
Taoufik Saleh Ksiksi
Department of Biology, College of Science, United Arab Emirates University, Al Ain, UAE
LiveDNA: 1.19883
Shaijal Babu Thruppoyil
Department of Biology, College of Science, United Arab Emirates University, Al Ain, UAE
LiveDNA: 91.19851
Abdul Rasheed Palakkott
Department of Biology, College of Science, United Arab Emirates University, Al Ain, UAE
Research Journal of Botany
Year: 2017 | Volume: 12 | Issue: 3 | Page No.: 53-62







ABSTRACT
Background and Objective: Elevated carbon dioxide (CO2) has the ability to mitigate plant drought stress. Not many studies were done to find the combined effects of drought stress and elevated CO2 on C4 plant growth under low photosynthetic photon flux density (PPFD). In this study, drought mitigating effects of elevated CO2 were assessed on the vegetative growth of maize (Zea mays L.) plants under low PPFD. Materials and Methods: Maize plants were grown in two growth chambers, one maintained at ambient CO2 (435 μmol mol–1) and the other at elevated CO2 (1000 μmol mol–1), conditions under low PPFD. Plants were subjected to three water stress treatments by reducing irrigation: Once every 7 days (SD, control), every 14 days (FD) and every 21 days (TD). Data were collected on chlorophyll content, stomatal and epidermal characteristics and plant vegetative growth (height and girth). Data were analyzed using SPSS and MANOVA. Results: The elevated CO2 in combination with TD water stress treatment, induced higher chlorophyll ‘a’ (6.95 vs 10.93 μg cm–2), chlorophyll ‘b’ (2.63 vs 3.80 μg cm–2) and total chlorophyll content (14.73 vs 9.38 μg cm–2) compared to TD water stress treatment in ambient CO2. The longest and the widest stomata were recorded under elevated CO2 conditions in the FD (39.40±1.56 cm) and TD (26.63±2.29 cm), respectively. Further, the highest stomatal density was observed in TD under elevated CO2 (151 stomata/mm2). The combination of elevated CO2 and FD water stress treatment registered the tallest (mean = 112.80 cm) and thickest (mean = 2.97 cm) maize plants in the experiment. Conclusion: Elevated CO2 increased vegetative growth of maize plants especially under FD water stress treatment. The low PPFD was not a constraint for elevated CO2 in mitigating the drought stress.
PDF Abstract XML References Citation
Received: September 14, 2017;
Accepted: October 16, 2017;
Published: October 31, 2017
How to cite this article
Taoufik Saleh Ksiksi, Shaijal Babu Thruppoyil and Abdul Rasheed Palakkott, 2017. Elevated CO2 Improves Vegetative Growth of Water-stressed Zea mays L. Under Low Photosynthetic Photon Flux Density. Research Journal of Botany, 12: 53-62.
URL: https://scialert.net/abstract/?doi=rjb.2017.53.62
URL: https://scialert.net/abstract/?doi=rjb.2017.53.62
INTRODUCTION
Maize (Zea mays L.) is a C4 plant and one of the most important food crops in the world in terms of production1. Maize production has been increasing in the last 70 years, due to improved plant breeding methods and cultivation techniques, in major maize producing countries like USA and China2,3. However, recent data show that global climate change has negatively impacted maize production in the world4,5,6. Water stress and increasing atmospheric CO2 have been reported as the major results of the climate change7.
Water stress reduces maize yield up to 30% by negatively impacting photosynthesis8. This is mainly due to the reduced water availability at reproductive stages and restricted leaf growth under water stress2. Non-expanding leaves intercept less photosynthetically active radiation (PAR) and result in reduced net CO2 assimilation rate and photosynthesis5,9. In addition, water stress forces stomata to close, which leads to a fall in CO2 absorption by leaves, which in turn reduces photosynthesis10. Further, water stress reduces plant chlorophyll content either by forcing chlorophyll break down11, hindering chlorophyll synthesis12 or by altering activities of enzymes involved in chlorophyll synthesis13.
Even though water stress reduces CO2 intake by closing stomata, C4 plants already have high bundle sheath CO2 content which is almost 10 times that of atmospheric CO2 content14. Thus, CO2 enrichment is thought not to increase the photosynthesis in C4 plants7. However, exposing C4 plants to double atmospheric CO2 concentration, promoted plant growth and increased CO2 assimilation rate15,16. In contrast to the above observation, some C4 plants were reported not to exhibit any positive growth responses to elevated CO2 conditions17. Leakey et al.7, demonstrated that water stress is an inevitable element for positive response in maize growth under CO2 enrichment.
