
Alicia Enriqueta Brechu-Franco
Departamento de Ecolog�a y Recursos Naturales, Facultad de Ciencias, Universidad Nacional Aut�noma de, Mexico, Avenida Universidad 3000, Colonia Universidad Nacional Aut�noma de Mexico, C.U., 04510, Coyoac�n, Mexico
LiveDNA: 52.24364
Yazmin Jurado-Gonzalez
Departamento de Ecolog�a y Recursos Naturales, Facultad de Ciencias, Universidad Nacional Aut�noma de, Mexico, Avenida Universidad 3000, Colonia Universidad Nacional Aut�noma de Mexico, C.U., 04510, Coyoac�n, Mexico
Guillermo Laguna-Hernandez
Departamento de Ecolog�a y Recursos Naturales, Facultad de Ciencias, Universidad Nacional Aut�noma de, Mexico, Avenida Universidad 3000, Colonia Universidad Nacional Aut�noma de Mexico, C.U., 04510, Coyoac�n, Mexico
Armando Gomez-Campos
Departamento de Ecolog�a y Recursos Naturales, Facultad de Ciencias, Universidad Nacional Aut�noma de, Mexico, Avenida Universidad 3000, Colonia Universidad Nacional Aut�noma de Mexico, C.U., 04510, Coyoac�n, Mexico
Rosa Maria Fonseca-Juarez
Departamento de Biolog�a Comparada, Facultad de Ciencias, Universidad Nacional Aut�noma de Mexico, Avenida Universidad 3000, Colonia Universidad Nacional Aut�noma de Mexico, C.U., 04510 Coyoac�n, Mexico
Research Journal of Botany
Year: 2018 | Volume: 13 | Issue: 1 | Page No.: 11-18







ABSTRACT
Background and Objective: Moussonia deppeana (Gesneriaceae) is one of the 25 species of the Moussonia genus living in montane cloud forests (MCF) and on the edges of pine-oak forests. It is used in traditional Mexican medicine. Pharmacological studies have shown its antioxidant, anti-inflammatory and cytotoxic properties. Because this plant is currently collected without regard to sustainable management, cultivation under controlled and standardized sanitary regulations should be recommended. Thus, this study assessed the germination capacity and short-term seedling survival of the species. Materials and Methods: Mature and dry fruits of M. deppeana were collected and photographed. A number of seeds from three fruits was calculated as well as their weight and size. Because a preliminary germination test revealed dependence of light, the germination of freshly harvested (FH) seeds and seeds stored for 15 and 13 days at 24±2°C (15S and 30S) under light was then measured. Seedlings that were approximately 1 cm in length 8 weeks after sowing were transplanted into a peat moss substrate and were incubated under controlled conditions for 21 additional weeks, when their survival was evaluated. Results: Germination in FH and 30S seeds reached approximately 80% after the 4th week. The 15S seeds showed germination close to 90% and above from the 2nd week. The seedling survival percentages were 71 and 61% in FH and 15S seeds respectively and 36% for 30S seeds. Conclusion: The abundant production of tiny seeds, more than 3,300 with 0.5 mm×0.2 mm in size and 20 μg in weight, besides high germination under light, could indicate that M. deppeana is a pioneer species, similar to other species from the Kohleria genus (Gesneriaceae). Propagation of this species in a standardized culture program would be successful under light using FH and 15S seeds owing to its high viability and vigor.
PDF Abstract XML References Citation
How to cite this article
Alicia Enriqueta Brechu-Franco, Yazmin Jurado-Gonzalez, Guillermo Laguna-Hernandez, Armando Gomez-Campos and Rosa Maria Fonseca-Juarez, 2018. Germination Capacity and Seedling Survival of Moussonia deppeana (Gesneriaceae) from the Montane Cloud Forests. Research Journal of Botany, 13: 11-18.
URL: https://scialert.net/abstract/?doi=rjb.2018.11.18
URL: https://scialert.net/abstract/?doi=rjb.2018.11.18
INTRODUCTION
The plant Moussonia deppeana or "tlachichinole" is a species with medicinal properties with high demand for its use, whose collection is done by extracting plants from the natural populations and has no reports under sustainable management.
