
S. Abd-Aziz
Department of Bioprocess Technology, Faculty of Biotechnology and Biomolecular Sciences, Universiti Putra Malaysia, 43400 Serdang, Selangor, Malaysia
N.A. Ab-Razak
Department of Bioprocess Technology, Faculty of Biotechnology and Biomolecular Sciences, Universiti Putra Malaysia, 43400 Serdang, Selangor, Malaysia
M.H. Musa
Department of Soil Microbiology, Faculty of Agriculture, Universiti Putra Malaysia, 43400 Serdang, Selangor, Malaysia
M.A. Hassan
Department of Bioprocess Technology, Faculty of Biotechnology and Biomolecular Sciences, Universiti Putra Malaysia, 43400 Serdang, Selangor, Malaysia
Research Journal of Environmental Sciences
Year: 2009 | Volume: 3 | Issue: 2 | Page No.: 251-256







ABSTRACT
The aim of this study is to produce and profile the mannan-degrading enzymes from local fungal isolates by submerged fermentation and saccharification of PKC. Results showed that Aspergillus niger and Sclerotium rolfsii can produce mannan-degrading enzymes. The fungi were grown in submerged fermentation of PKC to produce mannan-degrading enzymes. The highest alpha-galactosidase was obtained on day 13 of fermentation (0.128 U mL-1) when using A. niger and on day 18 (0.126 U mL-1) when using S. rolfsii. Analysis also showed that enzyme activities for beta-mannanase using S. rolfsii were the highest at day 17 (3.166 U mL-1) and for A. niger (2.482 U mL-1) at day 8. Meanwhile the highest beta-mannosidase were obtained at day 16 for A. niger (0.128 U mL-1) and for S. rolfsii at day 16 (0.116 U mL-1).
PDF Abstract XML References Citation
How to cite this article
S. Abd-Aziz, N.A. Ab-Razak, M.H. Musa and M.A. Hassan, 2009. Production of Mannan-Degrading Enzymes from Aspergillus niger and Sclerotium rolfsii Using Palm Kernel Cake as Carbon Source. Research Journal of Environmental Sciences, 3: 251-256.
DOI: 10.3923/rjes.2009.251.256
URL: https://scialert.net/abstract/?doi=rjes.2009.251.256
DOI: 10.3923/rjes.2009.251.256
URL: https://scialert.net/abstract/?doi=rjes.2009.251.256
INTRODUCTION
Palm kernel cake (PKC) is a by-product obtained from the palm oil extraction, which constitutes about 50% of the original oil palm (Elaeis guineensis) kernel. The PKC comprises mainly cell wall. This cell wall consists of major polysaccharides chains, which comprise mannan (57.8%), cellulose (11.6%) and a small amount of xylan (3.7%). PKC has a good potential as a protein source due to its relatively high protein content and essential amino acid profile (Iluyemi et al., 2006). However, little attention has been given to its because of mannan, the main component in PKC is a very hard, insoluble component, high crude fiber content, resistant to enzymatic degradation and largely constitutes of low branching structure. Thus, they are crystalline, insoluble and low in digestibility.
Since mannan is the main component in PKC, it would induce the production of mannan-degrading enzymes and end-product of mannose (Sachlehner and Haltrich, 1999; Ong et al., 2004). If the enzymatic depolymerisation of the mannan could be achieved, it would release the monomer mannose, with a small amount of galactose and possibly render the cellulose degradable by cellulose at the same time. Mannose is an isomer of glucose, having a different configuration at the second atom carbon. Mannan-degrading enzymes from fungal origin have extensively been studied such as Aspergillus niger, Sclerotium rolfsii, Trichoderma harzianum, Trichoderma konigii and Trichoderma longibrachiatum in solid and liquid fermentation (Iluyemi et al., 2006).
These mannan-degrading enzymes are in groups of hemicellulose-degrading enzymes, which hydrolyse specifically glycans that make up the backbone of the hemicelluloses, producing water-soluble monomers and oligosaccharides. Mannan-degrading enzymes have been used quite extensively and have an enormous range of applications of enzymes in industry, thus several study in purification and characterization have been conducted (Großwindhager et al., 1999). They are widely applied in the animal feed industry, for beverage processing, food processing, fruit-juices industries, paper industry and for therapeutic use. The new cheaper mannan-degrading enzyme is being investigated as commercial enzymes are expensive, hardly affordable, as the use for mannan-degrading enzymes has increased in many kinds of industry.
