
M. Hassanzadeh
Young Researchers Club, Department of Agronomy, Faculty of Agriculture, Islamic Azad University, Ardabil Branch, Ardabil, Iran
A. Ebadi
Department of Agronomy and Plant Breeding, Faculty of Agriculture, University of Mohaghegh Ardabili, Ardabil, Iran
M. Panahyan-e-Kivi
Department of Agronomy, Payam Noor University, Ardabil, Iran
A.G. Eshghi
Ardabil Research Center for Agriculture and Natural Resources, Moghan Station, Iran
Sh. Jamaati-e-Somarin
Young Researchers Club, Department of Agronomy, Faculty of Agriculture, Islamic Azad University, Ardabil Branch, Ardabil, Iran
M. Saeidi
Young Researchers Club, Department of Agronomy, Faculty of Agriculture, Islamic Azad University, Ardabil Branch, Ardabil, Iran
R. Zabihi-e-Mahmoodabad
Young Researchers Club, Department of Agronomy, Faculty of Agriculture, Islamic Azad University, Ardabil Branch, Ardabil, Iran
Research Journal of Environmental Sciences
Year: 2009 | Volume: 3 | Issue: 3 | Page No.: 345-350







ABSTRACT
In order to evaluate drought stress on relative water content (RWC) and chlorophyll content of 27 Sesame genotypes, a factorial experiment based on randomized complete block design with three replications was conducted in 2006 in Moghan region, Iran. Factors were 27 Sesame genotypes (Karaj 1, Yekta, Oltan, Moghan 17, Naz takshakheh, Naz chandshakheh, Borazjan 2, Borazjan 5, Darab 14, Varamin 37, Varamin 237, Varamin 2822, Zoodrass IS, Hendi, Chini, Yellow white, 5089, Panama, Do-1, TF-3 , TKG-21, J-I, RT-54, Hendi 9, Hendi 12, Hendi 14 and Jiroft) and irrigation levels (full irrigation and irrigation until flowering stage). Results showed that Varamin 2822 and Varamin 237 genotypes had the highest RWC of 84.100 and 81.217%, respectively. The most chlorophyll a content was observed in Hendi 9 genotype of 106.237, the most chlorophyll b in Karaj 1 genotype of 84.665 and the most chlorophyll total in Hendi genotype of 182.395 mg g-1 leaf fresh weight. It seems that Varamin 2822 genotype having the highest RWC and Hendi 9 and Hendi genotypes having the most chlorophyll a and chlorophyll total, respectively, are recommended for planting in arid and semi-arid conditions.
PDF Abstract XML References Citation
How to cite this article
M. Hassanzadeh, A. Ebadi, M. Panahyan-e-Kivi, A.G. Eshghi, Sh. Jamaati-e-Somarin, M. Saeidi and R. Zabihi-e-Mahmoodabad, 2009. Evaluation of Drought Stress on Relative Water Content and Chlorophyll Content of Sesame (Sesamum indicum L.) Genotypes at Early Flowering Stage. Research Journal of Environmental Sciences, 3: 345-350.
DOI: 10.3923/rjes.2009.345.350
URL: https://scialert.net/abstract/?doi=rjes.2009.345.350
DOI: 10.3923/rjes.2009.345.350
URL: https://scialert.net/abstract/?doi=rjes.2009.345.350
INTRODUCTION
Chlorophyll content is one of the major factors affecting photosynthetic capacity. Reduction or no-change in chlorophyll content of plant under drought stress has been observed in different plant species and its intensity depends on stress rate and duration (Rensburg and Kruger, 1994; Kyparissis et al., 1995; Jagtap et al., 1998). Chlorophyll content in plant is considered as a favorite aspect for plant growth (Farquhar and Richards, 1984). Chlorophyll content of leaf is indicator of photosynthetic capability of plant tissues (Nageswara et al., 2001; Wright et al., 1994). Flooding irrigation about 1 cm above soil surface led to senescence and decrease in chlorophyll content of leaves. In this study, severe drought stress resulted in increase in chlorophyll content and then, remained constant (Mensah et al., 2006). De-Souza et al. (1997) reported that soybean plants grown at the greenhouse and were subjected to drought stress from early seed filling stage to maturity stage, rapidly lost their leaf chlorophyll content than control plants.Schlemmer et al. (2005) stated that drought stress had no significant effect on chlorophyll content of maize leaf and concluded that decrease in turger pressure caused by water deficit, resulted in change in amount of far red radiation passed through the leaf and this reason, read of chlorophyll meter device was changed. In other words, light reflection from leaf was increased with increasing drought stress. Barry et al. (1992) reported chlorophyll destruction in barley as affected by water deficit. Xian-He et al. (1995) stated the same results in wheat. Also, Fotovat et al. (2007) found that by exerting severe drought stress on wheat, chlorophyll content of leaf significantly decreased.