Like the drought-driven stomatal closure, increased atmospheric CO2 concentration also forces stomatal closure due to localized increase in temperature at the leaf level. Artificial CO2 enrichment acts in the same way by altering leaf microclimate and reduces photosynthesis due to reduction in CO2 uptake10. Apart from the effect on stomata, enriched CO2 environment also affects chlorophyll content in plants. CO2 enrichment can increase18,19, decrease20 or have no affect21 on chlorophyll content, in comparison to ambient CO2 conditions. Previous researches, on the effects of CO2 enrichment and water stress in C4 plants, have reported contrasting results on the positive effect of CO2 enrichment22,23. Earlier studies concentrated on the effects of water stress and CO2 enrichment at reproductive stage. Therefore, it is acceptable to conclude that if vegetative stages are affected by water stress, the crop yield will be reduced.
Based on these previous research findings, the effects of CO2 enrichment during the vegetative growth stages were investigated, starting from two-leaves-stage up to the full-fledged development of 10th leaf. The main objective of this study was to assess the vegetative growth from plant growth traits such as height and stem girth. In addition, total chlorophyll content, length, width and density of stomata, length and width of epidermal cells surrounding stomata, were determined. Overall, this study was aimed to test the feasibility of using CO2 enrichment as tool to mitigate the negative impact of water stress on the vegetative growth of maize plants.
MATERIALS AND METHODS
Plant growth conditions: Hybrid maize seeds (‘Merit’, Seminis) were water soaked for 24 h and sown into bog peat media (Desert group, UAE) filled in 72 well plastic trays and were placed inside growth chambers (Binder KBW-720). There were five ‘Narva cool daylight colourex plus (LT 30W/865) electric tubes’ inside each growth chamber, with an overall light intensity of 63.15±10μmol m–2 sec–1. The irradiance was measured using an LI-189 quantum/Radiometer/Photometer (Li-Cor, Inc., Lincoln, NE, USA). The measurements were done at 2 cm below the tube (on the full fledged10th leaf) and at 50 cm below the lamp (halfway down, on the 6th leaf), to have a look at the effect of light penetration in between the leaves. The irradiance recorded were 28.02 Wm–2 at 2 cm and 9.56 Wm–2 at 50 cm.
Two growth chambers were used, one for ambient CO2 (435μmol mol–1) and the other for enriched CO2 (1000μmol mol–1) treatments. One week old maize seedlings were transplanted into the bog peat media filled in black plastic pots (19.3 cm diameter and 16.4 cm height). The seedlings were transplanted inside each chamber at a spacing of 20×20 cm (between rows and columns). There were 12 such seedlings allotted to each chamber totaling 24 seedlings in the whole experiment. The plants were arranged in a 2 factor completely randomized design (CRD) with multiple levels. The first factor, CO2, had 2 levels (two growth chambers) and the second factor, the water stress treatment, had 3 levels. Each treatment combination (CO2-waterstress) was repeated four times. A photoperiod of 15 h of light and 9 h of darkness was used. The temperature was set at 25°C during the light period and 19°C during the dark period. One week after transplanting maize seedlings, one of the growth chambers was enriched with a CO2 concentration of 1000μmol mol–1 using a CO2 tank (23 kg). The CO2 flow through a 0.4 cm (inner diameter) plastic tube was regulated using a CO2 system (Hydrofarm, CA, USA) and an autopilot digital environmental controller (Hydrofarm, CA, USA). The ambient CO2 concentration (435μmol mol–1) inside the second chamber was attained automatically through a hole on the side of the growth chamber, which equilibrated the CO2 concentration inside the chamber with that of outside air. Both growth chambers were kept at these settings until the completion of the experiment on the 70th day after transplanting. The whole experiment was repeated two times to confirm the results.
Determination of irrigation volume for the experiment: Water stress levels were determined using a pre-experimental set up. For this purpose, 400, 500, 600 and 700 mL of water were added to bog peat potting mix in different plastic pots. Three replications were used for each water treatment. The soil was kept on lab benches at 22°C temperature and 60% relative humidity for 2-3 days. The samples were then oven dried at 105°C for 24 h and the field capacity was calculated. The results from the pre-trial showed that, 600 mL of water was needed for the potting soil mix to reach its field capacity. Based on this value, three different water stress treatments were applied in the pots under ambient and enriched CO2 treatments, which were:
| • | Irrigation to field capacity every 7 days, abbreviated as SD |
| • | Irrigation to field capacity every 14 days, abbreviated as FD |
| • | Irrigation to field capacity every 21 days, abbreviated as TD |
After seed germination, fertilizers were given with the irrigation water to all the plants for the first 14 days. The TUROFORT 20+20+20+2MgO+TE (Adfert, AD, UAE) fertilizer containing micro nutrients was used for the fertigation of maize plants (pH = 6 and Electrical Conductivity = 2.5). After first 14 days, fertilizer application was regulated to once in every 21 days. Non-regulation of fertilizer application could have resulted in faulty measurements of plant growth traits due to the interaction of fertilizer and water stress treatments. Consequently, plants in SD could have been the biggest under both CO2 treatments.
