28.6, although there was no significant difference between the length of branches in D2 and D3. The lengths of branches for each density, explicitly showing that the higher the planting density, the shorter was the length of branches of M. malabathricum. It is possible to confirm this interpretation based on Scheffe’s test on length of branches for each density of Melastoma malabathricum (Table 8).
Circular-Linear Correlation among Branch Lengths with Vertical Rotation Angle
A natural question to address is whether there is a definite relationship between the lengths and vertical rotation angles of branches. Table 8 shows a plot of the vertical rotation angles θ and lengths of branches. Possible relationships between vertical rotation angle, circular variable and branch length and linear variable in M. malabathricum, as influenced by plant density regimes were attempted by using equation 2. The linear-circular correlation between the lengths and θ angles is given by r2 = 3.0x10-3. The highest circular-linear correlation (r = 0.594, p < 0.01) between the vertical rotation angle values and the lengths of branches in different plant densities was observed in the translations exceeding 150 cm of the plant height.
All measures in this study were taken during a growth period of 161 days from seedling to flower-displaying growth stages and the results showed that the vertical rotation angles of the branches bear an inverse relationship to the translations. Generally, the higher the translation of the canopy, the shorter branches were in length. Vertical rotation angles were proportional to branch lengths when the θ values did not exceed 50°. If θ is in excess of 50°, the branch length decreases slightly as θ increases. However, a circular-linear correlation coefficient shows that there is no clear association between vertical rotation angle θ and branch lengths. Plant density has a negative effect on the length of branches, but horizontal rotation angles φ were dependent on density to which the plants of M. malabathricum were subjected.
Light absorption by plants affects the development of a canopy directly, through photo-biological processes. Asymmetric tree growth is an adaptation to maximize photosynthesis by growing in response to the presence of gaps and neighbours, topographical site conditions or incoming solar radiation. High neighbourhood densities may result in density-dependent mortality of individuals or modules, or/and this may be compensated by shifting the crown centers away from the main stem as the tree expands branches on the side of canopy gaps in the lower translations, or closer or erect to the main stem in the higher translations. It follows that with higher plant densities, or with the measurement extended for a longer period, a long-term competitive effect among neighbouring plants may prevail. If this happens, asymmetric distribution of the horizontal rotation angle to the gap direction of the canopy as branch networks develop through plastic (Harper, 1977) response to a heterogeneous light environment because canopy structure is mainly subjected to maximize photosynthesis. It is generally assumed that trees optimize light harvest via more regular crown patterns; also, this is important in natural systems because the range of influence of plants becomes greater with size and age. Therefore, larger plants beyond immediate neighbours often have the greatest influence on the growth of a focal plant (Muth and Bazzaz, 2002; Getzin et al., 2006). Further plant modules tend to expand their growth toward the gaps prevailing in the canopy.
Results demonstrate that competitive interactions with neighbours affect the spatial arrangement of branch systems in M. malabathricum. Branch systems were more developed away from the maximum competitive pressure of neighbors, as asymmetry was not observed in crown shape. Density of neighbors did not appear to contribute significantly to the relation between branch system and pressure from neighbors for 161 days after transplanting the seedling into wooden boxes. The resulting spatial pattern tended to reduce the overlap between neighboring branch systems. A plant with a close neighbor responds by investing in branch growth away from the competitive pressure or simply into zones free of neighbors (Brisson and Reynolds, 1994). Under this model, two plants can be close to each other and not compete. Competition in the population is for space and only occurs when a plant branch system is crowded on all sides.
It would be interesting to see the photosynthetic efficiency of branches subjected to different light regimes within the translations and relate branch growth and dispersion into the gaps or away from the competition zones. It would be equally interesting to assess the azimuthal positioning of branches in relation to their photosynthetic efficiency, or whether uniformity in branch alignment and symmetry within a canopy are actually translated into photosynthetic efficiency or biomass production among competing plants.
ACKNOWLEDGMENTS
We acknowledge the financial assistance through Fundamental Grant No. PS074/2007B given to the first author by the University of Malaya and to the Institute of Biological Sciences, University of Malaya for providing the facilities throughout this study. The authors appreciate the help rendered by Prof. Dr. Bill Gregg of University of State Mississippi:, USA and two other anonymous referees for their review and useful suggestions on the manuscripts.
REFERENCES
- Baki, H.B., 2004. Invasive weed species in Malaysian agro-acosystems: Apecies, impacts and management. Malaysian J. Sci., 23: 1-42.
