
Shireen A. Jahan Quazi
Institute of Tropical Agriculture, Universiti Putra Malaysia, 43400 Serdang, Selangor, Malaysia
LiveDNA: 880.31808
Sariah Meon
Department of Plant Protection, Universiti Putra Malaysia, 43400 Serdang, Selangor, Malaysia
Hawa Jaafar
Department of Crop Science, Universiti Putra Malaysia, 43400 Serdang, Selangor, Malaysia
Zainal Abidin B.M. Ahmad
Department of Plant Protection, Universiti Putra Malaysia, 43400 Serdang, Selangor, Malaysia
Research Journal of Botany
Year: 2020 | Volume: 15 | Issue: 1 | Page No.: 6-15







ABSTRACT
Background and Objectives: Bakanae is one of the major fungal diseases of rice. It is difficult to develop bakanae resistant varieties due to high genetic variation of the pathogen. Hence, bakanae is alarming in the affected rice growing areas of the world. It is essential to find out the infection process of the causal pathogen in rice plant to manage this notorious disease. Therefore, screening was done first to categorize the susceptible and resistant variety and then infection process of the causal pathogen was studied. Materials and Methods: Selected nine varieties were screened against bakanae disease susceptibility. Using the disease severity scale of 0-5, three disease scores of 1, 3 and 5 were found to produce typical bakanae symptoms in the susceptible variety MR 211 after 7, 14 and 21 days of inoculation, respectively. Inoculated varieties were categorized as resistant, moderately resistant and susceptible irrespective of DSI and combined effect of GA3, FA, MON and chitinase activity. Among nine varieties, seven were identified as susceptible, one variety (G-27) as moderately resistant and one variety (BR3) as resistant. Results: It was observed that initiation of infection by F. proliferatum started after 24 h of inoculation and infection became established in the susceptible variety MR 211 after 5 days, while colonization was observed after 7 days. Thus, approximately 5-7 days for infection establishment and colonization were found suitable for symptoms expression. This infection was established through the seed coat of the susceptible variety MR 211. Conclusion: Knowing the duration of infection process and disease development of F. proliferatum in susceptible rice varieties, it will be helpful to take appropriate bakanae management option through disruption of its infection cycle.
PDF Abstract XML References Citation
How to cite this article
Shireen A. Jahan Quazi, Sariah Meon, Hawa Jaafar and Zainal Abidin B.M. Ahmad, 2020. Varietal Screening and Infection Process of Fusarium proliferatum in Rice Varieties. Research Journal of Botany, 15: 6-15.
URL: https://scialert.net/abstract/?doi=rjb.2020.6.15
URL: https://scialert.net/abstract/?doi=rjb.2020.6.15
INTRODUCTION
The bakanae disease of rice caused by Fusarium moniliforme was identified in Japan in a broad sense1. This taxon comprises of one or more Fusarium species that are now collectively termed as Gibberella fujikuroi species complex, as the disease is caused by one or more Fusarium species. A total of five Fusarium F. spp. belonging to “Gibberella fujikuroi species complex” under section Liseola (F. fujikuroi, F. proliferatum, F. sacchari, F. subglutinans and F. verticillioides) have been isolated and found to be associated with bakanae disease in Malaysian rice varieties2,3. Fusarium fujikuroi has also been reported as pathogenic2,4. Among the species complex, Fusarium proliferatum is also identified as a causal agent of bakanae disease of rice5.
The disease symptoms are expressed at seedling stage first and finally the infected seedlings are turned to death. Therefore, control measures should be taken before spreading infection inside seedlings. Disease control can be done in many ways, either by eliminating pathogen inoculum or by limiting disease spread or by escaping from pathogen or by using disease resistant material. To manage bakanae infection following any of the control measure stated above is depended on pathogen biology and mode of infection process. It has also been reported that bakanae symptom can be expressed after 5-14 days of artificially inoculation2,4. But, the detail infection process for bakanae disease development by any of the causal Fusarium species is yet to be published. It is essential to know the infection process by the causal pathogen before practice any control method of bakanae disease. Moreover, a limited number of resistant materials against bakanae disease have been identified and reported6.
