
Chenggang Duan
Research Center for Preclinical Medicine, Luzhou Medical College, Luzhou 646000, Sichuan, China
Zhiqiang Mei
Research Center for Preclinical Medicine, Luzhou Medical College, Luzhou 646000, Sichuan, China
Shu Gong
Research Center for Preclinical Medicine, Luzhou Medical College, Luzhou 646000, Sichuan, China
Haiqing Yu
Research Center for Preclinical Medicine, Luzhou Medical College, Luzhou 646000, Sichuan, China
Research Journal of Botany
Year: 2011 | Volume: 6 | Issue: 2 | Page No.: 87-94







ABSTRACT
Penthorum sedoides L. and Penthorum chinense Push is closely related aneuploids in genus Penthorum L. based on the previous data. However, P. chinense are treated as variety or subspecies of P. sedoides successively by botanists and little was known about their relationships on molecular level. An improved randomly amplified polymorphic DNA (RAPD) and Sequence Characterized Amplified Region (SCAR) were performed to obtain their genetic characterizations and species-specific DNA fragments. Obviously, five 10-mer random primers SBS-A3, SBS-I3, SBS-I18, SBS-M6 and SBS-Q9 demonstrated different fingerprints. Six candidate specific bands (I3-PS, I3-PC, I18-PC, M6-PS, Q9-PS and Q9-PC) displayed in two taxa were successfully cloned and sequenced. Based on these sequences, six pairs of SCAR primers from P. sedoides (I3PSF/I3PSR, M6PSF/M6PSR and Q9PSF/Q9PSR) and P. chinense (I3PCF/I3PCR, I18PCF/I18PCR and Q9PCF/Q9PCR) were designed, respectively. However, four of six primer pairs yielded amplicons common in the two taxa. The DNA amplification using Q9PSF/Q9PSR primers generated a single 555 bp band only in P. sedoides and Q9PCF/Q9PCR primers produced a 760 bp fragment unique to P. chinense. The results suggested that P. sedoides and P. chinense are still closely related, although there are a lot of variations in RAPD genetic sites. The developed RAPD and SCAR techniques are effective and useful in revealing genetic characterizations of P. sedoides and P. chinense. Also, the SCAR analysis based on the improved RAPD method is powerful in authentication of species with close relationships.
PDF Abstract XML References Citation
Received: January 06, 2011;
Accepted: April 09, 2011;
Published: June 23, 2011
How to cite this article
Chenggang Duan, Zhiqiang Mei, Shu Gong and Haiqing Yu, 2011. Genetic Characterization and Authentication of Penthorum Species Using RAPD and SCAR Markers. Research Journal of Botany, 6: 87-94.
URL: https://scialert.net/abstract/?doi=rjb.2011.87.94
URL: https://scialert.net/abstract/?doi=rjb.2011.87.94
INTRODUCTION
Penthorum sedoides L. and Penthorum chinense Push are the only two species in genus Penthorum L. with a disjunct distribution, the two taxa are perennial rhizomatous herbs living in wet habitats (Haskins and Hayden, 1987; Wen, 1999; Pan et al., 2001). P. sedoides (ditch stonecrop) distributed in eastern North America is useful in making cough syrups (Foster and Duke, 1999) and P. chinense (ganhuangcao) located in eastern Asia is effective in protecting the liver cells (Cao et al., 2007; Zhou et al., 2008; Yu et al., 2011). They have similar morphological and anatomical features. Also, variations between them are detected. In which, P. chinense is slightly different to P. sedoides in its longer and narrower leaf blades and shorter and thicker styles (Hara, 1962). Cytologically, these plants are aneuploids each other. P. sedoides is diploid with 2n = 18 chromosomes while the chromosomes in diploid P. chinense are 2n = 16 (Cronquist, 1981).