The water stress treatments were applied starting from one week after transplanting the maize seedlings. The SD treatment was used as the control treatment for the water stress treatments, since the plants were irrigated to field capacity in the SD. Maize leaves were detached and immediately processed to determine chlorophyll content, stomatal density, stomatal length, stomatal width, epidermal cell length and width. For the chlorophyll content determination, samples were collected from the 3rd leaf of each maize plant from the bottom due to the long period of exposure to light and CO2 treatments. An additional reason to use the 3rd leaf was that the lower most leaves (leaves 1 and 2) were dried-up when the leaves were collected on the 70th day (‘in-treatment’) for chlorophyll analysis.
Chlorophyll extraction and quantity determination: Chlorophyll content was determined by a method described by Richardson et al.24, with slight modifications. Instead of leaf discs, rectangular pieces (1 cm2 each, 100 mg total fresh weight) were cut from the third leaf counted from the bottom of the maize plant. The cut leaf pieces were immersed into 7 mL of pre-heated DMSO (65°C) contained in a glass vial. After, the glass vials were heated inside a water-bath for 30 min. The volume of DMSO inside the glass vials were then topped up to 10 mL using a Pasteur pipette. The absorbance of each DMSO-extracted chlorophyll sample was determined by a Lambda 25 UV/VIS spectrometer (PerkinElmer, USA). The spectrometer was calibrated to zero absorbance using DMSO in a quartz cuvette as a blank. We used 663 nm, 645 nm wavelengths for chlorophyll determination based on chlorophyll absorption spectra of 600-680 nm reported by Richardson et al.24. Five replications were done for each leaf sample from ambient and enriched drought treatments for both 663 and 645 nm wavelengths. The equations reported by Schlemmer et al.25, were used to convert the absorbance values to chlorophyll ‘a’, chlorophyll ’b’ and total chlorophyll contents. The chlorophyll contents calculated from these equations (g L–1) were then converted to leaf chlorophyll content per unit leaf area (μg cm–2).
Stomatal length, width and density: Stomatal length, width and density were determined from 70 days old maize leaves. Clear nail polish was applied onto the maize leaves and epidermal peels of 6×4 mm were removed from the leaves once the nail polish had dried. The peels were put on a glass slide and pressed with a cover slip. These samples were immediately observed under Olympus BX41 laboratory microscope fitted with a DP 71 microscopic camera (Olympus Corporation, Tokyo, Japan). Micrographs were taken for stomatal density from 200×200μm2 focal area (set using scale option) of the DP 71 camera field set under 40X objective and 10X eyepiece (400X magnification). Micrographs were taken to measure stomatal length, stomatal width and stomatal density. All measurements were done in 3 replications.
Epidermal cell length and width: The epidermal peels observed for stomatal characteristics were also used to determine epidermal cell length and width. Micrographs were taken using Olympus BX41 laboratory microscope fitted with a DP 71 microscopic camera (Olympus Corporation, Tokyo, Japan) from 200×200μm2 focal area of the DP 71 camera at 400X magnification to measure the lengths and widths of the epidermal cells surrounding stomata. All measurements were done in 3 replications.
Plant growth measurements: Maize plant height and stem girth were measured during four different time points to get an idea on the combined effects of water stress and CO2 treatments. The height and girth of maize plants on 15th, 24th, 31st, 47th and 70th days were measured while they were in treatments, referred from here onwards as DIT (days in treatment). The aim was to assess the gradual differences in growth of maize caused by enriched CO2 environment and water stress treatments.
Statistical analysis: Data were analyzed using IBM SPSS Statistics for Windows (IBM Corp., Armonk, NY). All the dependent variables except stomatal density followed a normal distribution and showed homogeneity of variances. Multivariate analysis of variance (MANOVA) GLM procedure was done on all the other dependent variables except for stomatal density (p<0.05). The stomatal density followed Poisson distribution and thus was analyzed using generalized linear model of SPSS. Bonferroni adjustment was used to report the significant pairwise comparisons of the water stress treatments and CO2 levels.
RESULTS
Chlorophyll content: Overall, maize chlorophyll content (μg cm–2) was significantly higher in 1000μmol mol–1 CO2 (enriched CO2) compared to ambient CO2 (p>0.001). Under enriched CO2, chlorophyll ‘a’ was significantly higher in both TD and FD treatments, compared to the ambient CO2 (Fig. 1). The SD (control) also showed higher chlorophyll ‘a’ under CO2 enrichment but it was not high enough to have a significant statistical difference in comparison to ambient CO2 (p>0.05). Similarly, under enriched CO2, Chlorophyll ‘b’ content was higher in the FD and TD treatments compared to ambient CO2, however the difference was not statistically significant (p>0.05). Total chlorophyll content (TCC) followed the similar trend of that of chlorophyll ‘b’ but was significantly higher in both FD and TD under enriched CO2 compared to ambient CO2.
