
Jaime Y.S. Low
Institute of Biological Sciences, Faculty of Science, University of Malaya, 50603 Kuala Lumpur, Malaysia
Noorlidah Abdullah
Institute of Biological Sciences, Faculty of Science, University of Malaya, 50603 Kuala Lumpur, Malaysia
S. Vikineswary
Institute of Biological Sciences, Faculty of Science, University of Malaya, 50603 Kuala Lumpur, Malaysia
Research Journal of Environmental Sciences
Year: 2009 | Volume: 3 | Issue: 3 | Page No.: 357-366







ABSTRACT
The ability of white rot fungus Pycnoporus sanguineus to colonize support materials and subsequently produce laccase and degradation of Polycyclic Aromatic Hydrocarbons (PAHs) was compared with free mycelia culture. Natural support, Ecomat, was found to be the best support material for P. sanguineus for mycelial colonization and laccase activity with maximum activity of 39 nkat mL-1 on day nine of incubation. Coconut husk and grey scouring sponge produces maximum laccase activity of 9.17 and 6.67 nkat mL-1, respectively on day 15. Pycnoporus sanguineus immobilized culture exhibited higher PAHs degradation efficiency compared to the free mycelia culture during the 20 days of incubation. The immobilized mycelia culture degraded 88% of phenanthrene, 93% of anthracene and 85% of pyrene within 20 days. The good correlation between the amount of PAHs degraded and laccase activity produced in the immobilization medium indicated that laccase was solely responsible for degradation of the three PAHs tested. In comparison, free mycelia culture rapidly degraded 42% of phenanthrene, 92% of anthracene and 87% of pyrene at the cessation of incubation. However, poor correlation between the amount of PAHs degraded and laccase activity measured in the free cell culture was obtained. This suggested that intracellular enzymes could be involved in PAHs degradation in the free cell culture.
PDF Abstract XML References
How to cite this article
Jaime Y.S. Low, Noorlidah Abdullah and S. Vikineswary, 2009. Evaluation of Support Materials for Immobilization of Pycnoporus sanguineus Mycelia for Laccase Production and Biodegradation of Polycyclic Aromatic Hydrocarbons. Research Journal of Environmental Sciences, 3: 357-366.
DOI: 10.3923/rjes.2009.357.366
URL: https://scialert.net/abstract/?doi=rjes.2009.357.366
DOI: 10.3923/rjes.2009.357.366
URL: https://scialert.net/abstract/?doi=rjes.2009.357.366
INTRODUCTION
Polycyclic Aromatic Hydrocarbons (PAHs), also known as polynuclear aromatic hydrocarbons or polyarenes, constitute a large class of organic compounds (Gomez et al., 2006). PAHs are highly water insoluble and are widespread pollutants in freshwater and seawater, particularly in estuaries and coastal waters with pollution from petroleum, coal or other heavy industries (Maldonado et al., 1999; Mitra et al., 1999).
Ligninolytic fungi, which colonize wood and other ligninocellulosic materials, are abundant in nature and have gained considerable attention for their bioremediation potential since the enzymes that are involved in lignin breakdown can also degrade a wide range of pollutants (Bumpus et al., 1985). These fungi which includes Phanerochaete chrysosporium (Brodkorb and Legge, 1992), Trametes versicolor (Field et al., 1992), Bjerkandera sp. strain BOS55 (Field et al., 1995) and Pleurotus ostreatus (Bezalel et al., 1997) have been extensively studied for their ability to degrade PAHs. Lignin peroxidase (LiP), manganese-dependent peroxidase (MnP), manganese-independent peroxidase and laccase are the enzymes that have been implicated in the oxidation of PAHs (Brodkorb and Legge, 1992; Bogan and Lamar, 1996; Pickard et al., 1999). These extracellular enzymes have low substrate specificity and diffuse into the soil matrix where the PAHs are entrapped.