Moussonia deppeana (Schltdl. and Cham.) Hanst., Gesneriaceae Dumort.1 is a perennial shrub or subshrub species that reaches 1.5 m in height and occurs in humid and semi-warm areas in montane cloud forests (MCFs) and gallery forests as well as in areas close to streams and canyons and on the edges of pine-oak forests (POFs). Its distribution ranges from north-central Mexico to western Panama. Moussonia deppeana is the most widely distributed species of this genus and is used for medicinal purposes in various parts of Mexico. In Mexico, this species is found in the states of Veracruz, Puebla, Oaxaca, Guerrero, Morelos, Hidalgo and Chiapas1-3. Moussonia deppeana was collected from an area of the Sierra Madre del Sur, Guerrero, Mexico, at 2000 masl on a rocky, shaded, very humid mountain wall within the MCF. The dominant tree species are Ostrya virginiana, Oreopanax xalapensis, O. sanderianus, Clethra galeottiana, Bernardia fonsecae, Quercus spp. and Pinus ayacahuite and abundant species of pteridophytes and epiphytic Araceae also occur. The presence of some Urticaceae indicates some degree of disturbance, likely due to the proximity to a dirt road. Flowering was recorded from August-December and fruiting was recorded from October to April4.
Gesneriaceae is a mainly tropical family, with 147 genera and approximately 3,460 species. Most species are herbs, but they are sometimes shrubs and rarely trees, lianas and epiphytes occurring in moist sites and they contain distinctive verbascosides (e.g., sanangoside), 3-deoxyanthocyanins, chalcones, aurones, caffeoyl phenylethanoid glycoside and phenolic glycosides. Many species are currently cultivated for ornamental purposes, the best known example being Saintpaulia ionantha or African violet, an indoor plant of multiple varieties; other genera containing ornamental species are Episcia, Columnea, Sinningia, Streptocarpus and Aeschynanthus. Gesneriaceae species often produce capsular fruits with small or minute seeds5 that are dispersed by the wind or by the action of raindrops that strike the fruit and cause the seeds to be released from the capsule in a type of dispersal known as "splash-cup dispersal", a dispersal mechanism that also acts on Marchantia's conceptacles and the crown-shaped fruits of Sagina decumbens and Oenothera rosea6,7. The Central American genus Moussonia was placed in Kohleria until recently due to their similar tubular flowers and inflorescences. Moussonia differs from Kohleria by having a basic chromosome number of 11 rather than 13, a capitate stigma and an annular nectary8,9. The number of species in this genus varies according to the authors who have studied it, with 18 accepted names (http://www.tropicos.org/) and possibly 24 species in total and 19 species in Mexico1.
The morphological characteristics of the seeds in the Kohleria genus of the Gesneriaceae family were previously assessed and it was reported that light plays an important role in several developmental phases in Streptocarpus rexii, such as seed germination and seedling development, which are related to adaptation to dense forest habitats9; however, Aud and Ferraz10 indicated that photoblastism and dependency on temperature alternation are not characteristics shared by all pioneer species, because some pioneer species can germinate in diffuse light under a dense canopy or in the dark and even at a constant temperature11,12. In epiphyte species in the same forest type (bromeliads), these conditions are important both in terms of germination and seedling survival13. The negative effect of edges on germination and adult occurrence in Gesneriaceae is modulated along a vertical gradient (height), with contrasting effects on seed germination and survival to adulthood14,15.
Although the Gesneriaceae are usually appreciated as ornamental plants, some of them also have medicinal uses, such as M. deppeana, which is known as tlachichinole or clachichinole in Mexico, where it has been used against gastrointestinal conditions, cough, stomach and/or duodenal ulcers and renal and vaginal infections and for blood purification and boosting energy16-20. Evaluations of the actions of organic extracts of M. deppeana against eleven strains of mycobacteria suggested that M. deppeana biosynthesizes compounds with antimycobacterial activity21; in addition, the anti-inflammatory and antioxidant properties of M. deppeana were endorsed by Dominguez-Ortiz et al.22 and it exhibits anti-proliferative and cytotoxic activities against the LNCaP prostate cancer cell line23.
