
M.A. Salam
Biotechnology and Genetic Engineering Discipline, Khulna University, Khulna, Bangladesh
M.S. Hossain
Biotechnology and Genetic Engineering Discipline, Khulna University, Khulna, Bangladesh
M.E. Ali
Gaco Pharmaceuticals and Research Laboratory, Dhaka-1000, Bangladesh
M.A. Asad
Fisheries and Marine Resource Technology Discipline, Khulna University, Bangladesh
M.H. Ali
Biotechnology and Genetic Engineering Discipline, Khulna University, Khulna, Bangladesh
Research Journal of Environmental Sciences
Year: 2009 | Volume: 3 | Issue: 1 | Page No.: 110-115







ABSTRACT
The experiment was carried out to determine the presence and numbers of arsenic resistant bacteria population. In this study, soil samples were collected from different contaminated sites of Khulna shipyard, Rupsha, Baghmara and Ramnagor in Khulna district of Bangladesh. Twenty arsenic resistant bacterial strains were isolated from the soil samples. They were isolated by growing them on Nutrient Broth (NB) medium impregnated with high concentration of arsenic. From them, six strains Bacillus lichnefomis (1/10), Listeria murrayi (2/9), Bacillus polymyxa (3/6), Moraxella urethralis (4/9) and Planococcus citreus (5/8) and Pseudomonas fluorescens (6/8) were finally selected and studied their morphological and biochemical characters in details. All six strains were able to tolerate very high concentration (>100 ppm). The optimum pH and temperature for the growth of all six strains were 8.5 and 37°C, respectively. It was concluded that bacteria living in arsenic free environment must posses a mechanism necessary to resist other toxic levels of arsenic.
PDF Abstract XML References Citation
How to cite this article
M.A. Salam, M.S. Hossain, M.E. Ali, M.A. Asad and M.H. Ali, 2009. Isolation and Characterization of Arsenic Resistant Bacteria from Different Environment in South-West Region of Bangladesh. Research Journal of Environmental Sciences, 3: 110-115.
DOI: 10.3923/rjes.2009.110.115
URL: https://scialert.net/abstract/?doi=rjes.2009.110.115
DOI: 10.3923/rjes.2009.110.115
URL: https://scialert.net/abstract/?doi=rjes.2009.110.115
INTRODUCTION
Arsenic is the 20th most abundant element in the Earth`s crust 14th in the seawater and 12th in the human body (Woolson, 1975) and is widely distributed throughout nature as a result of weathering, dissolution, fire, volcanic activity and anthropogenic input (Cullen and Reimer, 1989). The last includes the use of arsenic in pesticides, herbicides, wood preservatives and dye stuffs as well as production of arsenic-containing wastes during smelting and mining operations. In arsenic-enriched environments, a major concern is the potential for mobilization and transport of this toxic element to ground water and drinking water supplies. In Bangladesh, an estimated 57 million people have been exposed to arsenic through contaminated wells. This incident serves as an unfortunate reminder of the toxic consequences of arsenic mobilization and underscores the need to understand the factors controlling the mobility and solubility of arsenic in aquatic systems (Newman et al., 1997). The primary anthropogenic input derives from combustion of municipal solid waste, fossil fuels in coal and oil-fired power plants, release from metal smelters and direct use of arsenic-containing herbicides by industry and agriculture. There are a number of ways by which the human population can become exposed to arsenic. The most important one is probably through ingestion of arsenic in drinking water or food. Toxicity and detoxification of heavy metal and transition metal oxyanions in living organisms are tightly bound to membrane transport systems of ions and oxyanions. Due to its un-ionized form at neutral pH, arsenite can passively move across the membrane bilayer or be transported by a carrier protein similar to those that transport un-ionized organic compounds. Arsenate poisoning generally results from the transport of this ion by the phosphate transport system thereby competitively inhibiting the oxidative phosphorylation pathway. One phosphate transport system (Pit) takes up both, phosphate and arsenate, at similar rates, whereas the other (Pst, phosphate specific transport) is highly specific for phosphate. Bacteria defective in the Pit pathway (Pit) are usually arsenate resistant (Cervantes et al., 1994) and for Pit-bacteria the oxidation of As (III) to As (V) represents a potential detoxification process allowing them to tolerate high levels of arsenate. A number of microorganism had been isolated that could use Arsenic in their metabolism, either using arsenate as a terminal electron acceptor in an aerobic respiration (Ahmann et al., 1994; Stolz and Orenland, 1999) or as a means of generating energy through chemoautotrophic arsenite oxidation (Santini et al., 2000). Bacteria might show resistance to an arsenite through the activity of arsenite oxidase (Muller et al., 2003). More recently, a chemolithoautotrophic arseniteoxidizing bacterium belonging to the proteobacteria has been isolated from a gold mine (Santini et al., 2000).