Plant pathogens have been reported to be proficient in the synthesis of the 3 phytohormones Gibberellic acid (GA3), Indole Acetic Acid (IAA) and Abscisic acid (ABA)7-11. In addition, it was observed that pathogens can alter metabolic activity by producing different phytohormones in host plant cells and facilitate infection in plants12. The relationship among production of different phytohormones, quantity and plant resistance have already been established13. A number of plant diseases have been identified that are correlated with phytohormone level changes in plants. Fusarium species are often found to produce a wide range of metabolites including, fumonisins, moniliformin, beauvericin, fusaric acid, fusaproliferin, enniatin and trichothecenes that are toxic and can contaminate human and animal food14. Both F. fujikuroi and F. proliferatum have been found to produce similar metabolites. Metabolites produced by plant pathogens were mainly found to responsible for wilting, chlorosis and necrosis by the interruption of normal physiological functions in plants rather than disease development15. These metabolites may be produced either before penetration into the host or after the infection process has been established inside plants. In addition, a wide variety of fungal metabolites have been identified as both mycotoxic (toxic to animals) as well as phytotoxic (toxic to plants). It has also been established that many mycotoxin-producing fungal species cause plant disease under field conditions16. Hence, mycotoxins produced by the fungus in plants are becoming of greater concern to mycologists along with animal toxicologists and human disease epidemiologists. Pathogenesis Related (PR) proteins β-1, 3 glucanase and chitinase are found to be implicated in defense responses of plants against pathogens. Phytohormones, metabolites and pathogenesis related proteins might also have an impact on bakanae disease development. Therefore, this experiment was aimed to identify the susceptible and resistant varieties in terms of infection process observation under Scanning Electron Microscope (SEM) and the infection process of Fusarium proliferatum for bakanae disease development in susceptible variety vs. resistant variety.
MATERIALS AND METHODS
The experiments were carried out at greenhouse and laboratory of Institute of Tropical Agriculture (ITA), Universiti Putra Malaysia, Malaysia in 2011-2013.
Varietal screening of selected rice varieties: Varietal screening was performed on 9 selected commercial rice varieties. Three rice varieties, namely BR1, BR3 and Germplasm (G)-27 were obtained from Germplasm and Genetic Resources and Seed Division (GRSD), Bangladesh Rice Research Institute (BRRI), Bangladesh. Another 6 varieties: M-4, MR185, MR211, MR 219, MR 220 and MR 232 were collected from the Malaysian Agricultural Research and Development Institute (MARDI) in January, 2011.
Inoculation method: For varietal screening “seed inoculation” method was used for pathogenicity test and the virulent isolate of Fusarium proliferatum was used for inoculation5. In brief, 14 days old F. proliferatum isolates cultured on PDA were flooded with sterile water (100 mL) and scraped with a sterile spatula separately. The resulting suspensions were pooled and filtered through two layers of sterile muslin cloth and the final concentration was adjusted to 1×106 conidia/mL by adding sterile distilled water and used for inoculation. Seed of each variety was surface sterilized separately with 70% ethanol washed with sterilized distilled water and then soaked in sterilized distilled water overnight. The water was drained out and seeds were further soaked in conidial suspension of the virulent isolate (106 conidia/mL) for 48 h. Seeds for the control treatment was soaked in sterile distilled water for 72 h before sowing.
Planting and growth management: The inoculated seeds were then planted in sterilized soil separately (40% sand, 30% clay, 30% peat) on trays (2 kg soil/tray) measuring 38×28×11 cm arranged in a completely randomized design with three replications (20 seeds/replication). All trays were maintained in a glasshouse at room temperature and watered daily. Fertilizer comprising of N: P: K (15:15:15) were applied at 4 g/tray, twice at 15 day intervals.
Data collection and scoring for resistance/susceptibility reaction: The disease symptoms development was recorded at different intervals after inoculation from seedling to booting stage. Disease Severity Index (DSI) over time was calculated by observations of degree of infection progress in different varieties based on disease severity5 scale 0-5. Random re-isolation of the causal pathogen from the diseased plant parts was carried out for fulfillment of Koch’s postulate. Inoculated varieties were categorized irrespective of DSI in the range of <0.2 between 0.2-1.0 and >1.0 designated as resistant, moderately resistant and susceptible.
Relationship study between phytohormone, metabolites and PR-proteins in terms of disease resistance/susceptibility: Phytohormone (GA3), metabolites (FA, MON) were isolated and quantified by following Quazi et al.17,18. PR-protein (chitinase) was isolated and quantified according to Quazi19. Combine effect of GA3, FA, MON and chitinase activity were analyzed and a dendrogram was constructed through multi variate analysis by using PAST software (Version 3.0).