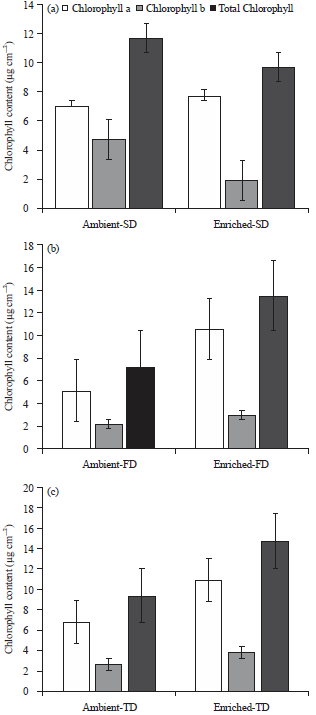 | |
| Fig. 1(a-c): | Chlorophyll ‘a’, chlorophyll ‘b’ and the total chlorophyll content (Mean±SEM) of maize leaves in three water stress treatments (a) SD: 7 days, (b) FD: 14 days and (c) TD: 21 days under ambient (435μmol mol–1) and enriched (1000μmol mol–1) CO2 |
Enriched CO2 actually reduced the TCC in maize plants under the control (SD) treatment.
Stomatal and epidermal characteristics: The highest stomatal length was measured in the FD treatment under enriched CO2 and the lowest in SD, under ambient environment conditions.
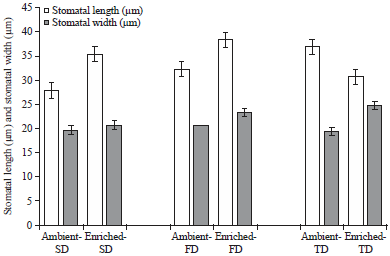 | |
| Fig. 2: | Stomatal length and width (Mean±SEM) of maize leaves in three water stress treatments, SD: 7 days, FD: 14 days and TD: 21 days) under ambient (435μmol mol–1) and enriched (1000μmol mol–1) CO2 |
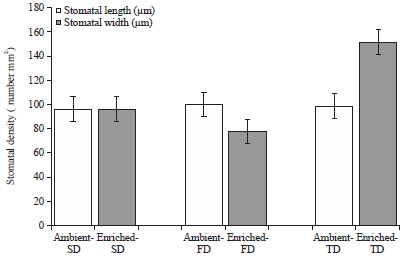 | |
| Fig. 3: | Stomatal density (Mean±SEM) of maize leaves in three water stress treatments (SD: 7 days, FD: 14 days and TD: 21 days) under ambient (435μmol mol–1) and enriched (1000μmol mol–1) CO2 |
The order of increasing stomatal length under ambient environment was SD<FD<TD (Fig. 2). Enriched CO2 significantly increased (p<0.05) the stomatal length in SD and FD treatments but decreased in TD compared to that of ambient CO2. In the enriched CO2, stomatal length increased in the order TD<SD<FD for the water stress treatments.
Stomatal widths were almost similar (p>0.05) among all water stress treatments (SD, FD and TD) under ambient CO2 conditions. A similar trend was also noticed under enriched CO2 environments (Fig. 2). However, stomatal width was significantly higher in TD under enriched CO2 compared to ambient CO2 (p<0.05). Overall, even though not significant, enriched CO2 increased the stomatal width compared to ambient CO2. The widest stomata were found in TD water stress treatment under enriched CO2 and the narrowest stomata was found in TD under ambient conditions.
Stomatal densities (Fig. 3) were not significantly different between the ambient-FD vs enriched-FD and ambient-SD vs enriched-SD treatments (p>0.05). However, for TD water stress treatment, stomatal density was significantly higher (p<0.05) under enriched CO2 environment compared to ambient CO2 environment. In addition, under ambient CO2, stomatal densities were similar under all water stress treatments. However, under enriched CO2, stomatal density in the water stress treatments increased in the order FD<SD<TD. The TD treatment showed significantly higher stomatal density (p<0.05) compared to both SD and FD, as mentioned earlier.
Epidermal cell lengths (p>0.05) and cell widths (p>0.05) were not significantly affected by the water stress and CO2 interaction (Fig. 4). However, epidermal cell width wassignificantly different among the CO2 levels (p<0.05).