Direct Link - Zar, J.H., 1998. Biostatistical Analysis. 4th Edn., Prentice Hall, New Jersey, ISBN-13: 978-0130815422.
Direct Link - Saudreau, M., H. Sinoquet, O. Santin, A. Marquier and B. Adam et al., 2007. A 3-D model for simulating the spatial and temporal distribution of temperature within ellipsoidal fruit. J. Agric. For. Meteorol., 147: 1-15.
CrossRefDirect Link - Rudnicki, M., U. Silins, V.J. Lieffers and G. Josi, 2001. Measure of simultaneous tree sways and estimation of crown interactions among a group of trees. Trees, 15: 83-90.
CrossRefDirect Link - Rock, J., K.J. Puettmann, H.A. Gockel and A. Schulte, 2004. Spatial aspects of the influence of silver birch (Betula pendula L.) on growth and quality of young oaks (Quercus spp.) in central Germany. J. For., 77: 235-247.
Direct Link - Borchert, R. and N.A. Slade, 1981. Bifurcation ratios and the adaptive geometry of trees. J. Botanical Gazette, 142: 394-401.
Direct Link - Neufeld, H.S., F.C. Meinzer, C.S. Wisdom, M.R. Sharifi and P.W. Rundel et al., 1988. Canopy architecture of Larrea trident at a (DC.) Co v., a desert shrub: Foliage orientation and direct beam radiation interception. J. Oecologia, 75: 54-60.
Direct Link - Muth, C.C. and F.A. Bazzaz, 2002. Tree canopy displacement at forest gap edges. Can. J. For. Res., 32: 247-254.
Direct Link - Caplat, P., M. Anand and C. Bauch, 2008. Symmetric competition causes population oscillations in an individual-based model of forest dynamics. Ecol. Model., 211: 491-500.
CrossRefDirect Link - Chelle, M., 2006. Could plant leaves be treated as Lambertian surfaces in dense crop canopies to estimate light absorption? Ecol. Model., 198: 219-228.
Direct Link - Meyer, K., 2001. Revision of the Southeast Asian genus Melastoma (Melastomaceae). Blumea, 46: 351-398.
Direct Link - Cornelissen, T. and P. Stiling, 2008. Clumped distribution of oak leaf miners between and within plants. J. Basic Applied Ecol., 9: 67-77.
CrossRef - Drouet, J.L. and B. Moulia, 1997. Spatial re-orientation of maize leaves affected by initial plant orientation and density. Agric. For. Meteorol., 88: 85-100.
CrossRef - Ezcurra, E., C. Montaña and S. Arizaga, 1991. Architecture, light interception and distribution of Larrea species in the Monte Desert, Argentina. J. Ecol., 72: 23-34.
CrossRef - Gatto, R. and S.R. Jammalamadaka, 2007. The generalized von mises distribution. Stat. Methodol., 4: 341-353.
CrossRefDirect Link - Getzin, S., C. Dean, F. He, J.A. Trofymow, K. Wiegand and T. Wiegand, 2006. Spatial patterns and competition of tree species in a douglas-fir chronose-quence on Vancouver Island. J. Ecography, 29: 671-682.
Direct Link - Getzin, S. and K. Wiegand, 2007. Asymmetric tree growth at the stand level: Random crown patterns and the response to slope. For. Ecol. Manage., 242: 165-174.
CrossRefDirect Link - Mardia, K.V. and P.E. Jupp, 2000. Statistics of Directional Data. John Wiley and Sons, Chicester, ISBN-13: 978-0124711501.
Direct Link - Girardin, P. and M. Tollenaar, 1994. Effects of intra-specific interference on maize leaf azimuth. Crop Sci., 34: 151-155.
Direct Link - Maddonnia, G.A., M. Chelleb, J.L. Drouetb and B. Andrieub, 2001. Light interception of contrasting azimuth canopies under square and rectangular plant spatial distributions: Simulations and crop measurements. Field Crops Res., 70: 1-13.
CrossRef - Kropff, M.J. and P.C. Struik, 2002. Developments in crop ecology. Netherlands J. Agric. Sci., 50: 223-237.
Direct Link - Judd, W.S., 1986. Taxonomic studies in the Miconieae (Melastomataceae). I. Variation in inflorescence position. Brittonia, 38: 150-161.
CrossRef