The aim of this study was to investigate the production of mannan-degrading enzymes using palm kernel cake as sole carbon source by submerged fermentation using two local isolated strains, Aspergillus niger and Sclerotium rolfsii.
MATERIALS AND METHODS
Microorganisms
Aspergillus niger was obtained from Mr. Onn bin Haji Ahmad of the Department of Biochemistry and Microbiology, while Sclerotium rolfsii was provided by Prof. Dr. Sariah Meon from the Department of Plant Protection, Universiti Putra Malaysia. Stock cultures were maintained on potato dextrose agar (PDA). Sub-culture was done every 4-5 weeks and after incubation at ambient temperature for 6 days and stored at 4°C for 2 h until further used. This study was undertaken in year 2007.
Growth and Culture Conditions
Potato dextrose agar (39 g) was suspended in 1 L of distilled water and boiled until soluble using microwave. It was then sterilized by autoclaving for 15 min at 121°C and PDA (15 mL) poured in agar plate. Pure cultures were sub-cultured onto PDA agar and incubated at 25°C for 6 days to obtain high concentration of spores for A. niger and mycelial at for S. rolfsii.
Substrate
PKC was obtained from Livestock Research Centre, MARDI. It was extracted by mechanical extraction using screw press. The particle size of PKC was about 0.5 mm sieve and stored in plastic bags at 4°C until further use. Prior to use, it was left for about 1 h at 25°C.
Fermentation Media
Aspergillus niger
The carbon source, PKC, was used in the preparation of the fermentation conditions. Inoculum preparations were conducted under submerged fermentation in 500 mL unbaffled shake flask with total volume of 250 mL. The fermentation media for Aspergillus niger composed of 0.1 g L-1 NH4SO4, 0.03 g L-1, KHPO4 and 0.4 g L-1 PKC (Laukevies et al., 1984) and inoculated aseptically with 10% (v/v) of 107 spores mL-1 at the initial stage into a sterile fermentation media for the fermentation.
Sclerotium rolfsii
The media used for fermentation of Sclerotium rolfsii followed the method described by Großwindhager et al. (1999) except that PKC was used in place of carbon source. One cm2 of 6-day-old mycelia mat was inoculated aseptically into 100 mL sterile fermentation media for the fermentation. Fermentation media was carried out using submerged fermentation in 500 mL unbaffled shake flask with total volume of 250 mL. All flasks were sealed with cotton-wool plugs.
Enzyme Activity Assay
Alpha-Galactosidase
Alpha-galactosidase activities were determined by monitoring the release of para-nitrophenol from the respective assayed as described by Großwindhager et al. (1999) in 50 mM sodium citrate buffer, pH 4.5.
Beta-Mannanase
Beta-mannanase activity was measured by method described by Michael (2000) using azo-carob galactomannan (2%) diluted in 2 M sodium acetate buffer, pH 4.5 as substrate.
Beta-Mannosidase
Beta-mannosidase activity was determined by monitoring the released of p-nitrophenol from the respective assayed as described by Großwindhager et al. (1999) in 50 mM sodium citrate buffer, pH 4.5.
Sample Assays
Total Soluble Protein Concentration
Soluble protein assay was used to indirectly assess the fungal biomass. Soluble protein content was determined using a modified Lowry method (Hartree, 1972) with BSA as standard.
RESULTS AND DISCUSSION
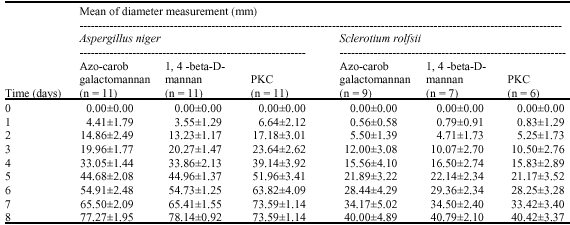
The growth of A. niger and S. rolfsii in carbon-limited medium gave different ability to produce mannan-degrading enzyme by converting mannan and galactomannan into simple sugar (mannose). The formation of clear zone was found to be a good identification for the quantification of mannan-degrading enzymes producer capability. Results showed that PKC have strong inducing effect on the formation of mannan-degrading enzymes by showing the formation of clear zone (Table 1).