Moussonia deppeana is considered one of the most commercially important plants in Mexico and other countries, especially the USA, for the preparation of several herbal remedies24. However, the commercial supply of M. deppeana depends exclusively on the extraction of the plant from its natural habitat and there are no current studies on the sustainable management of M. deppeana in relation to its capacity for forest regeneration and productivity. Thus, the germination capacity of the seeds should be identified to enable the development of a standardized cultivation program for this species. The study of the reproductive characteristics of M. deppeana should be framed within the objectives of Resolutions I and II of the XVI International Congress of Botany-1999 since this is a medicinal plant species, whose habitat in the MCF in Mexico has been highly degraded. This study was performed to characterize the germination of M. deppeana and its short-term seedling survival under controlled environmental conditions as a starting point for the preservation of the species. The results established the possibility to begin a cultivation program for this species.
MATERIALS AND METHODS
Seed collection: On February 22, 2010, mature M. deppeana plants bearing fruits were collected from MCF vegetation, adjacent to a road (17°35ʹN-99°47ʹ994" W) at 2,000 masl, in "El Puente", Leonardo Bravo district, Guerrero state, Mexico. The herborized samples of M. deppeana were deposited at Herbario de la Facultad de Ciencias (FCME) of Universidad Nacional Autonoma de Mexico, under reference voucher #4935.
Seed procurement: The capsular fruits were photographed after collection, using a Canon® E OS Rebel T6i 24 MP. Three ripe and dry fruits were opened in a Petri dish to extract the seeds under a Zeiss® Stemi SV6 stereoscopic microscope. The seeds from each fruit were weighed and the average weight of the three fruits was obtained; subsequently, the number of seeds in each fruit was obtained by counting them manually using a magnifying glass and the average was calculated. The relationship between the average number of seeds and their average weight corresponded to the average weight of each seed. Atrophied seeds, characterized by a collapsed or thin and smaller state, named empty seeds, were observed and their percentage was calculated. The seeds were photographed with the same camera attached to the microscope (Carl-Zeiss-Promenade 10, 07745 Jena, Germany). The seeds were measured using an Axio Vision System (Carl-Zeiss-Promenade 10, 07745 Jena, Germany).
Conditions for germination: Five sets of 50 seeds each per treatment were sown in 10 cm diameter Petri dishes with absorbent paper soaked with 10 mL of 0.2% Captan fungicide. The plates were sealed with Parafilm and stored at 24±2°C in Lab-Line Biotronette environmental chambers maintained at 24±2°C with a 12 h light/day cycle and a relative humidity of approximately 65-75%. The experiment was performed in a randomized design. Light was controlled using cool white fluorescent lamps with a very high intensity of 500 mM m–2 sec.
Seed germination: On March 11, 2010, freshly harvested (FH) seeds were separated into two groups: (1) Immediate germination test seeds and (2) Storage germination tests seeds.
A preliminary germination test to determine the light dependence of the seeds was performed from February 23 to March 9, 2010, with some sets of seeds placed under light conditions and others in darkness. Because the results showed that the seeds require light for germination, all the tests were carried out under light conditions.
Immediate germination test: Five sets of 50 seeds each were maintained under light within the incubator from March 11, 2010. The cumulative germination (%), calculated based on the number of seeds that germinated (radicle emergence or cotyledon stage) was obtained every two weeks for 2 months (from 2-8 weeks) until May 6, 2010.
Germination of stored seeds: Freshly harvested seeds were hermetically stored in sealed jars covered with aluminium foil and kept inside an incubator at 24±2°C for 15 or 30 days (from March 11-25, 2010 and from March 11 to April 8, 2010, respectively).
The evaluation of the germination of stored seeds under exposure to light proceeded according to immediate germination test after the storage period (from March 25 to May 20 and from April 8 to June 3, 2010, respectively).
Comparison of the germination of freshly harvested and stored seeds: The effect of seed storage on germination was analyzed through one-way ANOVA (Statgraphics Centurion XVI) with a p-level of 0.0017, using a randomized experimental design with a single factor (seed condition or seed age: freshly harvested or stored seeds) and a block representing the time to germination (2, 4, 6 and 8 weeks after sowing). The treatments took the age of the seeds into account: 1 = freshly harvested seeds (FH), 2 = seeds stored for 15 days (15S) and 3 = seeds stored for 30 days (30S) and the response was the cumulative germination (%)25.
Then, to verify which treatments differed, the means from the individual treatments were analyzed using Student’s t-test25, comparing two groups at once.