Sample preparation for Scanning Electron Microscope (SEM) analysis: Whole seeds (1, 2 and 5 days after inoculation) and tissues from seed sections (7 days after inoculation) were fixed in 4% glutaraldehyde for 2 days at 4°C. After fixation, the samples were washed with sodium cacodylate buffer 3 times for 30 min each, post fixed in 1% Osmium Tetra Oxide for 2 h at 4°C and subsequently washed again with 0.1 M sodium cacodylate buffer three times for 30 min each. Dehydration of the fixed samples was carried out at room temperature in a series of acetone concentrations of 35, 50, 75, 95 and 100%, each for 30 min. The dehydrated samples were then placed in the specimen basket and dried in a critical point dryer (LEICA EM CPD030) for 30 min. Dried samples were then mounted on to the stub by using double sided tape, gold coated for 2 min in a Sputter-Coater (BALTEC SCD 005) and examined directly under the SEM (Jeol, JSM 6400). Each sample was processed and viewed thrice for confirmation. Three sets of samples of each treatment were examined under SEM.
Observation of infection processes of Fusarium proliferatum under Scanning Electron Microscope (SEM): Infection process of F. proliferatum was studied in three different varieties: MR 211 (susceptible), BR3 (resistant) and G-27 (moderately resistant) artificially inoculated with a virulent isolate of F. proliferatum (106 conidia/mL). Inoculated and un-inoculated (control) samples of susceptible, resistant and moderately resistant varieties were observed under Scanning Electron Microscope (SEM) at 1, 2, 5 and 7 days after inoculation. This time frame for observation of infection process was designed as the disease symptom caused by F. proliferatum was observed after 5-7 days of inoculation5.
Statistical analysis: Means of plant height increase in seed and soil inoculation method irrespective of isolate and in varietal screening irrespective of variety and time was compared by using SAS 9.2 software.
RESULTS
Susceptibility of selected rice varieties to Fusarium proliferatum: First disease symptom was appeared on the highly susceptible seedlings (BR1and MR 211) after 5 days of inoculation. Seedling stem height was higher 2-5 times compared to control plants over time. Infected susceptible plants were observed as thinner with pale green to yellowish lower leaves which turned onto brown and dried-up with time. Adventitious roots were found to be produced on the upper 2nd-3rd nodes in some plants, while some plants became stunted and showed necrosis on lower internodes. At later stage of infection, white fungal mycelial masses were produced on infected plant parts at water level, which turned pinkish with time. Root necrosis was also associated with bakanae diseased plants. Variety BR3 categorized as resistant only showed symptoms that appeared as yellowish leaves at late growth stage, with no further development of bakanae. Symptoms associated with bakanae disease illustrates in Fig. 1.
 | |
| Fig. 1(a-h): | Different types of symptoms produced from bakanae diseased plants, (a, c) Stem height elongation of bakanae diseased plant compared to control plant, (a, b) Stunted bakanae diseased plant compared to control plant, (d) Green leaves turned into brown colored and finally turned to death, (e) Root formation on upper 2nd and 3rd nodes, (f) Fungal masses reduced on lower parts of plants, (g) Necrotic lesions on lower internodes and (h) Crown rot and root necrosis in bakanae diseased plants |
Rice varieties inoculated with F. proliferatum were categorized as susceptible; moderately resistant and resistant based on Disease Severity Scale (DSS) and Disease Severity Index (DSI) (Table 1). Seven varieties (BR1, MR 185, MR 211, MR 219, MR 220, MR 232 and M 4) were identified as susceptible in terms of higher mean DSI values ranging from 2.38-3.77. Although, there was no significant difference observed between BR3 and G-27 varieties in terms of mean DSI after the 9th week, but DSI was observed to be higher in G-27 compared to BR3 from week 5-9. Moreover, symptoms related to chlorosis were observed to be higher in G-27, whereas BR3 produced only mild symptoms with yellowing of lower leaves at later plant growth stage. Therefore, BR3 was categorized as resistant and G-27 as moderately resistant. The DSI increased with time for all the varieties and significant differences (p = 0.01%) in interaction between variety and time were also observed (Table 2).