The familial position of Penthorum has fascinated botanists and biogeographers since the Linnaean era. Penthorum species are considered to have close relationships to species in Crassulaceae, Saxifragaceae or Haloragaceae (Morgan and Soltis, 1993; Pan, 1995; Jian et al., 2008). Due to their disjunct distributions perhaps, the two species are seldom analyzed simultaneously, especially on molecular level (Fishbein et al., 2001; Davis and Chase, 2004; Moody and Les, 2007; Jian et al., 2008). Besides, P. chinense has ever been treated as variety or subspecies of P. sedoides. Therefore, it is necessary to reveal their genetic characterizations and authenticate the two taxa.
The randomly amplified polymorphic DNA (RAPD) technique has been shown to be useful in analysis of different species on molecular level (Lakshmi et al., 2008; Al-Atiyat, 2009; Shaptadvipa and Sarma, 2009; Su et al., 2009; Ruzicka et al., 2009; Thangaraj et al., 2011). Thereafter, the Sequence Characterized Applied Region (SCAR) which is usually converted from RAPD by specific primers, is a locus-specific technology with more reliability and more reproducibility for molecular discrimination (Paran and Michelmore, 1993). Recently, reliable SCAR markers have been already derived from RAPD fragments in different herbs (Devaiah and Venkatasubramanian, 2008; Liao et al., 2009). The aim of this study is to demonstrate the genetic characterizations and authenticate Penthorum species based on the RAPD and SCAR analysis.
MATERIALS AND METHODS
Plant materials: The taxa, accession numbers and geographic origins are listed in Table 1. P. sedoides in Richmond (Virginia, USA) is kindly provided by Dr. W. John Hayden and P. chinense is collected from Luzhou (Sichuan, China). The seeds were germinated and grown in the perennial nursery of Medicinal Botanical Garden, Luzhou Medical College. The mature plants were carefully identified by Dr. Haiqing Yu. All voucher specimens have been deposited at the Medicinal Botanical Association of Zhongshan Mountain (MBAZM), Luzhou Medical College.
Genomic DNA extraction: The leaf samples for each accession were collected from mature plants in the perennial nursery of Medicinal Botanical Garden and ground in liquid nitrogen in a 1.5 mL microfuge tube. DNA was extracted and purified with the cetyltrimethylammonium bromide (CTAB) procedure outlined by Doyle and Doyle (1990).
RAPD analysis: The PCR reaction was executed using SBS primers A3, I3, I18, M6 and Q9 (Beijing SBS Genetech Co., Ltd, China) (Table 2). RAPD was performed in a total volume of 20 μL containing 30 ng DNA, 1xreaction buffer, 2 mM MgCl2, 0.25 μM of each primer, 200 μM of each dNTP (TakaRa Biotechnology (Dalian) Co., Ltd), 1 unit of rTaq DNA polymerase (TakaRa) and sterile water to the final volume. 1 drop of mineral oil was added in each reaction tube.
| Table 1: | Materials used in this study |
 | |
The thermal profile consisted of an initial denaturation at 94°C for 4 min, 40 cycles of 1 min at 94°C, 1 min at 36°C, 2 min at 72°C, followed by final extension of 10 min at 72°C. PCR reactions of each accession were carried out in a Mastercycler 5331 (Eppendorf, Germany). The amplified PCR products were resolved by electrophoresis on 2% agarose gel in 1xTAE buffer. Gels were visualized by 0.5 μg mL-1 ethidium bromide staining and the images were documented using the ChemiDoc XRS Bio-Rad, USA). An improved method for increasing the efficiency of RAPD by prolonging the ramp time from annealing to extension and increasing the resolution and production was introduced by Fu et al. (2000).