Seedling survival: During the 8th week after the seeds were sown, seedlings greater than or equal to approximately 1 cm in length were measured and then transplanted from the Petri dishes to transparent boxes that were 12 cm in width×12 cm in length×6 cm in height and contained a moist peat moss substrate and were then maintained in the incubator under the same conditions. The survival rates were evaluated 21 weeks later, when seedlings greater than or equal to 4 cm in length were recorded. Transplanted seedlings from FH seeds were incubated from May 6-September 30; those from 15S were kept in incubation from May 20-October 14 and those from 30S were kept in incubation from June 3-October 28, 2010.
Comparative survival of the transplanted seedlings: A means test with three populations (seedlings that grew from FH seeds and those that grew from 15S and 30S seeds) and an one-way ANOVA (Statgraphics Centurion XVI) with a p-level of 0.000001, with fixed effects were used for a single treatment for each measurement time (8th and 21st weeks after the seeds were sown)25.
RESULTS
Moussonia deppeana has flowers arranged in axillary umbels, which produce bivalve, hairy, dry, dehiscent capsules fruits (Fig. 1a, b) with numerous tiny seeds that have an ellipsoid shape 0.5 mm in length×0.2 mm in width and feature longitudinal striations on the seed coat (Fig. 1c). Each fruit contained an average of 3,300 seeds, with each one weighing approximately 20 μg and the average percentage of empty seeds was 13%. Germination of M. deppeana seeds resulted in epigeous seedlings with above-ground cotyledons (Fig. 1d).
Immediate germination: The seeds exposed to light began to germinate from the 2nd week after sowing, with 66.4% germination, reaching a total percentage of 80% by the 8th week.
Germination of stored seeds: Almost 90% of the seeds stored for 15 days germinated by the second week, increasing to 92.8% towards the eighth week; the germination of the seeds stored for 30 days only reached 66.4% by the second week and 81.2% by the eighth week (Fig. 2).
 | |
| Fig. 1(a-d): | Moussonia deppeana, (a) Shrub with inflorescence, (b) Dehiscent fruit, (c) Ellipsoidal seeds with longitudinal striations and (d) Epigeous seedling |
| Co: Cotyledon, Hy: Hypocotyl, Sc: Seed coat | |
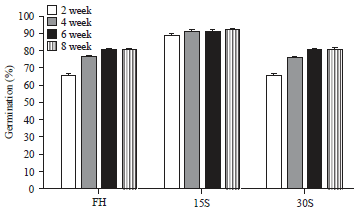 | |
| Fig. 2: | Cumulative germination percentages of M. deppeana seeds at 2nd, 4th, 6th and 8th weeks after planting |
| FH freshly harvested seeds, 15S and 30S seeds stored for 15 or 30 days at 24±2°C | |
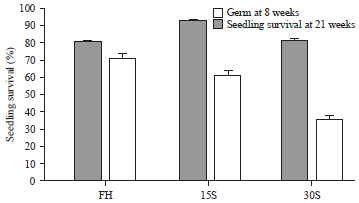 | |
| Fig. 3: | Survival of seedlings from the freshly harvested (FH) seeds and the seeds stored for 15 and 30 days (15S and 30S) at 24±2°C by the 21st week after transplanting |
The statistical analysis comparing the germination of FH, 15S and 30S seeds during the 8th week after sowing showed significant differences, with the highest value in the 15S seeds, followed by the FH seeds and finally the 30S seeds (F = 11.4, p = 0.0017) (Fig. 2).
Seedling survival: The seedlings grew slowly, reaching a length of 1.0 cm by approximately the 8th week after planting in the Petri dishes, the time at which the seedlings were transferred to the transparent plastic boxes. At 21st weeks after transplanting, the seedlings had reached 4.0 cm in length and their survival (%) were 71% for FH seeds, 61% for 15S seeds and 36% for 30S seeds. The statistical analysis comparing the survival of seedlings from the FH, 15 and 30S seeds by the 21st week after transplanting to the transparent boxes with peat moss showed significant differences (F = 47.01, p = 0.000001), with the highest value in FH seeds, followed by 15S seeds and finally 30S seeds (Fig. 3).
Overall survival: The number of seedlings surviving in the 8th week after sowing was 627 and the number of surviving seedlings at 21st weeks was 407, which represents 65% survival.