Combine effect of GA3, FA, MON and chitinase activity: The combined effect of GA3, FA, MON and chitinase activity were higher for disease development and symptoms expression in infected susceptible variety MR 211. In contrast, combined effect was minimal in infected resistant variety BR3 at different disease score levels (Fig. 2).
| Table 1: | Disease categorization of the selected rice varieties (Disease category was based on DSI as follows: Resistant (<0.2), moderately resistant (between 0.2-1.0) and susceptible (>1.0) |
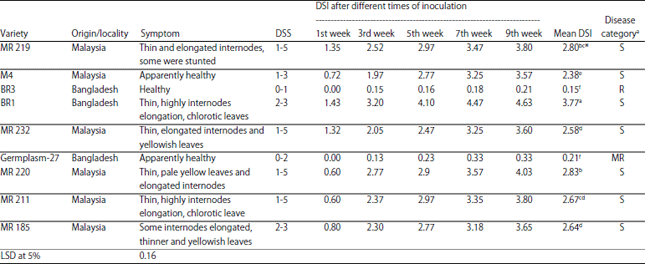 | |
*Means tagged with different letters within column are significantly different using LSD at p≤0.05, aS: Susceptible, MR: Moderate resistant, R: Resistant | |
| Table 2: | Varietal screening |
 | |
*Interaction of Disease Severity Index (DSI) of different varieties with time (DSI were calculated at 1st, 3rd, 5th, 7th and 9th weeks after inoculation) | |
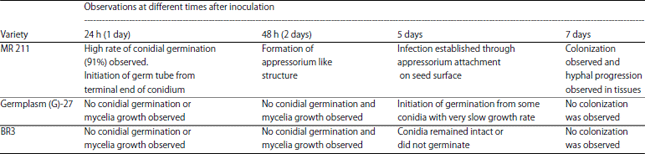
| Table 3: | Summary of infection processes of Fusarium proliferatum between varieties at different times of inoculation |
 | |
Further, it was noted that this combined effect was higher in disease score level 5 followed by disease score level 1 and disease score level 3. The lesser effect at disease score level 3 in susceptible variety was due to the lower amount of FA whereas, a higher amount of FA was associated with disease score 1 and 5, respectively.
Dendrogram of disease score levels in relation to disease development: In the Dendrogram of different disease score levels in relation to disease development and symptoms expression, three clusters were observed among the varieties MR 211 and BR3 at distance 0.35 (Fig. 3).
Two clusters were observed in susceptible variety MR 211. Disease score level 3 of MR 211 was separated from other cluster and this disease score level 3 expressed the typical bakanae symptom (thin, pale green leaves and elongated internodes). Other clusters belonging to disease score level 1 and disease score level 5 expressed stunting and growth cessation symptoms with browning in susceptible variety MR 211, respectively. All 3 scores belonging to infected resistant variety BR3 were observed as a separate and individual cluster. The cluster belonging BR3 is also separated from the other 2 clusters of infected susceptible variety MR 211 and did not express any bakanae symptom.
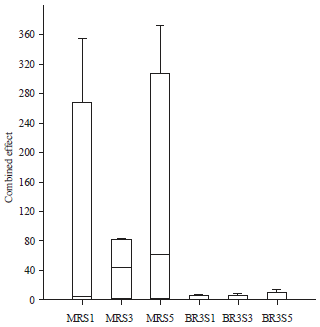 | |
| Fig. 2: | Combined effect of GA3, FA, MON and chitinase activity in relation to disease development and symptoms expression in infected susceptible variety MR 211 and in infected resistant variety BR3 |
MR S1: Disease score level 1 of MR 211 (S-1), MRS3: Disease score level 3 of MR 211(S-3), MRS5: Disease score level 5 of MR 211 (S-3), BRS1: Disease score level 1 of BR3 (S-1), BRS3 = BRS1: Disease score level 3 of BR3 (S-3), BRS5: Disease score level 5 of BR3 (S-5), vertical bar presents standard error | |
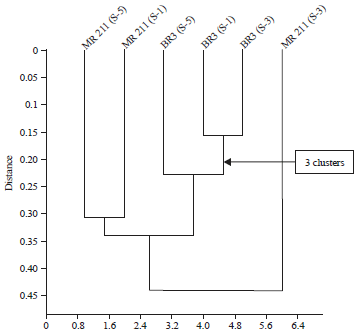 | |
| Fig. 3: | Disease score level similarities in relation to disease development and symptoms expression in infected susceptible variety MR 211 and in infected resistant variety BR3 |
S-1: Disease score level 1, S-3: Disease score level 3, S-5: Disease score level 5 | |
Infection processes of Fusarium proliferatum: The infection progress of F. proliferatum was observed on susceptible (MR 211), moderately resistant (G-27) and resistant (BR3) varieties and summarized in Table 3 and Fig. 4-6. Germination of conidia was significantly higher on the seed surface of susceptible variety MR 211 (91%) after 24 h of inoculation. Germination was initiated by postulation of germ tubes (Fig. 4a). Appressorium like structures were observed after 48 h of inoculation (Fig. 4b). The primary infection process was found to be established through the seed coat after 5 day of inoculation on susceptible variety MR 211 (Fig. 4c). Colonization and hyphal progress in tissues were observed in susceptible variety MR 211 after 7 days of inoculation (Fig. 4d, e).