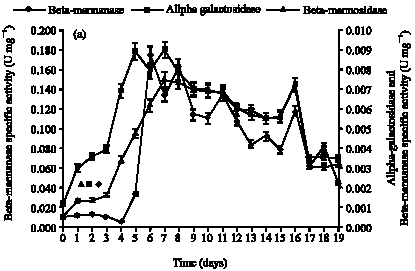
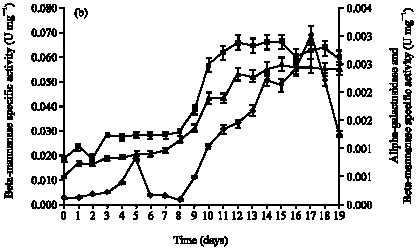
In submerged fermentation using A. niger (Fig. 1a), the activities of beta-mannanase was 2.48 U mL-1 which were maximum on day 8; alpha-galactosidase and beta-mannosidase activities were higher on day 13 with 0.13 U mL-1 and day 15 with 0.13 U mL-1, respectively. For mannan-degrading enzymes production by S. rolfsii (Fig. 1b), the activities of beta-mannanase was 3.17 U mL-1 with maximum activity at day 17; alpha-galactosidase and beta-mannosidase activities were higher on day 18 with 0.13 U mL-1 and day 16 with 0.12 U mL-1, respectively.
Since mannan is the main component in PKC, it would induce the production of mannan-degrading enzymes and end-product of mannose (Iluyemi et al., 2006; Sachlehner and Haltrich, 1999; Sachlehner et al., 1998). Formation of mannanase only commenced when glucose was depleted in the medium (Sachlehner and Haltrich, 1999; Großwindhager et al., 1999). The presence of hemicellulose in PKC had enhanced the production of mannan-degrading enzymes and reducing sugars. Besides, there is a possibility that the presence of mannan and hemicellulose in PKC may have a synergistic effect on the enzyme productions. For growth on a complex substrate such as PKC, fungi require several enzymes to be induced at the same time. The fermentation profile of Aspergillus niger and Sclerotium rolfsii is important in understanding the pattern of fermentation. Beta-mannan in PKC was found to induce the secretion of the enzyme from A. niger and S. rolfsii. The increase and decrease in enzyme activity may be due to induction and repression of hemicellulose enzymes.
| Table 1: | Mean of diameter measurement of clear zone by Aspergillus niger and Sclerotium rolfsii on azo-carob galactomannan, 1, 4-beta-D-mannan and PKC media |
 | |
| Fig. 1: | Profile for mannan-degrading enzymes using palm kernel cake as carbon source for (a) A. niger and (b) S. rolfsii |
At the initial stage of fermentation, the fungi grew and colonised the substrate and there was increased of enzyme activities (Fig. 2). After certain time, the PKC degradation process occurred thus the fluctuation of the enzymes were observed with time. The increase mycelial concentration which considered as the fungi growth resulted in an almost linear increase in the enzyme activities formed (Sachlehner et al., 1998). It can be explained by the fact that the fungi secretes enzymes to degrade hemicellulose and feeds on the products of degradation. It then resumes the secretion of more enzymes when the degradation products are being depleted. However under stress conditions, sugar can be accumulated leading to catabolite repression of enzymes synthesis, which is referred to as the glucose effect, glucose repression or carbon repression (Großwindhager et al., 1999). It was suggested that Aspergillus niger and Sclerotium rolfsii secretes the necessary enzymes for degradation of the available substrate molecules in order to meet their nutritional requirement (Iluyemi et al., 2006). Synthesis of enzymes, which are necessary for the degradation of energy-yielding polymeric material to be used as substrates for cell growth. Moloney et al. (1983) noted that the use of impure substrate, especially the PKC to be hydrolysed appears to induce the synthesis of a wider range of enzymes all of which contribute to an increased release of reducing sugars.
| Fig. 2: | Growth profile fermentation PKC using (a) A. niger and (b) S. rolfsii |
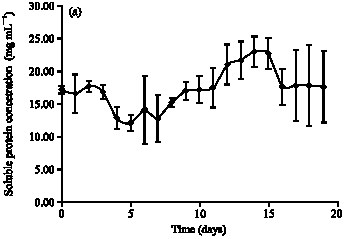
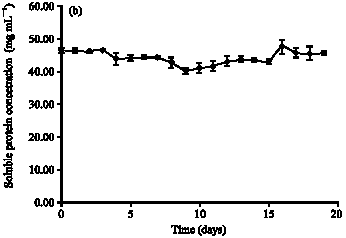
The lag phase follows inoculation of the nutrient medium is a period of adaptation. After that, synthesis or repression of enzymes or structural components of the cell reorganise both its micromolecular and macromolecular constituents. Therefore, the lag phase is short about 4 days. During this phase, cell mass may change without a change in cell number. The soluble low molecular weight catabolites can easily enter the cell, signal the presence of an extracellular substrate and provide the stimulus for the accelerated synthesis of the respective enzymes.