Immobilization of microorganisms can be defined as any technique that limits the free migration of cells. There are two types of cell immobilization: entrapment and attachment. In the cell 45822_ja., 2004). Alginate, chitosan, chitin and cellulose derivatives have been used as matrix in the entrapment technique for cell immobilization (Arica et al., 1993). While in the cell attachment technique, synthetic foams like polyurethane foams (Yang and Yu, 1996), nylon sponges, natural materials such as jute, straw and hemp (Shin et al., 2002) was commonly employed for the attachment procedure. Immobilization by entrapment in a matrix such as alginate, chitosan and chitin may be too costly while surface immobilization via attachment on inexpensive materials such as synthetic sponges and natural materials are cheaper. Rodriguez Couto et al. (2004) had shown that the entrapment technique was not suitable for the immobilization of Trametes hirsuta, whereas the attachment technique using the stainless steel sponge as the support gave better results. Immobilized cultures tend to have a higher level of enzymes activity and were more resilient to environmental perturbations such as pH or exposure to toxic chemicals compared to free cell culture (Shin et al., 2002).
Mycelial immobilization has been used for the production of ligninolytic enzymes and bioremediation of pollutants such as chlorophenols (Sedarati et al., 2003), textile dyes (Ziegenhagen and Hofrichter, 2000) and mercury (Arica et al., 1993), but has not been applied to the biodegradation of PAHs. Thus, in this study, the immobilization of Pycnoporus sanguineus (Linn. Ex Fr.) Murrill strain CY788 to produce high laccase activity for PAHs degradation was evaluated.
MATERIALS AND METHODS
The study of the effect of various types of support materials for P. sanguineus immobilization, laccase production and PAHs degradation was conducted in 2004.
Fungal Strain
Pycnoporus sanguineus (Linn. ex Fr.) Murrill strainCY788 isolated in Thailand was maintained in 30% (v/v) glycerol at 4°C. The stock was inoculated and re-stimulated by growing on sterilized wood chip placed on Potato Dextrose Agar (PDA) plates.
Support Materials
The synthetic sponges and natural support materials were shown in Fig. 1. The chemical structures of the support materials were not studied. Selection of support materials was made based on literature (Rodriguez Cuoto et al., 2004) and the texture and porosity of the material, which encouraged colonization of mycelium onto the support. Synthetic sponges such as grey scouring sponge (Crocodile), white sponge (Scotch Brite), green scouring pad (Scotch Brite), yellow nylon pad (Tono), soft sponge (Scotch Brite) and nylon mesh (Scotch Brite) were selected. The synthetic supports pre-treatment method was as described in Rodriguez Cuoto et al. (2004). The natural support materials evaluated in this study were Ecomat (made of ligninocellulolytic fibres of empty fruit bunches of oil palm) obtained from Ecofibre Technology and coconut husk. Ecomat was cut into cubes and irregular sized shreds of coconut husk were dried overnight at 65°C in the oven and cooled in the desiccator before the dry weights were taken.
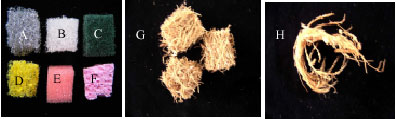 | |
| Fig. 1: | Synthetic and natural support materials selected for the immobilization of P. sanguineus; (A) Grey scouring sponge, (B) White sponge, (C) Green scouring pad, (D) Yellow nylon pad, (E) Soft sponge, (F) Nylon mesh, (G) Ecomat and (H) Coconut husk |
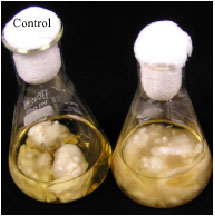 | |
| Fig. 2: | Ten days old fungal mycelial of immobilized P. sanguineus in GYMP medium (control) and medium exposed to 10 ppm PAHs |
Immobilization Procedure
The culture medium was Glucose-Yeast-Malt-Peptone (GYMP), containing, gram per litre, MgSO4.7H2O, 0.5; KH2PO4, 0.46; K2HPO4, 1.0; glucose, 20.0; peptone, 2.0; yeast extract, 2.0 and malt extract, 2.0. Two milliliters of 2% (v/v) P. sanguineus mycelial suspension was inoculated into 250 mL Erlenmeyer flask containing sterile 100 mL GYMP medium and the support material under aseptic condition. The flasks were incubated at 150 rpm at 29±2°C for 15 days. Mycelia of P. sanguineus was found to colonize the surface and within the porous matrix of Ecomat (Fig. 2).