DISCUSSION
The results obtained in this study correspond to important characteristics indicated for some species of Gesneriaceae, such as Streptocarpus rexii. Each fruit of M. deppeana produced 3,000-4,000 tiny seeds. The seeds have been qualitatively described in the literature as tiny or small for Gesneriaceae, but their dimensions have not been mentioned5,2,26,27. In this work, was determined the seed’ morphology in addition to their size and weight and the average number per fruit. Moussonia deppeana, also known as Kohleria deppeana (synonym), produces brown ellipsoidal seeds with a longitudinally striated seed coat. These characteristics are common to 11 of the 17 species of the Kohleria genus9. The species with this type of seeds are specifically found in exposed or unstable environments, e.g., along rivers and highways, similar to the habitat in which M. deppeana was collected. The MCF vegetation in this area occurs on a stone wall with humid soil covered by herbs, shrubs and trees and clearings bordering open paths. Seeds of M. deppeana germinated only under white light conditions and created an epigeal seedling28. Therefore, this species displays a light requirement or light-sensitive latency29, which is similar to the seeds of Streptocarpus rexii, another species in Gesneriaceae, which show dependency on light27.
Seed germination was defined here as the emergence of the hypocotyl through the seed coat and had a homogeneous response, with high percentages of between 80-90% reached from the 2nd week to the 8th week after sowing. The immediate and simultaneous germination syndrome, a common strategy in plants of the humid and warm forests of tropical regions has been reported to reduce seed mortality in the soil, which is due to the deleterious action of physical and biotic agents30-35. Results showed that the germination of FH seeds and 30S seeds reached approximately 80%, but those stored for 15 days attained a higher germination (%). The difference in germination between FH seeds and 15S seeds was possibly due to embryo immaturity and the difference in germination between 15S seeds and 30S seeds could be due to deterioration as a result of storage. However, the survival of the seedlings that were transplanted and kept in soil for up to 20 weeks was lower for the 30S seeds than the 15S seeds. This could indicated that the seeds of M. deppeana exhibit the immediate germination syndrome but that storage for 30 days diminished the viability of the seeds and the vigor of seedlings. These characteristics could be important for the colonization of MCF edges by this species. Slow seedling development did not influence survival, so it could be proposed that natural development depends on the presence of light, which occurs in Streptocarpus rexii, in which light plays an important role at different stages of development and this mechanism could be associated with the adaptability to dense forest habitats where light levels are low27.
Moreover, due to the small size of the seed, M. deppeana has limited nutritional reserves for seedling development5. In addition, the light requirement limits the options for seedling development and light is one of the necessary conditions for germination. The production of a considerable quantity of seeds per fruit is another strategy for ensuring successful dispersal and development under certain conditions, as established by Munoz-Garcia et al.36 in their study of pioneer tree species. Similar results were reported in the case of three epiphytic gesneriads of temperate Chilean rainforests, which present different germination responses depending on the light environment and substrate15. The existence of 13% empty seeds represents a small number among the thousands of potentially successful seeds for the propagation of the species due to the characteristics of germination, the development of the seedlings and survival, with low percentages of decrease among seedlings from FH seeds (9%) compared with the large difference in the seedlings from 15S (19%) and 30S (44%) seeds, which may indicated that survival in the forest depends on the germination of the newly scattered seeds under the appropriate conditions of light, humidity and temperature. The humid montane forest in Mexico is likely the most threatened ecosystem37,38 and is the one that occupies the lowest amount of surface area worldwide39,40.
It is estimated that less than 1% of the Mexican territory is occupied by primary MCF vegetation (8,809 km2) (http://www.inegi.org.mx/) and approximately 50% of the original area has been replaced by other types of cover38,41. In addition, mesophilic forests are affected by extensive livestock farming and, to a lesser extent, by urban growth and the logging of timber and extraction non-timber products42. Therefore, the preservation of the MCF is a priority due to its exceptional and unique composition of flora and species richness. It provides environmental services such as carbon sequestration, landslide and flood erosion prevention and stream water quality preservation43,44. Seedling growth and survival are critical for tropical rainforest restoration and studies of the propagation of species of MCFs therefore contribute to the understanding of the regeneration process, which is necessary for the development of protected area management strategies and serves as a basis for conservation and restoration practices.
Moussonia deppeana presents some characteristics of pioneer species, such as the abundance and small size of seeds and their dependence on light, which also occurs in Amazon species with small seeds10,45.