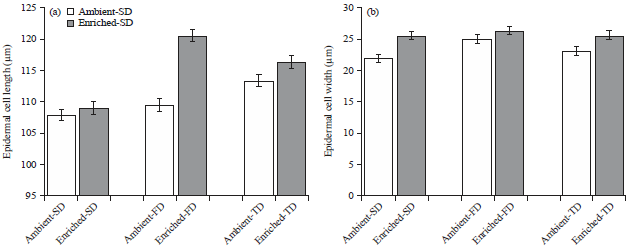 | |
| Fig. 4(a-b): | Length and width of epidermal cells (Mean±SEM) of maize leaves in three water stress treatments SD: 7 days, FD: 14 days and TD: 21 days) under ambient (435μmol mol–1) and enriched (1000μmol mol–1) CO2 |
A slight increase in epidermal cell lengths and widths were observed under FD and TD in both of the CO2 environments.
Plant growth measurements: Plant height was significantly higher under the combined effect of the water stress treatments and the CO2 enrichment treatments (p<0.05). The maize plants under the ambient CO2 environment were growing slow until 15 DIT in the SD, FD and TD treatments, compared to enriched CO2 environment. Gradually they grew taller and caught up with the plants in the same treatments under the enriched CO2, within 31 DIT (Fig. 5). From then until the end of the experiment, the water stressed plants under enriched CO2 treatments started to grow faster compared to their counter parts in the ambient CO2 treatments. In the comparison between the CO2 environments, largest height difference was observed in TD followed by SD and FD.
Plant girth in SD, FD and TD water stress treatments under the ambient CO2 were not significantly different from the enriched CO2 (p>0.05). The girth showed the opposite trend in growth compared to plant height. Until 31 DIT, girths of maize plants were lower in enriched treatments in comparison with ambient treatment; afterwards it surpassed the ambient treatment (Fig. 6). By 70th DIT, girths of water stressed plants under enriched CO2 treatments were higher than their ambient counterparts in ambient CO2. Plants under FD treatment showed the highest girth difference in between the CO2 environments. Plant girth values under enriched-TD and enriched-SD treatments were also higher compared to ambient-TD and ambient-SD, respectively.
DISCUSSION
Enriched CO2 environment significantly increased total chlorophyll content (TCC) in the drought treatments compared to the ambient CO2 environments. In enriched drought environments, 21 day-drought treatment (TD) showed highest total chlorophyll content, followed by 14 day-drought treatment (FD) and 7 day-drought treatment (SD). In the ambient drought environment, the SD treatment showed the highest TCC followed by TD and FD. However, the highest TCC recorded in ambient drought environment (11.67μg cm–2 in SD) was still less than that of second highest (13.50μg cm–2 in FD) in enriched drought environment. These results demonstrate that the CO2 enrichment effectively mitigated the negative effect of water stress on chlorophyll production.
Previously, it has been reported that total chlorophyll content was lower in the middle leaves of maize under CO2 enrichment, where the CO2 concentration was double of the ambient atmospheric CO220. Moreover, higher TCC was reported in that research compared to this study, both in ambient (around 27μg cm–2 vs 12.5μg cm–2) and enriched (15μg cm–2 vs 14μg cm–2) conditions. Driscoll et al.20, did not apply any water stress treatments and that might be the reason for the higher TCC in their study compared to this study. In addition, Driscoll et al.20, used higher levels of light intensity in their study (800μmol m–2 sec–1) compared to this study (163.15±10μmol m–2 sec–1), probably another reason for higher reported chlorophyll content. In this study, topmost fully fledged maize leaf (3rd leaf) received a PAR of 22.5±5μmol m–2 sec–1, when the plants were two weeks old and about 163.15±10μmol m2 sec–1, when they were 70 DIT (fully fledged 10th leaf).