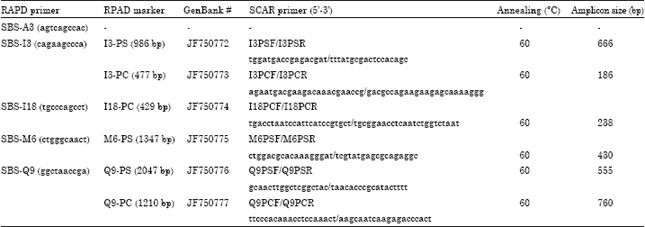
RAPD bands cloning and sequencing: The expected species-specific RAPD bands were excised from the agarose gels and purified using the Gel Extraction Kit (50) (Omega, GA, USA). These amplification products were then linked into a pMD18-T Easy Vector Systems according to the manufacturer’s instruction (TakaRa). The transformed competent E. coil DH5α (TakaRa) were plated on LB solid medium containing ampicillin (Sigma, USA) and cultured overnight at 37°C. The candidate clones were confirmed directly using specific M13F/M13R primers via PCR amplification. Five positive clones for each species were randomly selected and sequenced in both directions by Sunbiotech Co., Ltd. (Beijing, China). The cloned sequences alignment was executed with Clustal X program (Thompson et al., 1997) to reveal their homology and confirm the presence of a unique amplified product in the RAPD marker band, respectively. The accession numbers of six sequences used in this study are listed in Table 2.
SCAR primer designing and detecting: Six pairs of candidate specific primers from six sequences were designed using Primer Premier 5.0 (Premier Biosoft, USA) and synthesized in SBS Genetech (China). Primers data were summarized in Table 2. Based on PCR amplification, these primers were used to detect specific DNA fragments between the two species. The SCAR reaction program was as follows: 94°C for 3 min, 30 cycles at 94°C for 40 s, 60°C for 50 sec, 72°C for 1 min and final extension at 72°C for 8 min. This thermal profile was optimized and standardized according to the specific Tm of the primer pair. The gel electrophoresis and image documentation conditions were as described above.
RESULTS
Primers SBS-A3, SBS-I3, SBS-I18, SBS-M6 and SBS-Q9 were initially investigated for genetic characterizations between the two taxa. The five primers produced clear and reproducible RAPD fingerprints ranging in size from about 250 bp to 2000 bp (Fig. 1). Obviously, all primers demonstrated different amplification band patterns. Six expected specific bands displayed by the latter four primers were successfully cloned and sequenced, respectively. These RAPD markers were named based on information of primers and species (Table 2). Among which, I3-PS, M6-PS and Q9-PS fragments were unique to P. sedoides while I3-PC, I18-PC and Q9-PC fragments were found only in P. chinense (Fig. 1).
Sequence alignment showed that the similarity of clones from each unique fragment of P. sedoides was 100% and all clones from each single band of P. chinense were consistent too. After blast, no significant similar sequence of all RAPD fragments except M6-PS was detected in the NCBI nucleotide collection (nr/nt) database. M6-PS is closely related to predicted hypothetical protein of Vitis vinifera (accession number XM_002285443) and full-length cDNA clone from Zea mays (accession number BT063499) with 80% and 77% identities in query sites, respectively (Fig. 2). Besides, no open reading frame was revealed in the other five RAPD markers.
