
B. Solomon Tefera
Department of Animal Science, Faculty of Agriculture and Natural Science, University of Namibia, P/Bag 13301, Windhoek, Namibia
V. Mlambo
Department of Food Production, The University of the West Indies, St Augustine, Trinidad and Tobago, West Indies
Research Journal of Botany
Year: 2010 | Volume: 5 | Issue: 1 | Page No.: 1-13







ABSTRACT
This study was conducted in the semi-arid Borana rangelands of Ethiopia and focused on the distribution pattern of Acacia brevispica and Acacia drepanolobium and their influence on sub-canopy grasses. Both species had significantly greater total densities on communal lands than on a government ranch and on the nearest site than on the furthest site from water points. A total of 23 grass species were identified in the sub-canopy and open habitats surrounding A. brevispica and A. drepanolobium. Cenchrus ciliaris and Chrysopogon aucheri were dominant species surrounding A. drepanolobium in both habitats. For grasslands surrounding A. brevispica, Themeda triandra was the dominant sub-canopy grass species, while C. aucheri, Panicum turgidum and Loudetia flavida dominated open habitats. Sub-canopy habitats in both species had significantly higher yields of total, highly and intermediately desirable grasses than open habitats. Although A. brevispica and A. drepanolobium have encroached due to prolonged heavy grazing, they did not negatively impact on sub-canopy grass productivity and, therefore, their control should be considered with caution. Future research is required to examine if changes in total tree density or cover may alter results of this study. Research is also needed on determinants of changes in sub-canopy grass productivity.
PDF Abstract XML References Citation
How to cite this article
B. Solomon Tefera and V. Mlambo, 2010. Encroachment of Acacia brevispica and Acacia drepanolobium
in Semi-Arid Rangelands of Ethiopia and their Influence on Sub-Canopy Grasses. Research Journal of Botany, 5: 1-13.
URL: https://scialert.net/abstract/?doi=rjb.2010.1.13
URL: https://scialert.net/abstract/?doi=rjb.2010.1.13
INTRODUCTION
The semi-arid southern Borana rangelands of Ethiopia are dominated by savanna vegetation characterised by a continuous and well-developed layer of grasses, forbs and woody plants. Some decades ago, Borana rangelands were considered among the best grazing lands in East Africa (Gemedo et al., 2006). However, in the intervening years, increasing densities of woody plants have been reported as a serious ecological problem in this region (Angassa, 2005). The negative attitude towards bush encroachment has provided the impetus for research to identify the causes of bush encroachment, control methods and their implementation. Few studies have examined the woody-herbaceous interactions in Borana savanna ecosystems, especially by relating tree stand density and/or canopy cover to herbage production and composition.
Trees and grasses are known to compete with each other in savannas (Chesson, 2000; Amarasekare, 2003; Sankaran et al., 2004). Several studies have demonstrated that in most African savannas, grass yield decreases as tree density increases (Sankaran et al., 2004; Angassa, 2005) which has often been associated with the reduction of herbaceous productivity under canopy-root zones of individual trees. This forms the basis of the rationale for tree thinning or complete clearing and tree removal resulted in increased dry matter yield and composition. In contrast, some reports addressed the role of woody plants in improving sub-canopy grass yield (Smit and Swart, 1994; Belsky et al., 1993). This effect, however, can be explained by several factors such as density of woody plants, amount of rainfall and soil type (Teague and Smit, 1992; Belsky, 1994). A study by Billé and Corra (unpublished) reported no difference in grass productivity under and outside tree canopy habitats.