The plasmid determined arsenic resistance system has always had the same biochemical mechanisms, reduced uptake due to an ATPase efflux system in both gram-negative and gram-positive bacteria. Most studies of arsenic resistant bacteria have been conducted in environments that contain high concentrations of arsenic. However, research shows that common microorganisms such as Escherichia coli, Pseudomonas aeruginosa and Staphylococcus aureus, also exhibit arsenic resistance. This goal of this research study was to isolate bacterial from arsenic contaminated environments that contain mechanisms of resisting arsenic and begin characterization and identification of these isolates. We hypothesized that isolated bacteria are highly resistant to arsenic, so they could represent good candidates for bioremediation processes of native polluted sediments. Considering the aspects mentioned in the present research program has been undertaken the following objectives:
| • | To isolate bacterial species, which are resistant to arsenic from arsenic polluted environment |
| • | To characterize the putative arsenic resistant bacteria |
MATERIALS AND METHODS
The experiment was conducted through August 2007 to April 2008 at Microbiology Laboratory of Biotechnology and Genetic Engineering Discipline, Khulna University, Bangladesh to study soil samples were collected from different arsenic contaminated sites of Khulna shipyard (Dry soil near the ship), Bagmara (Dry soil side of the river, Wet soil near the tube well, Wet soil near the tube pond), Ramnagar (Wet soil near the tube well, Wet soil near the pond), Rupsha, Khulna in sterile polyethylene bags. Soil samples were best collected from top to 20 cm. Every time of collection hands were sterile with alcohol (95%) and then the bags were partially tilled with materials and labeled. Special care was always taken to avoid contamination of arsenic after collection the samples were brought to the laboratory carefully and put it in the freeze for immediate use. For long time preservation the sample will be storage at 4°C. Soil dilution technique is used for the isolation of arsenic resistance bacteria. At first 34 different bacterial colonies were isolated from the soil samples by culturing them in Nutrient Agar (NA) medium containing 5 ppm Arsenic. From this 6 bacterial strains were finally selected for detail study. The strains were inoculated in NA slant using a sterile loop and incubated at 37 °C for 18-27 h. After growth the slants were stored at 4 °C for short term storage and for uses. For long term preservation, bacterial growth was harvested from culture slant by sterile inoculating loop. Dense cell suspensions were prepared into 10% (v/v) aqueous glycerol in one vial and stored at 15 °C. The selected bacterial strains were morphological characterized through non-microscopic such as agar colony, agar slant, broth culture and microscopic fixed stained smear such as gram stain and spore stain modified method and biochemical characterized with the help of Bergey`s Manual (Buchanan and Gibson, 1984).
Growth of the Strains in Different Concentrations of Arsenic (As3+)
Tubes of nutrient broth containing different concentration of arsenic (0, 1, 5, 10, 20, 50 and 100 ppm) were equally inoculated with fresh culture and incubated at 37 °C. After 24 h, growth on nutrient broth measured by spectrophotometric reader (600 nm).
Growth at Different Temperature
Tubes of nutrient broth were equally inoculated with fresh culture and incubated at 4, 20, 37 and 60 °C. After 24 h, growth on nutrient broth measured by spectrophotometric reader (600 nm) in four times.
Growth at Different pH
Tubes of nutrient broth at different pH (4.5, 6.5 and 8.5) were prepared in duplicates and after inoculation incubated at 37 °C. After 24 h, growth on nutrient broth measured by spectrophotometric reader (600 nm) in four times.
RESULTS
Morphological Studies
Selected strains were microscopically studied. Vegetative cells, spores and gram reaction were observed under microscope and the results were shown in Table 1. From the observation there are 5 strains were gram positive and 1 strain is gram negative. In flagella reaction 4 strains were positive and 2 strains were negative.
From these morphological studies indicate that the strain number 1/10, 2/9, 3/6, 4/9, 5/8 and 6/8 may be Bacillus lichefomis, Listeria murrayi, Bacillus polymyxa, Moraxella urethralis, Planococcus citreus and Pseudomonas fluorescens.
Biochemical Characterization
Selected strains were studied for their physiological and biochemical tests which were needed for characterization of the strains.
From these biochemical studies shows that the strain number 1/10, 2/9, 3/6, 4/9, 5/8 and 6/8 determine Bacillus lichefomis, Listeria murrayi, Bacillus polymyxa, Moraxella urethralis, Planococcus citreus and Pseudomonas fluorescens (Table 2).