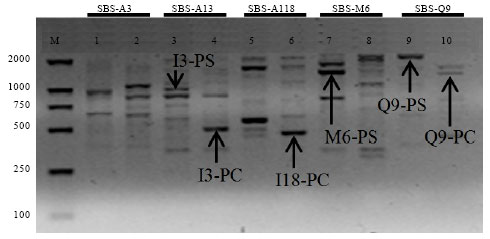 | |
| Fig. 1: | RAPD fingerprints of P. sedoides and P. chinense with primers SBS-A3, SBS-I3, SBS-I18, SBS-M6 and SBS-Q9. Lanes 1, 3, 5, 7 and 9 = P. sedoides; Lanes 2, 4, 6, 8 and 10 = P. chinense. M, DL2000 DNA ladder (bp). Arrows indicate the cloned RAPD genetic characterization bands (I3-PS = 986 bp, I3-PC = 477 bp, I18-PC = 429 bp, M6-PS = 1347 bp, Q9-PS = 2047 bp and Q9-PC = 1210 bp) unique to the two species, respectively |
 | |
| Fig. 2: | Identical hits of M6-PS sequence blast to NCBI nucleotide collection (nr/nt) database. The predicted hypothetical protein of Vitis vinifera (accession number XM-002285443) (A) and full-length cDNA clone from Zea mays (accession number BT063499) (B) are close related to M6-PS with 80 and 77% identities in query sites, respectively |
Six pairs of SCAR primers with longer lengths and higher annealing temperatures were designed according to six specific sequences (Table 2). These forward and reverse primers are located in interiors of six RAPD marker sequences, respectively. When using primers I3PSF/I3PSR, M6PSF/M6PSR and Q9PSF/Q9PSR to detect P. sedoides, the former two generated the expected 666 bp and 430 bp amplicons in the two taxa and primers Q9PSF/Q9PSR produced a 555 bp fragment unique to P. sedoides (Fig. 3). To diagnose P. chinense, I3PCF/I3PCR, I18PCF/I18PCR and Q9PCF/Q9PCR primers were used in analysis. In Q9PCF/Q9PCR amplification, the wanted 760 bp fragment was displayed only in P. chinense, however, the 186 bp and 238 bp bands were common in the two taxa when using primers I3PCF/I3PCR and I18PCF/I18PCR (Fig. 3).
| Table 2: | Primers used in RAPD and SCAR analysis |
 | |
| -: No specific band is cloned | |
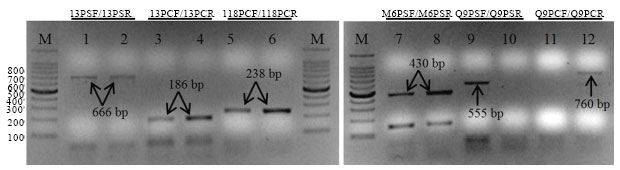 | |
| Fig. 3: | SCAR analysis of P. sedoides and P. chinense using primers pairs I3PSF/I3PSR, I3PCF/I3PCR, I18PCF/I18PCR, M6PSF/M6PSR, Q9PSF/Q9PSR and Q9PCF/Q9PCR. Lanes 1, 3, 5, 7, 9 and 11 = P. sedoides; Lanes 2, 4, 6, 8, 10 and 12 = P. chinense. M, 100 bp DNA ladder (bp). Arrows indicate the exact sizes of amplified fragments |
DISCUSSION
Five RAPD primers generated clear, reproducible and distinguishable band patterns in the current study. This indicated that it is effective to detect the genetic characterizations of two taxa via the improved RAPD technique. Based on the RAPD analysis, the genetic characterizations among different herbs were also revealed, respectively (Devaiah and Venkatasubramanian, 2008; Ruzicka et al., 2009). Due to the low annealing temperature conditions in RAPD analysis, the specific RAPD bands are usually cloned and converted into SCAR markers to improve the efficiency and stability of diagnosis (Liao et al., 2009; Ruzicka et al., 2009). Thus, six candidate fragments obtained by four RAPD primers were sequenced in two species respectively to reveal their RAPD genetic characterizations.
The published data indicated that RAPD markers often displayed specific sequences in introns (Lubbers et al., 1994; Pessino et al., 1997). When blasting six RAPD sequences in NCBI database, no similar query item and open reading frame were found except partial M6-PS fragment. This also suggested that most cloned sequences are likely located in non-coding regions. Introns in RAPD fragments could play an important role in positive or negative regulation of eukaryotic gene expression (Salgueiro et al., 2000; Hisatsune et al., 2005). However whether these sequences participate gene expression needs to be detected further. Moreover, M6-PS in P. sedoides most likely belongs to one kind of unknown function gene.
In the two taxa, all RAPD primers generated obviously dissimilar fingerprints. However, four of six candidate specific primers subsequently demonstrated that the amplicons are common in P. sedoides and P. chinense, respectively. This suggested that although there are a lot of variations in RAPD genetic sites, P. sedoides and P. chinense are still closely related. Lee et al. (1996) also revealed their near relationships base on allozymes and ITS sequences analysis. According to SCAR analysis, four SCAR primers showed the identical bands among two species respectively while it was different in the amplification of Q9PSF/Q9PSR or Q9PCF/Q9PCR primers. Therefore, it is most likely that Q9-PS and Q9-PC sequences are unique to P. sedoides and P. chinense respectively while the other four cloned sequences are common in the two taxa.