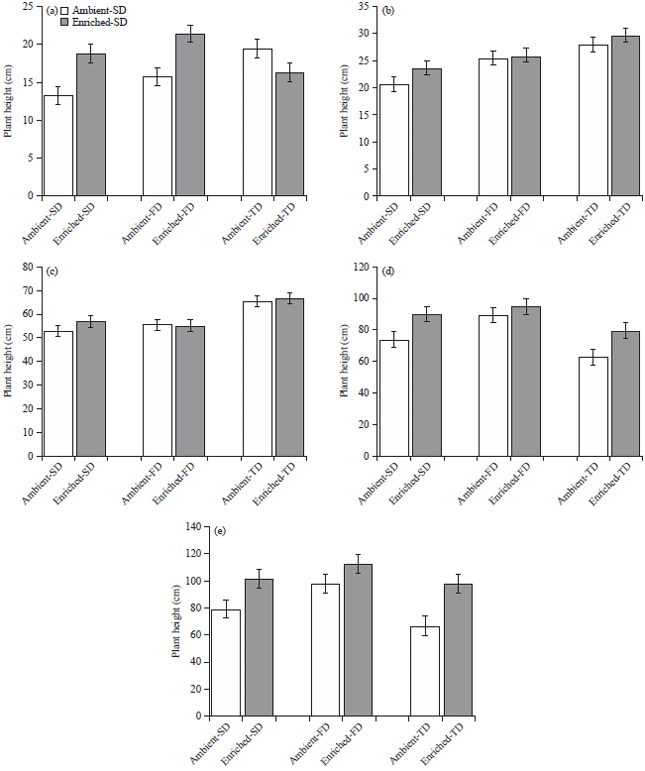 | |
| Fig. 5(a-e): | Height (Mean±SEM) of the maize plants in three water stress treatments SD: 7 days, FD: 14 days and TD: 21 days under ambient (435μmol mol–1) and enriched (1000μmol mol–1) CO2 |
Along with this, exposure to longer photoperiod (16 h of light Vs 15 h of light) for longer period of time (until 13th leaf stage Vs until 10th leaf stage) might also have contributed to the higher TCC in the above-mentioned study. Longer photoperiod has been reported to influence radiation use efficiency of maize leaves9. In addition, as per a previous study, TCC gradually increased up to VT stage of maize growth (13th leaf stage)26. In general chlorophyll ‘a’ was quantified higher than chlorophyll ‘b’, similar to those reported in non-water-stressed maize plants under ambient CO227. Zhu et al.28 reported that quantities of chlorophyll ‘a’ and chlorophyll ‘b’ in maize were not significantly different under drought stress.
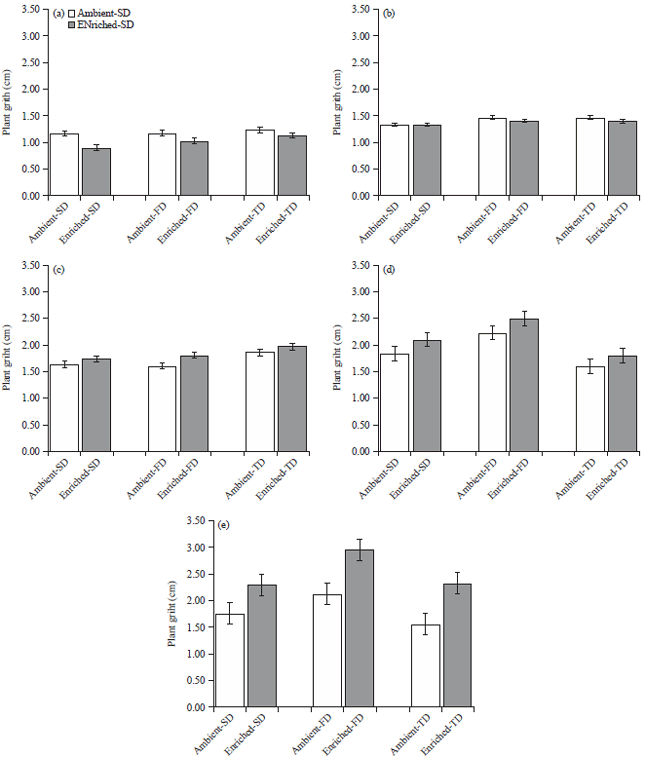 | |
| Fig. 6(a-e): | Stem girth (Mean±SEM) of the maize plants in three water stress treatments SD:7 days, FD: 14 days and TD: 21 days under ambient (435μmol mol–1) and enriched (1000μmol mol–1) CO2 |
Stomatal length was significantly higher (p<0.05) in enriched-SD and enriched-FD treatments compared to ambient-SD and ambient-SD treatments. In addition, stomatal length decreased (30.71μm) in enriched-TD when compared to enriched-FD (38.35μm). This indicates, that, as the water stress increases, further stomatal length reduction can be observed under enriched CO2 environment. However, Sarker and Hara29, reported that eggplant (C3 plant) grown under enriched CO2, stomatal length increased in 21-day water stress treatment. The difference in C3 and C4 stomatal arrangement and structure might have caused this difference. In this research, stomatal widths were not significantly different among the water stress treatments between ambient and enriched CO2 environments. The main effects, which were CO2 environments and water stress treatments, also did not show any significant impact on stomatal widths of maize leaves. In contrast to these results on stomatal widths, Sarker and Hara29, reported comparatively narrower stomata in 21-day water stress treatments under enriched CO2 environment compared to ambient CO2 environment. The contradictory results on stomatal length and width of this study, comparing to Sarker and Hara29, could be attributed again to the differences in monocot and dicot plant stomatal morphology and arrangement. In this study CO2 enrichment resulted in larger epidermal cells compared to ambient CO2. However, in other crops (e.g., rice) no significant change in the epidermal cell width under elevated CO2 levels has been reported30.