Moussonia deppeana may plays an important ecological role as a pioneer species on the edge of this type of vegetation and its propagation by seed reinforces its genetic variability12. Furthermore, M. deppeana is considered to be one of the most commercially important plants in Mexico and other countries, especially the USA, for the preparation of several herbal remedies24, but it is not included as a species at risk of extinction in the Mexican Legislation for Environmental Protection46. The aforementioned properties make this species suitable for cultivation in plots, family-owned gardens and large-scale greenhouses among the owners of forest resources. These advantages indicated that M. deppeana is a resource with the potential for large-scale commercialization and bringing M. deppeana into cultivation would allow for the continuous supply of chemically standardized plant material47,48 that is safe for direct consumption or for use as a raw material for the production of phytomedicines and nutraceuticals. The conservation and restoration of MCFs in forest landscapes necessitate the importance of considering the particular conditions of edges9. In this way, the diverse uses of the forest would be promoted, adding to its value and the forest populations of M. deppeana would be protected against intensive extraction.
CONCLUSION
This study showed that Moussonia deppeana has a high production of tiny seeds with a valuable viability and a considerable survival of the seedlings, similar to a pioneer species. It was observed that the seeds had a better response of germination and survival of the seedlings when they are freshly harvested seeds and with 15 days of storage. It was also found that they are seeds that depend on the presence of light to germinate.
SIGNIFICANCE STATEMENT
This work studied the sexual propagation of the medicinal plant Moussonia deppeana or "tlachichinole" and the survival of the seedlings in the early stages of development. Therefore, knowing the germination process and the survival response would allow a basis for the cultivation of the species and development of the plants in order to obtain the resource without the extraction from their natural environment.
ACKNOWLEDGMENTS
Authors thanks the following academic staff of the UNAM Faculty of Sciences (Facultad de Ciencias): Patricia Olguín Santos and María Eugenia Muñiz Díaz de León, for technical support during seedling incubation, Ana Isabel Bieler Antolin and Alfonso Bautista García for image processing, Argelia Díaz Rico for support in the bibliographic search and María del Pilar Alonso Reyes for the review of the statistical data.
REFERENCES
- Santiago-Perez, A.L., E.J. Jardel-Pelaez, R. Cuevas-Guzman and F.M. Huerta-Martinez, 2009. Border vegetation in a cloud forest in Western Mexico. Bol. Soc. Bot. Mexico, 85: 31-49.
Direct Link - Brodie, H.J., 1951. The splash-cup dispersal mechanism in plants. Can. J. Bot., 29: 224-234.
CrossRefDirect Link - Ertelt, J., 2013. Observations on splash seed dispersal among neotropical gesneriaceae. Selbyana, 3: 234-239.
Direct Link - Wiehler, H., 1975. The re-establishment of Moussonia regel (Gesneriaceae). Selbyana, 1: 22-31.
Direct Link - Kvist, L.P. and L.E. Skog, 1992. Revision of Kohleria (Gesneriaceae). Smithsonian Contrib. Bot., 79: 1-83.
CrossRefDirect Link - Aud, F.F. and I.D.K. Ferraz, 2012. Seed size influence on germination responses to light and temperature of seven pioneer tree species from the central Amazon. An. Acad. Bras. Cienc., 84: 759-766.
CrossRefDirect Link - Kyereh, B., M.D. Swaine and J. Thompson, 1999. Effect of light on the germination of forest trees in Ghana. J. Ecol., 87: 772-783.
CrossRefDirect Link - Pearson, T.R.H., D.F.R.P. Burslem, C.E. Mullins and J.W. Dalling, 2002. Germination ecology of neotropical pioneers: Interacting effects of environmental conditions and seed size. Ecology, 83: 2798-2807.
CrossRefDirect Link - Winkler, M., K. Hulber and P. Hietz, 2005. Effect of canopy position on germination and seedling survival of epiphytic bromeliads in a Mexican humid montane forest. Ann. Bot., 95: 1039-1047.
CrossRefDirect Link - Magrach, A., L. Santamaria and A.R. Larrinaga, 2013. Edge effects in a three-dimensional world: Height in the canopy modulates edge effects on the epiphyte Sarmienta repens (Gesneriaceae). Plant Ecol., 214: 965-973.