A few numbers of conidia were appeared to show initiation of germination in moderately resistant variety G-27. But, this poor germination of conidia did not take part in infection establishment and colonization in tissues after 5 days of inoculation (Fig. 5a, b). On the other hand, the conidia remained intact in the resistant variety BR3 or did not germinate after 5 days of inoculation (Fig. 6a). Infection progress or colonization was not observed in the tissues of BR3 after 7 days of inoculation (Fig. 6b).
DISCUSSION
Plant pathogen infection causes disruption of normal physiological processes by using a mixture of biochemical factors in plants. These biochemical factors include cell wall degrading enzymes/PR-proteins, phytohormones and toxic compounds/metabolites20,21. Although, the role of phytohormones in relation to cause diseases in plants are not clearly understood yet, but it has been reported that there is a relationship between production of different phytohormones quantity and plant resistance12. Other researcher had also shown possible scenarios on different phytohormones production in plants after pathogen infection22. By alteration of these different phytohormones level in plants also have ability to change the expression of defense related genes and thereby possess activation of defense responses by showing disease susceptibility or resistance.
Relationship studies between symptoms expression and involvement of phytohormone and metabolites produced by F. proliferatum in infected plants were found. Polynomial relationships of GA3 and FA were associated with disease symptoms expression in disease score 1 and 3 in susceptible variety. Higher amount of GA3 and lower amount of FA in disease score 3 are responsible for plants height increase.
 | |
| Fig. 4(a-e): | Infection processes of Fusarium proliferatum in susceptible variety MR 211, (a) Initiation of germ tube after 24 h of inoculation, (b) Formation of appressorium like structure after 48 h of inoculation, (c) Infection established through seed coat after 5 days of inoculation, (d) Colonization and (e) Hyphal progression, in tissues after 7 days of inoculation |
 | |
| Fig. 5(a-b): | Infection process of Fusarium proliferatum in the moderately resistant variety Germplasm (G)-27, (a) Very few conidia were observed to initiate germination after 5 days of inoculation on seed surface and (b) No colonization was observed after 7 days of inoculation in tissues |
Conversely, this association was vice versa in disease score 1 that resulted in stunting of plants. Moreover, polynomial relationship for FA and logarithmic relationship for MON was found in association with different disease score levels. However, higher amounts of FA and MON produced by F. proliferatum in infected plants at disease score 5, might be responsible for a decrease in the amount of GA3. Hence, decrease of root length, ceased growth, plant death and necrosis of roots were observed in susceptible variety MR 211.
 | |
| Fig. 6(a-b): | Infection process of Fusarium proliferatum in the resistant variety BR3, (a) Conidia remained intact after 5 days of inoculation on seed surface and (b) No colonization was observed in tissues after 7 days of inoculation |
It is assumed that FA and MON act synergistically, whereas both metabolites act antagonistically with GA3 in bakanae diseased plants. Thus, different symptoms were expressed depending on the combination of GA3, FA and MON at different disease score levels. Moreover, disease score level 3 had been identified as a completely different disease score level in relation to symptoms expression. This might be due to the higher amount of GA3 associated with lower amounts of metabolites FA and MON. Quite the opposite, symptoms at disease score level 1 and 5 were associated with higher amounts of metabolites FA and MON rather than GA3. In terms of the combined effect of phytohormone, metabolites and chitinase activity, variety MR 211 and BR3 were clearly differentiated on the characteristics responsible for making the plants susceptible/resistant.