In mid 80s, RWC was introduced as a best criterion for plant water status which, afterwards was used instead of plant water potential as RWC referring to its relation with cell volume, accurately can indicate the balance between absorbed water by plant and consumed through transpiration. Schonfeld et al. (1988) showed that wheat cultivars having high RWC, are more resistant against drought stress. Generally, it seems that osmoregulation is one of the main mechanisms preserving turger pressure in most plant species against water loss from plant so, it causes plant to continue water absorption and retain metabolic activities (Gunasekera and Berkowitz, 1992). Zlatko Stoyanov (2005) found that by exerting drought stress for 14 days and reaching soil water potential to -0.9 MPa, osmotic potential and turger pressure in first leaf of bean strongly was decreased. Ramos et al. (2003) stated that RWC of bean leaves under drought stress significantly was lesser than control. Lazacano-Ferrat and Lovat (1999) subjected bean plant to drought stress and after 10, 14 and 18 days after irrigation was withholded, they evaluated RWC of stem and found that RWC was significantly lower comparing with control plants. Gaballah et al. (2007) applied antitranspirant maters on two Sesame cultivars named Gize 32 and Shanavil 3 and observed that this matters by preventing water transpiration from leaves, led to in crease in RWC in these cultivars.
The objective of this research was to determine RWC and chlorophyll content of Sesame leaves under drought stress in Moghan region, Iran.
MATERIALS AND METHODS
In order to evaluate drought stress on relative water content (RWC) and chlorophyll content of 27 Sesame genotypes, a factorial experiment based on randomized complete block design with three replications was conducted in 2006 in Moghan region, Iran. Factors were 27 Sesame genotypes (Karaj 1, Yekta, Oltan, Moghan 17, Naz takshakheh, Naz chandshakheh, Borazjan 2, Borazjan 5, Darab 14, Varamin 37, Varamin 237, Varamin 2822, Zoodrass IS, Hendi, Chini, Yellow white, 5089, Panama, Do-1, TF-3, TKG-21, J-I, RT-54, Hendi 9, Hendi 12, Hendi 14 and Jiroft) and irrigation levels (full irrigation and irrigation until flowering stage). In order to calculate leaf chlorophyll content, 1 g punched fresh leaf sample was grinded along with 40 mL acetone 80% (v/v) until it was well smoothed.
Resulted green liquid was transferred through Whatman paper No. 2. Eventually, final liquid volume using acetone 80% reached to 100 mL. Thereafter, chlorophyll extract light densities were read using Spectrophotometer at 645, 663 and 652 mn wavelengths. Chlorophyll a, b and total as mg g-1 leaf fresh weight were calculated according to Dhopte and Manuel (2002):
Mg chlorophyll a = [12.7(D663) - 2.69 (D645)]xV/1000 w |
Mg chlorophyll b = [22.9(D645) - 4.68(D663)]xV/1000 w |
Mg chlorophyll total = [20.2(D645) + 8.02(D663)]xV/1000 w |
where, D is the chlorophyll extract light density, V is final volume of chlorophyll extract in acetone 80% and w is leaf fresh weight as gram. Also, in order to calculate RWC, leaf fresh weight samples were weighed, then were submerged in distilled water and finally were dried at 70 °C for 48 h and were weighed again. RWC was calculated according to Dhopte and Manuel (2002):
RWC = (FW- DW/TW-DW)x100 |
where, FW is fresh weight, DW is dry weight and TW is turger weight of leaf samples.
RESULTS AND DISCUSSION
Change of Leaf Chlorophyll
Effects of irrigation was significant (p<0.01) on leaf chlorophyll content. Chlorophyll a content significantly was higher (Table 1) in irrigation treatment than stress. Its value in irrigation and stress conditions was 80.039 and 77.239 mg g-1 leaf fresh weigh. Based on the results, it was revealed that genotypes with lowest yields, had the highest chlorophyll a content so that, Hendi 9 genotype which is classified in low yielding genotypes, significantly had the highest chlorophyll a. Chlorophyll b content under irrigation and stress treatments was obtained of 74.626 and 85.862 mg g-1 leaf fresh weight. The highest chlorophyll b was belonged to Karaj 1 genotype of 100.513 mg g-1 leaf fresh weight. Unlike the chlorophyll a, it is clear that the genotypes with the lowest yields had the lowest chlorophyll b contents. Also, chlorophyll total like the chlorophyll b content, significantly was higher (p<0.01) in irrigation treatment than stress. The highest chlorophyll total was observed in Hendi genotype of 182.395 mg g-1 leaf fresh weight.