Biomass Determination
Immobilized cultures were filtered through Whatman filter paper. The immobilized mycelia were washed twice with distilled water to remove mycelium which was not colonized on the surface of the support material and oven dried at 65oC until a constant weight was obtained (3-4 days).
Laccase Activity Assay
Laccase activity in the culture medium during immobilization of P. sanguineus on the selected support materials and free mycelia culture were assayed at intervals of three days for 15 days. Laccase activity was measured spectrophotometrically at 525 nm (Harkin and Obst, 1973). One unit (U) of enzyme activity was defined as the amount of enzyme producing 1OD unit in 1 mL of the culture medium after one minute. For conversion, 1U is equivalent to 16.67 nkat (SI unit).
Biodegradation of PAHs
The culture medium of 3-day-old immobilized and free cell culture was decanted and incubated with 100 mL sterile GYMP medium containing PAHs (anthracene, phenanthrene and pyrene) at 10 ppm final concentrations of each PAH. Cultures were incubated for 20 days.
Extraction of PAHs from Liquid Cultures
Residual PAHs in the medium was extracted with 20 mL of dichloromethane and shaken vigorously for 5 min in a 250 mL-separatory flask. The lower dichloromethane layer was collected and evaporated to dryness. The dried extract was then re-dissolved in 1 mL of acetonitrile and analyzed.
Analysis of PAHs
Samples were analyzed by high performance liquid chromatography (Spectra-Physics HPLC system). The column (150x4.6 mm ID) packed with Prosphere 300A PAH (5 μ particles) and coupled with guard column was used for PAHs separation. The flow rate was set at 1.5 mL min-1. Samples were eluted isocratically with an eluent consisting 38/62 (% v/v) water/acetonitrile. The PAHs were detected at 254 nm with a UV detector. Product inhibition studies were not conducted as degradation of PAHs was rapid and laccase activity was maintained at high levels throughout the whole incubation period, which suggested the by-products were not inhibitory to P. sanguineus and inhibit the biodegradation rate.
RESULTS
Growth of Free Mycelia and Immobilized Mycelia Culture
Figure 3 shows the dry weight increase in mycelial biomass of free mycelia culture and immobilized mycelia culture on different support materials. The growth in free mycelia culture of P. sanguineus was visible two days after inoculation. Fungal biomass as tiny pellets at day three was 188 mg and steadily increased to a maximum of 250 mg at day nine of incubation.
All support materials, except the Scotch Brite nylon mesh, facilitated mycelia colonization during the 15 days incubation. Pycnoporus sanguineus grew well on the synthetic supports without colour leaching from the supports and there was no loss in the integrity of the supports over a two-week period. The dry weight of biomass increased rapidly and maximum biomass concentration was obtained on day nine, after which the biomass level declined. Among the synthetic support materials selected in this study, there was good mycelial growth on grey sponge with highest dry mycelial biomass of 298 mg on day nine of incubation. This was followed by soft sponge, yellow nylon pad, white sponge, green scouring pad and antibiotic mesh.