Fusarium proliferatum is a seed and soil borne pathogen and can infect rice plants through the seed coat. Similar results were observed by other researcher who noted that the infecting hyphae of F. proliferatum can penetrate the epidermal cell wall directly23. Fusarium proliferatum has been shown to produce appressorium after 48 h of inoculation on the seed surface. However, it did not produce typical well-defined appressorium like other fungi. Usually, the germ tube tips grow first into the host cell surface and then swell slightly and after that it infects the host by developing a constricted penetration peg. Similar findings were reported by Zhao et al.23. It was observed that at least 5 days were required for establishment of the pathogen on to the seed surface. It was also reported that the subsequent 3 days after inoculation were found to be vulnerable for seed infection2. On contrary, it was reported that the bakanae causing pathogen Gibberella fujikuroi can infect plants either through the roots or crowns24.
There was chlorotic symptoms development along with some conidial germination on the G-27 seed surface. No other bakanae symptoms associated with conidial germination on the G-27 seed surface is termed as moderate resistant variety. On the other hand, no conidial germination on the seed surface of BR3 having no bakanae symptom production termed as resistant variety. Initiation of poor mycelial growth of the pathogen on G-27 and inhibition of conidial germination on seed surface of BR3 might be due to different chemical composition of these varieties compared to the susceptible variety MR 211. There might be a disruption of the FPK1 gene in F. proliferatum when inoculated into the resistant variety (BR3) and the moderately resistant variety (G-27). Similar observations were supported by other researcher and reported that spore germination, hyphal growth and pathogenicity of F. proliferatum is influenced by this FPK1 gene22. Alternatively, antioxidants might be produced in the resistant variety BR3 and moderately resistant variety G-27 and resulted symptomless infection. A similar explanation was provided that pathogenicity is correlated with the presence of pectic enzymes of the host and virulence as well as the infection process of the pathogen25. But all the assumptions might be more applicable to the resistant variety (BR3) compared to moderately resistant and resistant variety. Therefore, spore germination was hindered in BR3 and little mycelial growth occurred in G-27.
The present study has established the infection processes of F. proliferatum for bakanae disease development through different symptoms expression. However, the physiological factors affecting symptoms expression in the susceptible variety (MR 211) in comparison to the resistant variety (BR3) was not clearly understood. Therefore, further studies are needed to clarify the symptoms expression of bakanae disease by the pathogen F. proliferatum in susceptible as well as in resistant varieties.
CONCLUSION
This is the first report on identification of the detail infection process of F. proliferatum in rice plants. In addition, bakanae disease infection process is progressed either through seed coat or through young roots, respectively. From the findings of this research, it is evident that F. proliferatum germinate after 24 h of inoculation followed by appressorium formation. At least 5 days are required for pathogen establishment, then penetration and colonization produce symptoms in susceptible variety MR 211. Thus, 5-7 days are required for disease development and symptoms expression. The infection and establishment period of F. proliferatum for bakanae disease development is identified.
SIGNIFICANCE STATEMENT
This study discovers the infection process of F. proliferatum in rice that can be beneficial for management of bakanae disease. This infection and establishment period of F. proliferatum is crucial to take appropriate management option to control the disease. Thus, it is important to take management approach at infection initiation stage to disrupt the pathogen’s infection cycle before establishment and symptom expression in infected plants of rice. This study will help the researcher to uncover the critical areas of infection establishment period in rice caused by F. proliferatum that many researchers were not able to explore. Thus, a new theory on infection process of F. proliferatum may be arrived at.
ACKNOWLEDGMENTS
The authors wish to express their sincere thanks and appreciations to Universiti Putra Malaysia (UPM) for the research grant administered through RUGS (01-04-10-1040-RU), the Bangladesh Agricultural Research Council (BARC) through a Doctoral fellowship and the Institute of Tropical Agriculture and the Faculty of Agriculture, Universiti Putra Malaysia for providing the research facilities. The authors also express special thanks to the Technical Editor and Head of Publications and Public Relations Division, Bangladesh Rice Research Institute for assist English language proofreading service to prepare this manuscript.
REFERENCES
- Izzati, M.Z.N.A. and B. Salleh, 2009. Genetic variability amongst Fusarium spp. in the section Liseola from bakanae-infected rice in Malaysia and Indonesia by RAPD analysis. Malaysian Applied Biol. J., 38: 71-79.