At some point, the growth rate started to decrease, either because of the depletion of essential nutrients, or because of the accumulation of a inhibitory products. Nevertheless, the cells go through a transition until net growth rate was zero (Christa, 1990; Großwindhager et al., 1999).
The fermentation profile of A. niger and S. rolfsii is important for understanding the growth pattern of both fungi studied. Beta-mannan in PKC was found to induce the secretion of the mannan-degrading enzyme from A. niger and S. rolfsii. The increase and decrease in mannan-degrading enzyme activity may be due to induction and repression of hemicellulose enzymes.
This study showed that mannan-degrading enzymes are essential in degrading mannan in PKC into simple sugars; beta-mannanase was the most abundant enzyme produced by S. rolfsii compared to A. niger in the submerged fermentation. The production of mannan-degrading enzymes from A. niger and S. rolfsii was relatively high using PKC as the carbon sources under the fermentation conditions employed (aeration at 40 L min-1 for 10 min daily and agitated continuously on orbital shaker at 180 rpm).
CONCLUSION
The results demonstrated the importance of liquid-solid fermentation for the production of enzyme using palm kernel cake as substrate, which offer significance benefit due to cheaper cost and abundant availability. This also showed that mannan-degrading enzymes are essential in degrading mannan in PKC into simple sugar; beta-mannanase was the most abundant enzyme produced by S. rolfsii in the submerged fermentation. From the parameter observed in this study, it can be concluded that, production of mannan-degrading enzymes are associated with growth of fungus. The production of mannan-degrading enzymes from Aspergillus niger and Sclerotium rolfsii was relatively high using PKC as the carbon sources under the fermentation conditions employed.
ACKNOWLEDGMENTS
The authors would like to thank the National Biotechnology Directorate, Malaysia for their financial support of this study and the Livestock Research Centre, MARDI, Serdang for their collaboration and technical assistance.
REFERENCES
- Hartree, E.F., 1972. Determination of protein: A modification of the lowry method that gives a linear photometric response. Anal. Biochem., 48: 422-427.
CrossRefPubMedDirect Link - Iluyemi, F.B., M.M. Hanafi, O. Radziah and M.S. Kamarudin, 2006. Fungal solid state culture of palm kernel cake. Bioresour. Technol., 97: 477-482.
CrossRefPubMedDirect Link - Laukevies, J.J., A.F. Apsite, U.E. Viesturs and R.P. Tengerdy, 1984. Solid-substrate fermentation of wheat straw to fungal protein. Biotechnol. Bioeng., 26: 1465-1474.
CrossRefPubMedDirect Link - Moloney, A.B., P.J. Considine and M.P. Coughlan, 1983. Cellulose hydrolysis by the cellulase produced by Taloromyces emersonii when grown on different inducing substrates. Biotechnol. Bioeng., 25: 1169-1173.
CrossRefDirect Link - Sachlehner, A. and D. Haltrich, 1999. Purification and some properties of a thermostable acidic endo-β-1,4-D-mannanase from Sclerotium (Athelia) rolfsii. FEMS Microbiol. Lett., 177: 47-55.
CrossRefDirect Link - Sachlehner, A., B. Nidetzky and K.D. Kulbe, 1998. Induction of mannanase, xylanases and endoglucanase activities in Sclerotium rolfsii. Applied Environ. Microbiol., 64: 594-600.
PubMedDirect Link - Ong, L.G.A., S. Abd-Aziz, S. Noraini, M.I.A. Karim and M.A. Hassan, 2004. Enzyme production and profile by Aspergillus niger during solid substrate fermentation using palm kernel cake as substrate. Applied Biochem. Biotechnol., 118: 73-79.
Direct Link - Grobwindhager, C., A. Sachslehner, B. Nidetzky and D. Haltrich, 1999. Endo-β-1,4-d-mannanase is efficiently produced by Sclerotium (Athelia) rolfsii under derepressed conditions. J. Biotechnol., 67: 189-203.
CrossRef