After 31 DIT of growth till the end of experiment (70 DIT), plants grown under enriched conditions grew taller than those under ambient conditions. The maximum plant growth is recorded in FD, while the maximum growth in between ambient and enriched conditions was recorded in TD. A previous study reported that plant height increased 14% only after 3 years of exposure to CO2 enrichment31. Further, two other studies reported that there is no difference in plant growth between ambient and enriched conditions32,33. Furthermore, Leakey et al.7, reported that in absence of water stress maize growth is not affected by elevated CO2. In these experiments, maize plants did grow bigger in presence of CO2 enrichment, contradicting the reports of Leaky et al.7 and Kim et al.32,33.
CONCLUSION
In the controlled environment growth chambers, enriched CO2 (1000μmol mol–1) increased vegetative growth of maize plants compared to ambient CO2, under low light condition. Although low PPFD affected the chlorophyll content, it did not hinder the overall drought mitigating effects of elevated CO2 on maize plants. The growth enhancement was much pronounced in 14-days water stress treatments. Under 1000μmol mol–1 CO2 and 14-days water stress, chlorophyll ‘a’ and chlorophyll ‘b’ and stomatal characteristics improved in such a way that they indirectly contributed to the overall plant growth. More levels of water stress treatments and CO2 enrichments should be studied in both growth chambers and outside growth chamber conditions (e.g., greenhouse or field), to find out how the maize plants react to the combined treatments of enriched CO2 and water stress.
SIGNIFICANCE STATEMENTS
The current research used elevated CO2 to mitigate the effect of drought stress in maize growing under low PPFD. The results showed that negative effect of drought stress, especially withholding irrigation for 2 weeks, was not pronounced in maize vegetative growth due to the positive effects of elevated CO2 conditions.
ACKNOWLEDGMENTS
This work was funded by the UAE University National Water Center (Grant No: 31R044). The authors are grateful to all colleagues at Biology Department, College of Science (UAE University) for their support and encouragement.
REFERENCES
- Manderscheid, R., M. Erbs and H.J. Weigel, 2014. Interactive effects of free-air CO2 enrichment and drought stress on maize growth. Eur. J. Agron., 52: 11-21.
CrossRefDirect Link - Bruce, W.B., G.O. Edmeades and T.C. Barker, 2002. Molecular and physiological approaches to maize improvement for drought tolerance. J. Exp. Bot., 53: 13-25.
CrossRefPubMedDirect Link - Chen, G., H. Liu, J. Zhang, P. Liu and S. Dong, 2012. Factors affecting summer maize yield under climate change in Shandong province in the Huanghuaihai region of China. Int. J. Biometeorol., 56: 621-629.
Direct Link - Lobell, D.B., W. Schlenker and J. Costa-Roberts, 2011. Climate trends and global crop production since 1980. Science, 333: 616-620.
CrossRefDirect Link - Cairns, J.E., J. Hellin, K. Sonder, J.L. Araus, J.F. MacRobert, C. Thierfelder and B.M. Prasanna, 2013. Adapting maize production to climate change in sub-Saharan Africa. Food Secur., 5: 345-360.
CrossRefDirect Link - Leakey, A.D.B., M. Uribelarrea, E.A. Ainsworth, S.L. Naidu, A. Rogers, D.R. Ort and S.P. Long, 2006. Photosynthesis, productivity and yield of maize are not affected by open-air elevation of CO2 concentration in the absence of drought. Plant Physiol., 140: 779-790.
CrossRefDirect Link - Lobell, D.B., M.J. Roberts, W. Schlenker, N. Braun, B.B. Little, R.M. Rejesus and G.L. Hammer, 2014. Greater sensitivity to drought accompanies maize yield increase in the U.S. Midwest. Science, 344: 516-519.
CrossRefDirect Link - Earl, H.J. and R.F. Davis, 2003. Effect of drought stress on leaf and whole canopy radiation use efficiency and yield of maize. Agron. J., 95: 688-696.
Direct Link - Chaves, M.M., J. Flexas and C. Pinheiro, 2009. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot., 103: 551-560.
CrossRefDirect Link - Din, J., S.U. Khan, I. Ali and A.R. Gurmani, 2011. Physiological and agronomic response of canola varieties to drought stress. J. Anim. Plant Sci., 21: 78-82.
Direct Link - Abu, S.S., 2014. Biochemical and photosynthetic evaluation of responses in Zea mays L. under drought stress. Curr. World Environ., 9: 87-95.
Direct Link - Ashraf, M. and P.J.C. Harris, 2013. Photosynthesis under stressful environments: An overview. Photosynthetica, 51: 163-190.
CrossRefDirect Link - Von Caemmerer, S. and R.T. Furbank, 2003. The C4 pathway: An efficient CO2 pump. Photosynthesis Res., Vol. 77.