| Table 1: | Mean comparisons of main effects of genotypes and irrigation levels on measured traits |
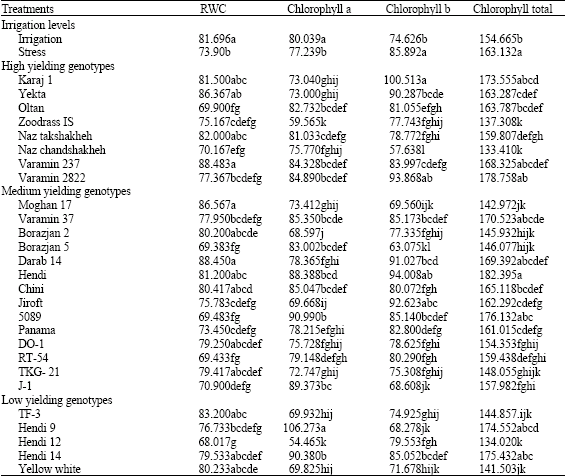 | |
| Numbers with the same letters, have no significant difference to each other | |
Results showed that the genotypes with the lowest yield had the lowest chlorophyll total and vice versa. Water deficit can destroy the chlorophyll and prevent making it (Lessani and Mojtahedi, 2002). Also, some researchers have reported damages to leaf pigments as a result of water deficit (Montagu and Woo, 1999; Nilsen and Orcutt, 1996). Mensah et al. (2006) found that subjecting Sesame to drought stress caused leaf chlorophyll was increased and then remained unchanged. Beeflink et al. (1985) reported increase in chlorophyll in onion under drought stress. Mentioned results are in accordance with this results confirming increase in chlorophyll b and total under drought stress. Ramalho et al. (2000) stated that ratio of chlorophyll a/b in late summer is less than spring of 2.6 units which may be attributed to more precipitations in spring. A reason for decrease in chlorophyll content as affected by water deficit is that drought or heat stress by producing reactive oxygen species (ROS) such as O2¯ and H2O2, can lead to lipid peroxidation and consequently, chlorophyll destruction (Mirnoff, 1993; Foyer et al., 1994). Also, with decreasing chlorophyll content due to the changing green color of the leaf into yellow, the reflectance of the incident radiation is increased (Schlemmer et al., 2005). It seems that this mechanism can protect photosynthetic system against stress. According to the Lawlor and Cornic (2002) reduction of carbon assimilation confronting water deficit, is due to limitation of Rubisco synthesis and ATP storage. Studies done as in vivo, showed that water deficit resulted in destruction of D1 protein of photosystem 2 (Xian-He et al., 1995) but the reason have not been known, yet.
Change of Relative Water ContentMensah
Irrigation and genotypes significantly (p<0.01) affected RWC (Table 1). In full irrigation, RWC was 81.69% and in stress, it was 73.90%. Moghan 17, Darab 14 and Varamin 237 genotypes had the highest values of 86.56, 88.45 and 88.48%, respectively. Hendi 12 genotype had the lowest RWC of 68.02%. Mensah et al. (2006) reported that with decreasing irrigation, RWC in Sesame decreased from 79.8 to 66.5%. Leaf RWC is of the best growth/biochemical indices revealing the stress intensity (Alizadeh, 2002). The rate of RWC in plants with high resistance against drought is higher than others. In other words, plant having higher yields under drought stress should have high RWC. So, based on the results, mentioned genotypes which are classified as high and medium yielding genotypes, should be of high-content RWC. Decrease in RWC in plants under drought stress may depend on plant vigor reduction and have been observed in many plants (Liu et al., 2002). Under water deficit, cell membrane subjects to changes such as increase in penetrability and decrease in sustainability (Blokhina et al., 2003). Microscopic investigations of dehydrated cells, revealed damages including cleavage in the membrane and sedimentation of cytoplasm content (Blackman et al., 1995). Probably, in these conditions, ability to osmotic adjustment is reduced (Meyer and Boyer, 1981). It seems that concentration of appropriate solutes to preserve membrane is not sufficient in this case.