Growth Responses of the Selected Strains at Different Temperature
For the determination of optimum temperature, strains were grown in nutrient broth medium. Tubes of NB were equally inoculated with inocula and incubated at different temperature 4, 25, 37 and 60 °C. In both cases, growth on nutrient broth was measured by spectrophometric reading at 600 nm after 24 h. Table 3 indicated that 37 °C are suitable for bacterial growth and then optical density is 8.7.
| Table 1: | Morphological studies and staining properties of the selected bacterial strains |
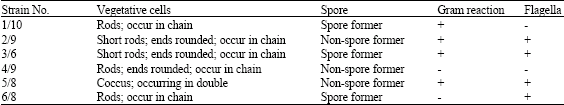 | |
| +: Indicates positive result, -: Indicates negative result | |
| Table 2: | Some biochemical characterization of selected strains |
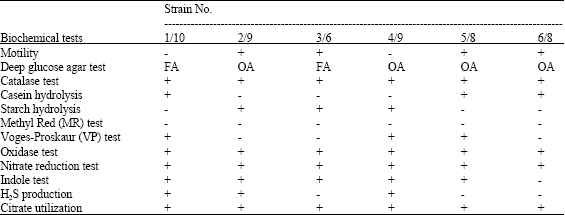 | |
| +: Indicates positive result, -: Indicates negative result, FA: Facultative anaerobe, OB: Obligate aerobe | |
| Table 3: | Growth responses of the selected strains at different temperature |
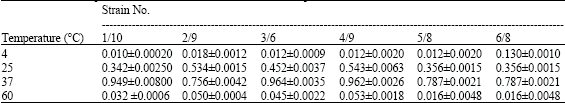 | |
| Table 4: | Growth responses of the selected strains at different pH |
 | |
| Growth responses of the selected strains at different concentrations of Arsenic (As3+) | |
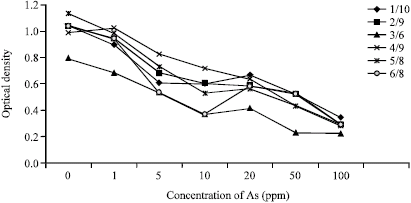 | |
| Fig. 1: | Growth responses of all strains at different concentrations of arsenic |
Growth Response of Different pH
Results were taken in spectrophotometer at 600 nm. Growth response of bacterial strains depends on different pH. Table 4 indicated that 8.5 were suitable for bacterial growth.
Organisms were studied in different concentrations of Arsenic (As3+) containing culture media e.g., nutrient broth with Arsenic (As3+) 0, 1, 5, 10, 20, 50 and 100 ppm (mg L-1) in spectrophotometer at 600 nm. Cultural characteristics of the selected isolates were shown in Fig. 1.
DISCUSSION
Anderson and Cook (2004) reported 17 morphologically distinct arsenic resistant heterotrophic bacteria to be members of the genera Exigeobacterium, Aeromonas, Bacillus, Pseudomonas, Escherichia and Acinetobacter. Macur et al. (2001) also found that members of the Caulobacter, Sphingomonas and Rhizobium may be responsible for the reduction and mobilization of arsenic. Hoeft et al. (2002) found that sulfurospirillium and Desulfovibrio use arsenate as an electron acceptor for their growth. This study reveals that the three genera such as Listeria, Moraxella and Planococcus are also resistant to arsenic. The diversity of arsenic resistance gene is probably much greater and more complex than is apparent from studies on known arsenic resistant isolates (Jackson and Dugas, 2003). Heavy metal toxicities and binding are pH dependent (Wood, 1983) and it appear that the strains require environmentally relevant pH for growth. Bouchard et al. (1996), Christiansen and Ahring (1996) and Niggemeyer et al. (2001) reported optimum pH 7.5 for arsenic resistant strains Desulfitobacterium frappieri, D. hafniense and Desulfitobateriun strain GBFH, respectively.
Temperature is another important environmental factor, which affects bacterial growth (Herbert and Bhakoo, 1979). The optimum temperature (37°C) for the growth of all the strains in the present study support the results observed by Bouchard et al. (1996), Christiansen et al. (1996) and Niggemeyer et al. (2001).
Jackson et al. (2005) isolated numbers of culturable arsenate (V) resistant bacteria from which some were capable to tolerate very high (100 mM) level of arsenate, although arsenic resistance was generally much lower. In addition, Zelibor et al. (1987) isolated As (V)r bacteria in well water samples. These isolates tolerated up to 2,000 pg of As (V) per mL. However, they did not test for As (III) resistance. Honschopp et al. (1996) isolated an arsenic resistant and arsenic methylating bacterium belonging to the Flavobacterium-Cytophaga group, which was able to tolerate 200 ppm concentration of As in the culture media. Plasmids also have been detected in some bacteria exhibiting high level of resistance to arsenate, arsenite and antimonate (Cervantes et al., 1994; Dabbs and Sole, 1988; Mobley et al., 1983).