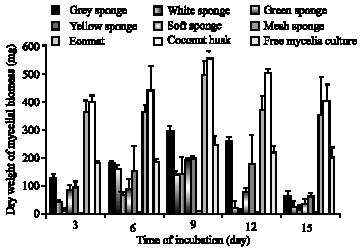 | |
| Fig. 3: | Comparison of mycelial biomass of P. Sanguineus in immobilized and free cell cultures. Results are expressed as the mean of triplicate values |
In contrast, natural support materials showed better colonization by P. sanguineus compared to synthetic support materials. The mycelial biomass was well attached to the support materials and the supports stayed intact throughout the incubation period. The culture grown on Ecomat and coconut husk gave higher mycelial biomass dry weights of 497 and 554 mg, respectively on day nine of incubation.
Laccase Activity of Free Mycelia and Immobilized Mycelia Cultures on Selected Support Materials
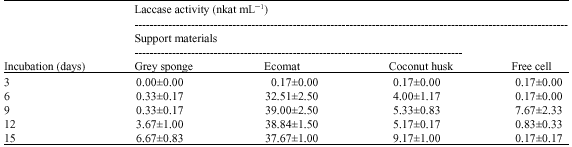
Previously, higher biomass was obtained in cultures immobilized on grey scouring sponge, Ecomat and coconut husk. Immobilization of mycelia on these support materials were further evaluated for the production of laccase and compared with free cell cultures. Table 1 shows the laccase activity of the free mycelia and immobilized mycelia cultures measured during the 15 days of incubation. Laccase activity was assayed on free mycelia culture and immobilized mycelia on grey sponge, Ecomat and coconut husk. The onset of laccase activity occurred after three days of incubation in free cell culture. Rapid laccase production only occurred after day six and attained its maximum activity of 7.63 nkat mL-1 on day nine of incubation. Comparatively, it was shown that optimum laccase production in free mycelia culture at day nine was 27 folds and 1.4 fold higher than cultures immobilized on grey sponge and coconut husk, respectively. In contrast, laccase activity obtained in free mycelia culture at day nine was five folds lower than immobilized culture on Ecomat. Thus, Ecomat was selected as the immobilization support for P. sanguineus for further studies of PAHs degradation.
Biodegradation of PAHs by P. sanguineus
All three PAHs studied in this study registered sharp exponential decline in concentration in the free mycelia culture after two days of incubation (Fig. 4). It was found that phenanthrene, anthracene and pyrene were efficiently degraded, with relative removals of over 13, 64 and 58%, to give final concentrations of 8.62, 3.55 and 4.15 ppm, respectively after two days of incubation. Anthracene was degraded most rapidly by the free mycelia culture of P. sanguineus with relative removal of over 92%, closely followed by pyrene and phenanthrene, with relative removal of over 87 and 42%, respectively, within 20 days of incubation. Their corresponding final concentrations were 5.74, 0.74 and 1.28 ppm for phenanthrene, anthracene and pyrene, respectively, at the cessation of incubation. Concomitant to the rapid PAHs degradation, laccase activity in free mycelia culture increased during the whole incubation period. However, the increase in degradation of PAHs by P. sanguineus free cell culture was not proportionate with the increase of laccase activity. Therefore, there were relatively low correlations between degradation of these PAHs and laccase activity (r2<0.67). This indicated that laccase may not be solely responsible for PAHs degradation in free mycelia culture.