Relations Between RWC and Chlorophyll Content
As shown in Table 2, RWC had the positive and significant correlations with chlorophyll b and total but non-significant and negative correlation with chlorophyll a. Also chlorophyll a had the negative and significant correlation with chlorophyll b and had non-significant correlation with chlorophyll total. In addition, chlorophyll b had non-significant correlation with chlorophyll total, as well.
| Table 2: | Correlations between measured traits |
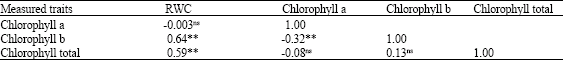 | |
| ns, * and ** are non significant, significant at p<0.01 and significant at p<0.05, respectively | |
CONCLUSIONS
As shown, drought stress making mechanisms inside the plant, leads to decrease in chlorophyll a but increase in chlorophyll b and total. Also leaf RWC was decreased as affected by drought. The highest chlorophyll a, b and total was belonged to Hendi 9, Karaj 1 and Hendi genotypes, respectively which two latter ones were of high and medium yielding genotypes. So, this indicates that Sesame genotypes with high yields include high chlorophyll b and total under drought conditions. Moreover, Moghan 17, Darab 14 and Varamin 237 genotypes which were among the high and medium yielding genotypes, had the highest RWC. So, mentioned genotypes (because of retaining chlorophyll and RWC against drought along with the high yields) in order to planting as dry-farming, are recommended.
ACKNOWLEDGMENTS
This study was supported by the Central Laboratory of Agricultural Faculty, University of Mohaghegh Ardabili. Valuable experimental support by Aziz Jamaati-e-Somarin and Assad Gholizadeh is greatly appreciated. This study was extracted from M.Sc. Thesis of Mohammad Hassanzadeh.
REFERENCES
- Barry, P., R.P. Evershed, A. Young, M.C. Prescott and G. Britton, 1992. Characterization of carotenoid acyl esters produced in drought-stressed barley seedlings. Phyto-Chemistry, 9: 3163-3168.
Direct Link - Blackman, S.A., R.L. Obendorf and A.C. Leopold, 1995. Desiccation tolerance in developing soybean seeds: The role of stress proteins. Plant Physiol., 93: 630-638.
CrossRefDirect Link - Blokhina, O., E. Virolainen and K.V. Fagerstedt, 2003. Anti oxidants, oxidative damage and oxygen deprivation stress. Ann. Bot., 91: 179-194.
CrossRef - Brevedan, R.E. and D.B. Egli, 2003. Short periods of water stress during seed filling, leaf senescence and yield of soybean. Crop Sci., 43: 2083-2088.
Direct Link - Chakir, S. and M. Jensen, 1999. How does Lobaria pulmoria regulate photosystem II during progressive desiccation and osmotic water stress? A chlorophyll fluorescence study at room temperature. Physiol Plant., 105: 257-265.
CrossRefDirect Link - De-Souza, P.I., D.B. Egli and W.P. Bruening, 1997. Water stress during seed filling and leaf senescence in soybean. Agron. J., 89: 807-812.
Direct Link - Fotovat, R., M. Valizadeh and M. Toorchi, 2007. Association between water-use-efficiency components and total chlorophyll content (SPAD) in wheat (Triticum aestivum L.) under well-watered and drought stress conditions. J. Food. Agric. Environ., 5: 225-227.
Direct Link - Gaballah, M.S., B. Abou Leila, H.A. El-Zeiny and S. Khalil, 2007. Estimating the performance of salt-stressed sesame plant treated with antitranspirants. J. Applied Sci. Res., 3: 811-817.
Direct Link - Gunasekera, D. and G.A. Berkowitz, 1992. Evaluation of contrasting cellular-level acclimation responses to leaf water deficits in three wheat genotypes. Plant. Sci., 86: 1-12.
Direct Link - Jagtap, V., S. Bhargava, P. Sterb and J. Feierabend, 1998. Comparative effect of water, heat and light stresses on photosynthetic reactions in Sorghum bicolor (L.) moench. J. Exp. Bot., 49: 1715-1721.
CrossRefDirect Link - Kyparissis, A., Y. Petropoulou and Y. Manetas, 1995. Summer survival of leaves in a soft-leaved shrub (Phlomis fruticosa L., Labiatae) under Mediterranean field conditions: Avoidance of photoinhibitory damage through decreased chlorophyll contents. J. Exp. Bot., 46: 1825-1831.