To ensure the reproducibility and stability, each of amplification was repeated at least five times in SCAR detection and all the expected bands were identical. In the present study, the developed RAPD and SCAR techniques are effective and useful in revealing genetic characterizations of P. sedoides and P. chinense. Also, the SCAR analysis based on the improved RAPD method is powerful in authentication of species with close relationships.
ACKNOWLEDGMENTS
The authors contribute equally to the present study. We are thankful to the Youth Foundation, the National Natural Science Pre-Research Foundation and the Scientific Research Starting Foundation for Doctors of Luzhou Medical College (No. 676, 409 and 606). The authors particularly thank Dr. W. John Hayden (University of Richmond, Virginia, USA) and Qifeng Wang (Luzhou Medical College, Sichuan, China) for providing leaves and seeds, respectively.
REFERENCES
- Al-Atiyat, R.M., 2009. DNA polymorphism of indigenous chickens in Jordan. Aian J. Anim. Vet. Adv., 4: 237-244.
CrossRefDirect Link - Cao, H., J. Yang, Z.S. Peng, C.Y. Kang, D.C. Chen, Z.C. Gong and X. Tan, 2007. Micropropagation of Penthorum chinense through axillary bud. In vitro Cell. Dev. Plant, 43: 149-153.
CrossRef - Davis, C.C. and M.W. Chase, 2004. Elatinaceae are sister to malpighiaceae; Peridiscaceae belong to Saxifragales. Am. J. Bot., 91: 262-273.
Direct Link - Devaiah, K.M. and P. Venkatasubramanian, 2008. Genetic characterization and authentication of Embelia ribes using RAPD-PCR and SCAR marker. Planta Med., 74: 194-196.
PubMed - Fishbein, M., C. Hibsch-Jetter, D.E. Soltis and L. Hufford, 2001. Phylogeny of Saxifragales (Angiosperms, Eudicots): Analysis of a rapid, ancient radiation. Syst. Biol., 50: 817-847.
PubMed - Fu, J.J., L.Y. Li, X. Xu, Z. Wang, G. Tang, C.M. Yin and G.X. Lu, 2000. An improved method for increasing the efficiency of the technique of random amplified polymorphic DNA (RAPD). Hereditas, 22: 251-252.
Direct Link - Hara, H., 1962. Racial differences in widespread species, with special reference to those common to Japan and North America. Am. J. Bot., 49: 647-652.
Direct Link - Haskins, M.L. and W.J. Hayden, 1987. Anatomy and affinities of Penthorum. Am. J. Bot., 74: 164-177.
Direct Link - Hisatsune, H., K. Matsumura, M. Ogawa, A. Uemura and N. Kondo et al., 2005. A high level of endo-thelial cell-specific gene expression by a combination of 5'flanking region and 5'half of the first intron of VE-cadherin gene. Blood, 105: 4657-4663.
CrossRef - Jian, S.G., P.A. Soltis, M.A. Gitzendanner, M.J. Moore and R.Q. Li et al., 2008. Resolving an ancient, rapid radiation in Saxifragales. Syst. Biol., 57: 38-57.
PubMed - Lakshmi, P., P.A.A. Khan, P.N. Reddy, K. Lakshminarayana and S. Ganapaty, 2008. Genetic relationship among Tephrosia species as revealed by RAPD analysis. Asian J. Biol. Sci., 1: 1-10.
CrossRefDirect Link - Lee, N.S., T. Sang, D.J. Crawford, S.H. Yeau and S.C. Kim, 1996. Molecular divergence between disjunct taxa in Eastern Asia and Eastern North America. Am. J. Bot., 83: 1373-1378.