The bioremediation of arsenic from contaminated sites involves reduction and oxidation of arsenic with the use of arsenic resistant microorganisms.
The successful exploitation of these bacterial strains with proper biotechnology for bioremediation of arsenic will be beneficial. Therefore, more advance research is required for a deeper understanding about these bacterial strains to improve arsenic bioremediation process.
REFERENCES
- Anderson, C.R. and G.M. Cook, 2004. Isolation and characterization of arsenate-reducing bacteria from arsenic-contaminated sites in New Zealand. Curr. Microbiol., 48: 341-347.
CrossRefPubMedDirect Link - Bouchard, B., R. Beaudet, R. Villemur, G. McSween and F. Lepine et al., 1996. Isolation and characterization of Desulfitobacterium frappieri sp. nov., an anaerobic bacterium which reductively dechlorinates pentachlorophenol to 3-chlorophenol. Int. J. Syst. Bacteriol., 46: 1010-1015.
Direct Link - Cervantes, C.G.J., J.L. Ramairez and S. Silver, 1994. Resistance to arsenic compounds in microorganisms. FEMS Microbiol. Rev., 15: 355-367.
PubMed - Christiansen, N. and B.K. Ahring, 1996. Desulfitobacterium hafniense sp. nov., an anaerobic, reductively dechlorinating bacterium. Int. J. Syst. Bacteriol., 46: 442-448.
Direct Link - Cullen, W.R. and K.J. Reimer, 1989. Arsenic speciation in the environment. Chem. Rev., 89: 713-764.
CrossRefDirect Link - Dabbs, E.R. and G.J. Sole, 1988. Plasmid-borne resistance to arsenate, arsenite, cadmium and chloramphenicol in a Rhodococcus species. Mol. Gen. Genet., 211: 148-154.
Direct Link - Hoeft, S.E., F. Lucas, J.T. Hollibaugh and R.S. Oremland, 2002. Characterization of microbial reduction in the anoxic bottom waters of Mono Lake, California. Geomicrobiol. J., 19: 23-40.
CrossRef - Honschopp, S., N. Brunken, A. Nehrhorn and H.J. Breunig, 1996. Isolation and characterization of a new arsenic methylating bacterium from soil. Microbiol. Res., 151: 37-41.
PubMed - Macur, R.E., J.T. Wheeler and T.R. Mcdermott, 2001. Microbial populations associated with the reduction and enhaned mobilization of arsenic in mine tailings. Environ. Sci. Technol., 35: 3676-3682.
Direct Link - Mobley, H.L., C.M. Chen, S. Silver and B.P. Rosen, 1983. Cloning and expression of R-factor mediated arsenate resistance in Escherichia coli. Mol. Gen. Genet., 191: 421-426.
PubMed - Muller, D., D. Lievremont, D.D. Simeonova, J.C. Hubert and M.C. Lett, 2003. Arsenite oxidase aox genes from a metal-resistant b-Proteobacterium. J. Bacteriology, 185: 135-141.
Direct Link - Newman, D.K., T.J. Beveridge and F.M.M. Morel, 1997. Precipitation of arsenic trisulfide by Desulfotomaculum auripigmentum. Applied Environ. Microbiol., 63: 2022-2028.
Direct Link - Niggemeyer, A., S. Spring, E. Stackebrandt and R.F. Rosenzweig, 2001. Isolation and characterization of a novel As(V)-reducing bacterium: Implications for arsenic mobilization and the genus Desulfitobacterium. Applied Environ. Microbiol., 67: 5568-5580.
PubMedDirect Link - Santini, J.M., L.I. Sly, R.D. Schnagl and J.M. Macy, 2000. A new chemolithoautotrophic arsenite-oxidizing bacterium isolated from a gold mine: Phylogenetic, physiological and preliminary biochemical studies. Applied Environ. Microbiol., 66: 92-97.
Direct Link - Stolz, J.F. and R.S. Oremland, 1999. Bacterial respiration of arsenic and selenium. FEMS Microbiol. Rev., 23: 615-627.
PubMed - Wood, J.M., 1983. Selected biochemical reactions of environmental significance. Chem. Scripta., 21: 155-159.
Direct Link - Woolson, E.A., 1975. The persistence and chemical distribution of aesanilic acid in the soils. J. Food Chem., 23: 677-681.
Direct Link - Zelibor, J.L.J., M.W. Doughten, D.J. Grimes and R.R. Colwell, 1987. Testing for bacterial resistance to arsenic in monitoring well water by the direct viable counting method. Applied Environ. Microbiol., 53: 2929-2934.
PubMed
Dr. Sachin P Bachate Reply
This paper would have presented in better way, Use of poor english makes the article difficult to understand. Discussion has no relevance with the results obtained. I just wonder how is it possible to identify the bacteria on the basis of morphology??