CrossRefDirect Link - Salinas, F. and J.J. Armesto, 2012. Regeneration niche of three epiphytic species of gesneriaceae from Chilean rainforests: implications for the evolution of growth habits in coronanthereae. Bot. J. Linean Soc., 170: 79-92.
CrossRefDirect Link - Nieto, M.A.V. and B.E.P. Escandon, 2010. Vegetacion e inventario de la flora util de la huasteca y la zona otomi-tepehua de hidalgo. Cienc. Univ., 1: 23-33.
Direct Link - Jimenez-Arellanes, M.A., L. Yepez-Mulia, J. Luna-Herrera, G.A. Gutierrez-Rebolledo and R.V. Garcia-Rodriguez, 2013. Antimycobacterial and antiprotozoal activities from Moussonia deppeana (Schldl and Cham) Hanst. Rev. Mex. Cienc. Farm., 44: 31-35.
Direct Link - Dominguez-Ortiz, M.A., O. Munoz-Muniz, R.V. Garcia-Rodriguez, M. Vazquez-Hernandez, J. Gallegos-Estudillo and J.S. Cruz-Sanchez, 2010. Antioxidant and anti-inflammatory activity of Moussonia deppeana. Boletin Latinoamericano y del Caribe de Plantas Medicinales y Aromaticas, 9: 13-19.
Direct Link - Nishii, K., T. Nagata, C.N. Wang and M. Moller, 2012. Light as environmental regulator for germination and macrocotyledon development in Streptocarpus rexii (Gesneriaceae). S. Afr. J. Bot., 81: 50-60.
CrossRefDirect Link - Baskin, C.C. and J.M. Baskin, 2014. Seeds: Ecology, Biogeography and Evolution of Dormancy and Germination. 2nd Edn., Elsevier, USA., ISBN-13: 9780124166837, pp: 1166.
Direct Link - Macedo, M., 1977. Dispersao de plantas lenhosas de uma Campina Amazonica. Acta Amaz., 7: 5-69.
CrossRefDirect Link - Ng, F.S.P., 1980. Germination ecology of Malaysian woody plants. Malaysian Forester, 43: 406-438.
Direct Link - Rees, M., 1993. Trade-offs among dispersal strategies in British plants. Nature, 366: 150-152.
CrossRefDirect Link - Ray, G.J. and B.J. Brown, 1994. Seed ecology of woody species in a Caribbean dry forest. Restor. Ecol., 2: 156-163.
CrossRefDirect Link - Rzedowski, J., 1996. Analisis preliminar de la flora vascular de los bosques mesofilos de montana de Mexico. Acta Bot. Mex., 35: 25-44.
CrossRefDirect Link - Sanchez-Ramos, G. and R. Dirzo, 2014. El Bosque Mesofilo de Montana: Un Ecosistema Prioritario Amenazado. In: Bosques Mesofilos de Montana de Mexico: Diversidad, Ecologia y Manejo, Gual-Diaz, M. and A. Rendon-Correa (Eds.). Comision Nacional para el Conocimiento y Uso de la Biodiversidad, Mexico, D.F., ISBN: 978-607-8328-07-9, pp: 352.
- Manson, R.H., 2004. Los servicios hidrologicos y la conservacion de los bosques de Mexico. Madera Bosques, 10: 3-20.
Direct Link - Valio, I.F.M. and F.M. Scarpa, 2001. Germination of seeds of tropical pioneer species under controlled and natural conditions. Rev. Bras. Bot. 24: 79-84.
CrossRefDirect Link - Norma Oficial Mexicana, 2001. Norma Oficial Mexicana NOM-059-ECOL-2001, Proteccion ambiental-Especies nativas de Mexico de flora y fauna silvestres-Categoruias de riesgo y especificaciones para su inclusion, exclusion o cambio-Lista de especies en riesgo. Miercoles 6 de marzo de 2002 DIARIO OFICIAL (Segunda Sección) SEMARNAT, pp: 1-85. http://www.biodiversidad.gob.mx/pdf/NOM-059-ECOL-2001.pdf.
- Sahoo, N., K. Choudhury and P. Manchikanti, 2009. Manufacturing of biodrugs: Need for harmonization in regulatory standards. BioDrugs, 23: 217-229.
CrossRefDirect Link - Sahoo, N., P. Manchikanti and S. Dey, 2010. Herbal drugs: Standards and regulation. Fitoterapia, 81: 462-471.
CrossRefDirect Link