| Table 1: | Comparison of laccase production by P. sanguineus immobilized on grey scouring sponge, Ecomat, coconut husk and free cell culture |
 | |
| Means of triplicates±SD | |
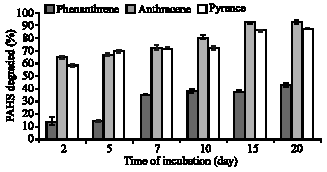 | |
| Fig. 4: | Percentage of PAHs degraded during 20 days of incubation with P. sanguineus free cell culture. Results are expressed as the mean of triplicate values |
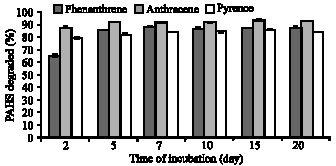 | |
| Fig. 5: | Percentage of phenanthrene degraded during the 20 days of incubation with immobilized culture P. sanguineus. Results are expressed as the mean of triplicate values |
The immobilized P. sanguineus culture proved to be more efficient in PAHs degradation as the degradation rate was more rapid compared to free cell culture. The PAHs degraded by immobilized culture on day two was over 65, 89 and 80%, with corresponding concentrations of 6.57, 8.91 and 8.03 ppm for phenanthrene, anthracene and pyrene, respectively (Fig. 5). At the cessation of the incubation, only 11.7, 6.3 and 14.9% of phenanthrene, anthracene and pyrene, respectively was recovered from the culture medium. Their corresponding final concentrations were 0.12, 0.06 and 0.15 ppm for phenanthrene, anthracene and pyrene, respectively. In this study, the high correlation coefficient (r2>0.96) between degradation of PAHs and laccase activity indicated that the increase in PAHs degradation was proportionate with the increase in laccase activity of P. sanguineus. Thus, laccase activity of the immobilized culture highly correlated with degradation of the PAH compounds.
Initially, removal of PAHs from the liquid culture was more rapid in the immobilized mycelia culture compared to the free mycelia culture. Degradation efficiency of phenanthrene, anthracene and pyrene in immobilized culture was 4.8, 1.4 and 1.4 fold higher than the free mycelia culture on day two of incubation, respectively. Subsequently, degradation efficiency of PAHs was reduced in the immobilized mycelia culture. Degradation efficiency of phenanthrene in immobilized mycelia culture was over 2 fold higher than the free mycelia culture at the cessation of incubation. Degradation efficiency of anthracene was comparable between immobilized mycelia and free mycelia cultures of P. sanguineus at the end of the incubation. Degradation efficiency of pyrene in immobilized mycelia culture was 2.4% lower than the free mycelia culture at the cessation of incubation.
DISCUSSION
This study had shown that the natural material such as Ecomat was able to maintain high levels of laccase production compared to free mycelia. The advantages of selecting natural materials as an immobilization support as these were compostable and inexpensive. Reports had shown that natural materials such as wheat straw (Zafar et al., 1996) and barley bran (Rodriguez Cuoto et al., 2003) had supported biofilms and sustained high enzyme activity. Shin et al. (2002) had also reported that natural supports such as jute, straw and hemp fiber were shown to provide the carbon source to the culture of Trametes versicolor and high levels of degrading enzymes were sustained during growth. In this study, immobilization of P. sanguineus on natural support materials supported better mycelial growth compared to the free mycelia culture. This could be due to these natural materials provide exogenous nutrients (carbon source from the breakdown of cellulose and lignin) to encourage biomass proliferation. Ecomat was found to be the best support for immobilization of P. sanguineus as the mycelium adhered to it readily and the biomass did not slough off easily. Ecomat made from 100% natural oil palm residues contained a higher proportion of easily accessible and biodegradable plant material such as cellulose or hemicellulose. Furthermore, immobilized culture on Ecomat was able to sustain high laccase productivity for longer incubation period compared to other supports. This could be due to the ability of P. sanguineus to metabolize the carbohydrates in Ecomat, making the whole process much more economical. Comparatively, the laccase activity of immobilized mycelia of P. sanguineus on Ecomat was higher than other species of white rot fungi cultivated in submerged liquid culture such as Pleurotus ostreatus (Medeiros et al., 1999) and Trametes spp. (Rodriguez Cuoto et al., 2003, 2004; Trupkin et al., 2003).Thus, this revealed the high potential of Ecomat to improve laccase production by P. sanguineus.