Direct Link - Lawlor, D.W. and G. Cornic, 2002. Photosynthetic carbon assimilation and associated metabolism in relation to water deficits in higher plants. Plant Cell Environ., 25: 275-294.
CrossRefDirect Link - Lazacano-Ferrat, I. and C.J. Lovat, 1999. Relationship between relative water content, nitrogen pools, and growth of Phaseolus vulgaris L. and P. acutifolius A. Gray during water deficit. Crop. Sci., 39: 467-475.
Direct Link - Liu, Y., G. Fiskum and D. Schubert. 2002. Generation of reactive oxygen species by the mitochondrial electron transport chain. J. Neurochem., 80: 780-787.
CrossRefDirect Link - Mensah, J.K., B.O. Obadoni, P.G. Eroutor and F. Onome-Irieguna, 2006. Simulated flooding and drought effects on germination, growth, and yield parameters of Sesame (Sesamum indicum L.). Afr. J. Biotechnol., 5: 1249-1253.
Direct Link - Meyer, R.F. and J.S. Boyer, 1981. Osmoregulation solute distribution and growth in soybean seedlings having low water potentials. Planta, 151: 482-489.
CrossRefDirect Link - Montagu, K.D. and K.C. Woo, 1999. Recovery of tree photosynthetic capacity from seasonal drought in the wet-dry tropics: The role of phyllode and canopy processes in Acacia auriculiformis. Aust. J. Plant Physiol., 26: 135-145.
CrossRefDirect Link - Nageswara Rao, R.C., H.S. Talwar and G.C. Wright, 2001. Rapid assessment of specific leaf area and leaf nitrogen in peanut (Arachis hypogaea L.) using a chlorophyll meter. J. Agron. Crop Sci., 189: 175-182.
CrossRefDirect Link - Ramalho, J.C., J.A. Lauriano and M.A. Nunes, 2000. Changes in photosynthetic performance of Ceratonia siliqua in summer. Photosynthetica, 38: 393-396.
Direct Link - Ramos, M.L.G., R. Parsons, J.I. Sprent and E.K. Games, 2003. Effect of water stress on nitrogen fixation and nodule structure of common bean. Pesq. Agropec. Bras. Brasilia., 38: 339-347.
Direct Link - Rensburg, L.V. and G.H.J. Kruger, 1994. Evaluation of components of oxidative stress metabolism for use in selection of drought tolerant cultivars of Nicotiana tabacum L. J. Plant Physiol., 143: 730-737.
Direct Link - Schlemmer, M.R., D.D. Francis, J.F. Shanahan and J.S. Schepers, 2005. Remotely measuring chlorophyll content in corn leaves with differing nitrogen levels and relative water content. Agron. J., 97: 106-112.
CrossRefDirect Link - Schonfeld, M.A., R.C. Johnson, B.F. Carwer and D.W. Mornhinweg, 1988. Water relations in winter wheat as drought resistance indicators. Crop. Sci., 28: 526-531.
Direct Link - Skotnica, J., M. Matouskova, J. Naus, D. Lazar and L. Dvooak, 2000. Thermoluminescence and fluorescence study of changes in Photosystem II photochemistry in desiccating barley leaves. Photosynth. Res., 65: 29-40.
CrossRefDirect Link - Smirnoff, N., 1993. The role of active oxygen in the response of plants to water deficit and desiccation. New Phytol., 125: 27-58.
CrossRefDirect Link - Wright, G.C., R.C. Nageswara Rao and G.D. Farquhar, 1994. Water use efficiency and carbon isotope discrimination in peanut under water deficit conditions. Crop Sci., 34: 92-97.
Direct Link - Xian-He, J., J. Wang and H. Guo Liang, 1995. Effects of water stress on photochemical function and protein metabolism of photosystem II in wheat leaves. Physiol Plant., 93: 771-777.
Direct Link - Yordanov, I., V. Velikova and T. Tsonev, 2003. Plant responses to drought and stress tolerance. Bulg. J. Plant Physiol., 29: 187-206.
Direct Link - Zlatko Stoyanov, Z., 2005. Effects of water stress on leaf water relations of young bean. J. Cent. Eur. Agric., 6: 5-14.
Direct Link - Foyer, C.H., K. Lelandais and K.J. Kunert, 1994. Photooxidative stress in plants. Physiol. Plant., 92: 696-717.
CrossRef - Farquhar, G.D. and R.A. Richards, 1984. Isotopic composition of plant carbon correlates with water-use efficiency of wheat genotypes. Aust. J. Plant Physiol., 11: 539-552.
CrossRefDirect Link