Direct Link - Liao, L.Q., J. Liu, Y.X. Dai, Q. Li and M. Xie et al., 2009. Development and application of SCAR markers for sex identification in the dioecious species Ginkgo biloba L. Euphytica, 169: 49-55.
CrossRef - Lubbers, E.L., L. Arthur, W.W. Hanna and A.P. Ozias, 1994. Molecular markers shared by diverse apomictic Pennisetum species. Theor. Appl. Genet., 89: 636-642.
CrossRef - Moody, M.L. and D.H. Les, 2007. Phylogenetic systematics and character evolution in the angiosperm family Haloragaceae. Am. J. Bot., 94: 2005-2025.
Direct Link - Morgan, D.R. and D.E. Soltis, 1993. Phylogenetic relationships among Saxifragaceae sensu lato based on rbcL sequence data. Ann. Missouri Bot. Garden, 80: 631-660.
Direct Link - Pan, J.T., 1995. A study on the tribe Astilbeae Miq. (Saxifragaceae). Acta Phytotaxon. Sin., 33: 390-402.
Direct Link - Pan, J.T., C. Gu, S. Huang, C.F. Wei and S.Y. Jin et al., 2001. Flora of China, Penthorum L. FOC, 8: 269-272.
Direct Link - Paran, I. and R.W. Michelmore, 1993. Development of reliable PCR-based markers linked to downy mildew resistance genes in lettuce. Theor. Applied Genet., 85: 985-993.
CrossRef - Pessino, S.C., J.P.A. Ortiz, O. Leblanc, C.B. Valle, C. Evans and M.D. Hayward, 1997. Identification of a maize linkage group related to apomixis in Brachiaria. Theor. Appl. Genet., 94: 439-444.
CrossRef - Ruzicka, J., B. Lukas, L. Merza, I. Gohler, G. Abel, M. Popp and J. Novak, 2009. Identification of Verbena officinalis based on ITS sequence analysis and RAPD-derived molecular markers. Planta Med., 75: 1271-1276.
PubMed - Salgueiro, S., C. Pignocchi and M.A.J. Parry, 2000. Intron-mediated gusA expression in tritordeum and wheat resulting from particle bombardment. Plant Mol. Biol., 42: 615-622.
CrossRef - Shaptadvipa, B. and R.N. Sarma, 2009. Assessment of nature and magnitude of genetic diversity based on DNA polymorphism with RAPD technique in traditional glutinous rice (Oryza sativa L.) of Assam. Asian J. Plant Sci., 8: 218-223.
CrossRefDirect Link - Su, X., K. Sun, W. Chen, H. Zhang and R.J. Ma, 2009. Molecular identification and genetic relationship between Viola yedoensis with its related species by RAPD. J. Tradit. Chin. Vet. Med., 28: 8-10.
Direct Link - Thangaraj, M., V. Prem, T. Ramesh and A.P. Lipton, 2011. RAPD fingerprinting and demonstration of genetic variation in three pathogens isolated from mangrove environment. Asian J. Biotechnol., 3: 269-274.
CrossRefDirect Link - Thompson, J.D., T.J. Gibson, F. Plewniak, F. Jeanmougin and D.G. Higgins, 1997. The clustal X windows interface: Exible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res., 25: 4876-4882.
PubMed - Wen, J., 1999. Evolution of Eastern Asian and eastern North American disjunct distributions in flowering plants. Annu. Rev. Ecol. Syst., 30: 421-455.
Direct Link - Yu, H.Q., C. Zhang, Z.Q. Mei, L. Wang and J. Li et al., 2011. Molecular authentication of medicinal Penthorum chinense push from different localities in China by RAPD analysis. Int. J. Bot., 7: 97-102.
CrossRef - Zhou, X.M., Q.Z. Li, Y.Z. Yin and Y.Y. Chen, 2008. Identification of medicinal Ganoderma species based on PCR with specific primers and PCR-RFLP. Planta Med., 74: 197-200.
PubMed