The degradation of each PAH by P. sanguineus in both free mycelia and immobilized culture in descending order was anthracene>pyrene>phenanthrene. Although phenanthrene and anthracene shared the same molecular weight (178.23 g mol-1), the structure of phenanthrene is more compact and angular compared to anthracene. This may be a contributory factor in its higher resistance to fungal enzyme attack. Furthermore, the Ionization Potential (IP) of anthracene (7.44 eV) is lower than phenanthrene (7.90 eV), indicating its higher susceptibility to enzymatic reactions (Dasbetani and Ivanov, 1999). Pyrene, on the other hand, was readily degraded by P. sanguineus free mycelia culture and immobilized culture compared to phenanthrene during the 20 days incubation period. Based on the results, there was no direct correlation between the number of aromatic rings and the metabolism of PAHs as pyrene is a four-ring PAH compound. The IP of pyrene (7.42 eV) was lower than both anthracene (7.44 eV) and phenanthrene (7.90 eV), which renders it liable to attack by ligninolytic enzymes (Hammel et al., 1986). Collins et al. (1996) had reported on laccases of Trametes versicolor, which were able to oxidize PAHs with IP lower than 7.45 eV and thus, was proven by the rapid degradation of anthracene and pyrene by laccase from P. sanguineus compared to phenanthrene in this study. The degradation of PAHs was rapid while laccase activity of the free mycelia culture was found to be low in the early stage of incubation. Results showed that correlation between PAHs degradation and laccase activity in free mycelia culture was relatively low. It was deduced that laccase was not responsible for PAHs degradation in free mycelia culture. Thus, this suggests the involvement of intracellular degradation of PAHs where intracellular enzymes like cytochrome P450 monooxygenases oxidized PAH compounds that were adsorbed to the biomass. Cytochrome P450 monooxygenase has been reported to cause the degradation of phenanthrene by Phanerochaete chrysosporium (Sutherland et al., 1991) and Pleurotus ostreatus (Bezalel et al., 1997). A preliminary study done by Verdin et al. (2004) assessing crude enzymatic extract of Fusarium solani grown on synthetic medium containing benzo[a]pyrene showed the presence of cytochrome P450 monooxygense and low levels of laccase activity. An assumption that intracellular enzymes of P. sanguineus were likely involved in the oxidation of PAHs, in which the degradation of PAH compounds are attacked by intracellular enzymes and extracellular laccase.
The capacity of P. sanguineus immobilized on Ecomat to remove anthracene, phenanthrene and pyrene exceeded the efficiency of the free mycelia culture. The rapid degradation of these compounds correlated with the increasing laccase activity during the whole incubation period. The high correlation between percentages of PAHs degraded and laccase activity observed could have resulted from a direct relationship, in which, laccase could be solely responsible for PAHs degradation in P. sanguineus. According to Braun-Lullemann et al. (1999), the removal of PAHs was probably due to the extracellular oxidative enzymes produced by the fungi. The direct involvement of laccase in PAHs degradation was further confirmed by Gramss et al. (1999), where PAHs degradation was correlated with laccase activity in wood-degrading fungi with correlation coefficient equivalent to 0.85.
Pycnoporus sanguineus tested in this study was proven to be an efficient PAHs degrader compared to some wood-degrading fungi included Bjerkandera adusta, Hypholoma frowardii and Pleurotus ostreatus reported by Gramss et al. (1999). They tested 58 fungal strains, consisting of wood-degrading basidiomycetes isolated from the soil. They had reported the ability of these basidiomycetes to remove 40-60% of phenanthrene, 72-82% of anthracene and 58-70% of pyrene in a mixture of five PAHs (total 10 ppm PAHs) in 14 days of incubation. However, P. sanguineus in both free cell and immobilized culture were able to remove 51-81% of phenanthrene, 93-94% of anthracene and 83-85% of pyrene in 15 days of incubation. While anthracene removal (total 10 ppm) were comparable with cultures inoculated with Phanerochaete chrysosporium and Bjerkandera sp. strain Bos55 as reported by Field et al. (1992). Therefore, P. sanguineus proved to be an efficient PAHs degrader comparable to other white rot fungi species. Furthermore, the involvement of laccase in the immobilized culture that played an important role in PAHs degradation was confirmed. This study also revealed the advantages of immobilization of P. sanguineus, particularly on Ecomat, for rapid PAHs degradation.