Direct Link - Zainudin, N.I.M., A. Razak and B. Salleh, 2008. Bakanae disease of rice in Malaysia and Indonesia: Etiology of the causal agent based on morphological, physiological and pathogenicity characteristics. J. Plant Protect. Res., 48: 475-485.
CrossRefDirect Link - Amatulli, M.T., D. Spadaro, M.L. Gullino and A. Garibaldi, 2010. Molecular identification of Fusarium spp. associated with bakanae disease of rice in Italy and assessment of their pathogenicity. Plant Pathol., 59: 839-844.
CrossRefDirect Link - Quazi, S.A.J., S. Meon, H. Jaafar and Z.A.B.M. Ahmad, 2013. Characterization of Fusarium proliferatum through species specific primers and its virulence on rice seeds. Int. J. Agric. Biol., 15: 649-656.
Direct Link - Qureshi, O., H. Sohail, A. Latos and J.L. Strap, 2013. The effect of phytohormones on the growth, cellulose production and pellicle properties of Gluconacetobacter xylinus ATCC 53582. Acetic Acid Bacteria, 2: 39-46.
CrossRefDirect Link - Khan, A.L., M. Hamayun, S.M. Kang, Y.H. Kim, H.Y. Jung, J.H. Lee and I.J. Lee, 2012. Endophytic fungal association via gibberellins and indole acetic acid can improve plant growth under abiotic stress: An example of Paecilomyces formosus LHL10. BMC Microbiol., Vol. 12, No. 1.
CrossRefDirect Link - Kilaru, A., B.A. Bailey and K.H. Hasenstein, 2007. Moniliophthora perniciosa produces hormones and alters endogenous auxin and salicylic acid in infected cocoa leaves. FEMS Microbiol. Lett., 274: 238-244.
CrossRefDirect Link - Patten, C.L. and B.R. Glick, 2002. Role of Pseudomonas putida indoleacetic acid in development of the host plant root system. Applied Environ. Microbiol., 68: 3795-3801.
CrossRefPubMedDirect Link - Robert-Seilaniantz, A., L. Navarro, R. Bari and J.D.G. Jones, 2007. Pathological hormone imbalances. Curr. Opin. Plant Biol., 10: 372-379.
CrossRefPubMedDirect Link - Husaini, A.M., A. Sakina and S.R. Cambay, 2018. Host-pathogen interaction in Fusarium oxysporum infections: Where do we stand? Mol. Plant-Microbe Interact., 31: 889-898.
CrossRefDirect Link - Desjardins, A.E. and T.M. Hohn, 1997. Mycotoxins in plant pathogenesis. Mol. Plant-Microbe Interact., 10: 147-152.
CrossRefDirect Link - Quazi, S.A.J., S. Meon, H. Jaafar and Z.A.B.M. Ahmad, 2015. The role of phytohormones in relation to bakanae disease development and symptoms expression. Physiol. Mol. Plant Pathol., 90: 27-38.
CrossRefDirect Link - Quazi, S.A.J., M. Sariah, Z.A.B.M. Ahmad and J. Hawa, 2016. Detection of fungal metabolites from bakanae diseased plants and their relationship with bakanae disease symptoms expression. Am. J. Biosci. Bioeng., 4: 77-89.
CrossRefDirect Link - Kazan, K. and R. Lyons, 2014. Intervention of phytohormone pathways by pathogen effectors. Plant Cell, 26: 2285-2309.
CrossRefDirect Link - Lebeda, A., D. Jancova and L. Luhova, 1999. Enzymes in fungal plant pathogenesis. Phyton, 39: 51-56.
Direct Link - Bari, R. and J.D.G. Jones, 2009. Role of plant hormones in plant defence responses. Plant Mol. Biol., 69: 473-488.
CrossRefDirect Link - Zhao, P.B., A.Z. Ren and D.C. Li, 2011. The FUS3/KSS1-type MAP kinase gene FPK1 is involved in hyphal growth, conidiation and plant infection of Fusarium proliferatum. J. Mol. Microbiol. Biotechnol., 21: 110-119.
CrossRefPubMedDirect Link - Kikot, G.E., R.A. Hours and T.M. Alconada, 2009. Contribution of cell wall degrading enzymes to pathogenesis of Fusarium graminearum: A review. J. Basic Microbiol., 49: 231-241.
CrossRefDirect Link