CrossRefDirect Link - Ziska, L.H. and J.A. Bunce, 1997. Influence of increasing carbon dioxide concentration on the photosynthetic and growth stimulation of selected C4 crops and weeds. Photosynth. Res., 54: 199-208.
Direct Link - Wand, S.J.S., G.Y.F. Midgley, M.H. Jones and P.S. Curtis, 1999. Responses of wild C4 and C3 grass (poaceae) species to elevated atmospheric CO2 concentration: A meta-analytic test of current theories and perceptions. Global Change Biol., 5: 723-741.
CrossRefDirect Link - Sgheri, C.L.M., M.F. Quartacci, M. Menconi, A. Raschi and F. Navari-Izzo, 1998. Interactions between drought and elevated CO2 on alfalfa plants. J. Plant Physiol., 152: 118-124.
CrossRefDirect Link - Gouk, S.S., J. He and C.S. Hew, 1999. Changes in photosynthetic capability and carbohydrate production in an epiphytic CAM orchid plantlet exposed to super-elevated CO2. Environ. Exp. Bot., 41: 219-230.
CrossRefDirect Link - Driscoll, S.P., A. Prins, E. Olmos, K.J. Kunert and C.H. Foyer, 2005. Specification of adaxial and abaxial stomata, epidermal structure and photosynthesis to CO2 enrichment in maize leaves. J. Exp. Bot., 57: 381-390.
CrossRefDirect Link - Monje, O. and B. Bugbee, 1998. Adaptation to high CO2 concentration in an optimal environment: Radiation capture, canopy quantum yield and carbon use efficiency. Plant Cell Environ., 21: 315-324.
CrossRefDirect Link - Leakey, A.D.B., E.A. Ainsworth, C.J. Bernacchi, A. Rogers, S.P. Long and D.R. Ort, 2009. Elevated CO2 effects on plant carbon, nitrogen and water relations: Six important lessons from FACE. J. Exp. Bot., 60: 2859-2876.
CrossRefDirect Link - Richardson, A.D., S.P. Duigan and G.P. Berlyn, 2002. An evaluation of noninvasive methods to estimate foliar chlorophyll content. New Phytol., 153: 185-194.
CrossRefDirect Link - Schlemmer, M., A.A. Gitelson, J. Schepers, R. Ferguson, Y. Peng, J. Shanahan and D.C. Rundquist, 2013. Remote estimation of nitrogen and chlorophyll contents in maize at leaf and canopy levels. Int. J. Appl. Earth Observ. Geoinform., 25: 47-54.
CrossRefDirect Link - Ciganda, V., A.A. Gitelson and J. Schepers, 2009. Non-destructive determination of maize leaf and canopy chlorophyll content. J. Plant Physiol., 166: 157-167.
CrossRefPubMedDirect Link - Daughtry, C.S.T., C.L. Walthall, M.S. Kim, E.B. de Colstoun and J.E. McMurtrey III, 2000. Estimating corn leaf chlorophyll concentration from leaf and canopy reflectance. Remote Sens. Environ., 74: 229-239.
CrossRefDirect Link - Zhu, X.C., F.B. Song, S.Q. Liu, T.D. Liu and X. Zhou, 2012. Arbuscular mycorrhizae improves photosynthesis and water status of Zea mays L. under drought stress. Plant Soil Environ., 58: 186-191.
Direct Link - Sarker, B.C. and M. Hara, 2011. Effects of elevated CO2 and water stress on the adaptation of stomata and gas exchange in leaves of eggplants (Solanum melongena L.). Bangladesh J. Bot., 40: 1-8.
Direct Link - Tsutsumi, K., M. Konno, S.I. Miyazawa and M. Miyao, 2014. Sites of action of elevated CO2 on leaf development in rice: Discrimination between the effects of elevated CO2 and nitrogen deficiency. Plant Cell Physiol., 55: 258-268.
CrossRefDirect Link - Ainsworth, E.A. and S.P. Long, 2005. What have we learned from 15 years of Free‐Air CO2 Enrichment (FACE)? A meta‐analytic review of the responses of photosynthesis, canopy properties and plant production to rising CO2. New Phytol., 165: 351-372.
CrossRefDirect Link - Kim, S.H., R.C. Sicher, H. Bae, D.C. Gitz, J.T. Baker, D.J. Timlin and V.R. Reddy, 2006. Canopy photosynthesis, evapotranspiration, leaf nitrogen and transcription profiles of maize in response to CO2 enrichment. Global Change Biol., 12: 588-600.
CrossRefDirect Link - Kim, S.H., D.C. Gitz, R.C. Sicher, J.T. Baker, D.J. Timlin and V.R. Reddy, 2007. Temperature dependence of growth, development and photosynthesis in maize under elevated CO2. Environ. Exp. Bot., 61: 224-236.
CrossRefDirect Link