ACKNOWLEDGMENTS
The authors gratefully acknowledge the financial support of the University of Malaya for the Vote F grant F0144/2003C and Ministry of Science and Environment Grant IRPA 01-02-03-1002.
REFERENCES
- Arica, M.Y., F.A. Sharif, N.G. Alaeddinoglu, N. Hasirci and V. Harsici, 1993. Covalent entrapment of Aspergillus niger on pHAEMA membrane: Application to continuous-flow reactors. J. Chem. Technol., 58: 281-285.
Direct Link - Bezalel, L., Y. Hadar and C.E. Cerniglia, 1997. Mineralization of ploycyclic aromatic hydrocarbons by the white rot fungus Pleurotus ostreatus. Applied Environ. Microbiol., 62: 292-295.
Direct Link - Bogan, B.W. and R.T. Lamar, 1996. Polycyclic aromatic hydrocarbon-degrading capabilities of Phanerochaete laevis HHB-1625 and its extracellular ligninolytic enzymes. Applied Environ. Microbiol., 62: 1597-1603.
PubMedDirect Link - Braun-Lullemann, A., A. Huttermann and A. Majcherczyk, 1999. Screening of ectomycorrhizal fungi for degradation of polycyclic aromatic hydrocarbons. Applied. Microbiol. Biotechnol., 53: 127-132.
CrossRef - Brodkorb, T.S. and R.L. Legge, 1992. Enhanced biodegradation of phenanthrene in oil tar-contaminated soils supplemented with Phanerochaete chrysosporium. Applied. Environ. Micobiol., 58: 3117-3121.
Direct Link - Bumpus, J.A., M. Tien, D. Wright and S.D. Aust, 1985. Oxidation of persistent environmental pollutants by a white rot fungus. Science, 228: 1434-1436.
CrossRef - Collins, P.J., M.J.J. Kotterman, J.A. Field and A.D.W. Dobson, 1996. Oxidation of anthracene and benzo[a]pyrene by laccases from Trametes versicolor. Applied Environ. Microbiol., 62: 4563-4567.
Direct Link - Dasbetani, R and I.V. Ivanov, 1999. A compilation of physical, spectroscopic and photophysical properties of polycyclic aromatic hydrocarbons. Photochem. Photobiol., 70: 10-34.
CrossRef - Field, J.A., E.D. Jong, G.F. Costa and J.A.M. Bont, 1992. Biodegradation of polycyclic aromatic hydrocarbons by new isolates of white rot fungi. Applied Environ. Microbiol., 58: 2219-2228.
Direct Link - Field, J.A., F. Boelsma, H. Baten and W.H. Rulsken, 1995. Oxidation of anthracene in water/solvent mixtures by the white-rot fungus, Bjerkandera sp. strain BOS55. Applied Microbiol. Biotechnol., 44: 234-240.
CrossRef - Gomez, J., D.R. Solar, M. Pazos and M.A. Sanroman, 2006. Appliedicability of Coriolopsis rigida for biodegradation of polycyclic aromatic hydrocarbons. Biotechnol. Lett., 28: 1013-1017.
CrossRef - Gramss, G., B. Kirsche, K.D. Voigt, T. Gunther and W. Fritsche, 1999. Conversion rates of five polycyclic aromatic hydrocarbons in liquid cultures of fifty-eight fungi and the concomitant production of oxidative enzymes. Mycol. Res., 103: 1009-1018.
CrossRef - Harkin, J.M. and J.R. Obst, 1973. Syringaldazine, an effective reagent for detecting laccase and peroxidase in fungi. Experientia, 29: 381-387.
CrossRef - Hammel, K.E., B. Kalyanaraman and T.K. Kirk, 1986. Oxidation of polycyclic aromatic hydrocarbons and dibenzo[p]dioxins by Phanerochaete chrysosporium ligninase. J. Biol. Chem., 261: 16948-16952.
Direct Link - Maldonado, C., J.M. Bayona and L. Bodineau, 1999. Sources, distribution and water column processes of aliphatic and polycyclic aromatic hydrocarbons in the northwestern Black Sea water. Environ. Sci. Technol., 33: 2693-2702.
CrossRef - Medeiros, M.B., A.V. Bento, A.L.L. Nunes and S.C. Oliveora, 1999. Optimization of some variables that affect the synthesis of laccase by Pleurotus ostreatus. Bioprocess Eng., 21: 483-487.
CrossRef - Mitra, S, R.M. Dickhut, S.A. Kuehl and K.L Kimbrough, 1999. Polycyclic aromatic hydrocarbon (PAH) source, sediment deposition patterns and particle geochemistry as factors influencing PAH distribution coefficients in sediments of Elizabeth River, VA., USA. Mar. Chem., 66: 113-127.
CrossRef - Pickard, MA, R. Roman, R., Tinoco and R. Vazquez-duhalt, 1999. Polycyclic aromatic hydrocarbon metabolism by white rot fungi and oxidation by Coriolopsis gallica UAMH 8260 laccase. Applied. Environ. Microbiol., 65: 3805-3809.
Direct Link - Rodriguez Couto, S., D. Moldes, A. Liebanas and M.A. Sanroman, 2003. Investigation of several bioreactor for laccase production by Trametes versicolor operating in solid-state conditions. Biochem. Eng. J., 15: 21-26.
CrossRef - Couto, S.R., M.A. Sanroman, D. Hofer and G.M. Gubitz, 2004. Stainless steel sponge: A novel carrier for the immobilisation of the white-rot fungus Trametes hirsuta for decolourization of textile dyes. Bioresour. Technol., 95: 67-72.
CrossRefDirect Link - Sedarati, M.R., T. Keshavarz, A.A. Leontievsky and C.S. Evans, 2003. Transformation of high concentrations of chlorophenols by the white-rot basidiomycetes. Trametes versicolor immobilized on nylon mesh. Electronic J. Biotechnol., 6: 1-11.
Direct Link - Shin, M., T. Nguyen and J. Ramsay, 2002. Evaluation of support materials for the surface immobilization and decoloration of amaranth by Trametes versicolor. Applied Microbiol. Biotechnol., 60: 218-223.
CrossRef - Sutherland, J.B., A.L. Selby, J.P. Freeman, F.E. Evans and C.E. Cerniglia, 1991. Metabolism of phenanthrene by Phanerochaete chrysosporium. Applied. Environ. Mcirobiol., 57: 3310-3316.
Direct Link - Trupkin, S., L. Levin, A. Forchiassin and A. Viale, 2003. Optimization of a culture medium for ligninolytic enzyme production and synthetic dye decolorization using response surface methodology. J. Ind. Microbiol. Biotechnol., 30: 682-690.
CrossRefDirect Link - Verdin, A., A.L.H. Sahraoui and R. Durand, 2004. Degradation of benzo[a]pyrene by mitosporic fungi and extracellular oxidative enzymes. Int. Biodeterior. Biodegrad., 53: 65-70.
CrossRef - Yang, F.C. and J.T. Yu, 1996. Development of a bioreactor system using an immobilized white rot fungus for decolorization. Bioproc. Eng., 15: 307-310.
CrossRef - Zafar, S.I., N. Abdullah, M. Iqbal, Q. Sheeraz, 1996. Influence of nutrient amendment on the biodegradation of wheat straw during solid state fermentation of Trametes versicolor. Int. Biodeterior. Biodegrad., 38: 83-87.
CrossRef - Ziegenhagen, D. and M. Hofrichter, 2000. A simple and rapid method to gain high amounts of manganese peroxidase with immobilized mycelium of the agaric white-rot fungus Clitocybula dusenii. Applied Microbiol. Biotechnol., 53: 553-557.
CrossRef