Several species of Acacia, particularly A. brevispica Harms and A. drepanolobium Harms ex Y.Sjöstedt have been reported to encroach the southern rangelands of Ethiopia (Oba et al., 2000; Angassa and Baars, 2000; Angassa,, 2005; Solomon et al., 2007). Nevertheless, there is little information on the density and height distribution of these species in relation to different land use systems and distance gradient from a point of reference (e.g., water point). A previous study by Solomon et al. (2007) focused on a larger area of land whereas this study focuses on a smaller area where A. drepanolobium and A. brevispica are problematic encroachers. In addition, few studies have examined the effects of these encroachers on the productivity of the surrounding grass layer. Understanding the distribution of these woody plants and their interaction with herbaceous plants that are vital for livestock production is critical to development of effective bush control and management systems. The objectives of this study were therefore to investigate: (1) the distribution of A. brevispica and A. drepanolobium under two land use systems (communal grazing land and government ranch) and along a distance gradient from watering point and (2) the effect of individual trees of these species on the composition and productivity beneath and between their canopies
MATERIALS AND METHODS
Study Area
The study was conducted in the Borana rangelands of Southern Ethiopia from December 2000-May 2001. The Borana plateau covers approximately 95 000 km2 and ranges in altitude generally from 1000-1500 m, with peaks up to 2000 m. Rainfall is bimodal with annual rainfall ranging from 400 to 600 mm. Bushlands and thickets, Gemedo by Acacia and Commiphora species, cover the major parts of the rangelands (Gemedo et al., 2006). Granitic and volcanic soils and their mixtures cover the area (Coppock, 1994). Bottomland and upland soils, which characterize the study areas, are dominated by vertisols and red soil, respectively. The pH of the soil in study areas ranged from 6.34-6.98; the sand, silt and clay contents were 70.69-75.2%, 17.59-25.06% and 7.3-11.42%, respectively (Solomon et al., 2007). Percent organic Carbon and Nitrogen were in the range of 0.96-1.56 and 0.22-0.24, respectively (Solomon et al., 2007). Area is shown in Fig. 1.
Survey of Acacia brevispica and Acacia drepanolobium
This survey was designed to include two adjacent land use systems and a distance gradient from water sources. For the first part of the survey, the two land uses were communal grazing land (04°11'37.6''N, 05°48'72.4''E) and a government ranch (Did-Tuyura Ranch) (04°12'63.5''N, 05°47'89.1''E).
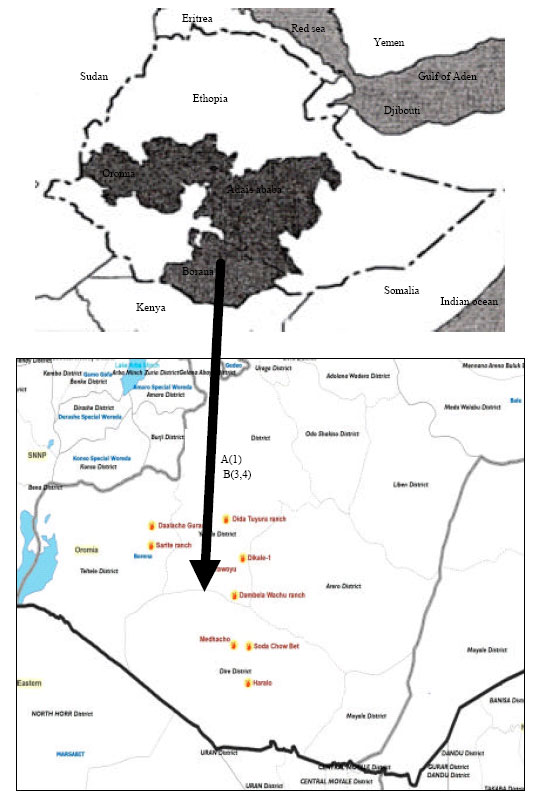 | |
| Fig. 1: | Map showing the study area A (1)-the communal land and B (3,4)-sites in the government ranch |
The soil, landscape and altitude in these areas are similar. Unfortunately, there is a lack of accurate information on the grazing history of these two land uses. However, the communal area experienced a higher grazing pressure over an extended period with estimated stocking rates of between 0.17 and 0.32 head ha-1 from 1982 to 1996 (Desta and Coppock, 2002). The stocking rate at the ranch during this period was 0.12 heads ha-1 which was lower than that of the communal land. The ranch is mainly grazed by cattle, while communal lands have different herbivores: grazers (mainly cattle and sheep) and browsers (goats and camels). For this study, 7 plots of 20x50 m were randomly surveyed in the ranch and the communal land, respectively, to record the distribution of A. brevispica and A. drepanolobium.
For the second part of the survey, four watering points were selected on the communal land. The selection criteria ensured general similarity in water holding capacity and age. We excluded watering points with either small capacity (<6000 m3) or with very recent age (<10 years old). The watering points are mechanically excavated water sources constructed in the 1980s to reduce grazing pressure around permanent water sources and to increase utilization of wet season grazing areas (Oba, 1998). One transect (about 6 km) was established at each of the selected ponds in an anticipated direction of main grazing activity evident through the density of cattle trails. Each transect was divided into three sub-transects (2 km each) and was recorded as near, middle and far from each water point. In the middle of each sub-transect a plot of 20x50 m was marked.
In both surveys three 10x10 m sub-plots were marked along the centre of each main plot (20x50 m). All rooted live A. brevispica and A. drepanolobium were recorded in each sub-plot and allocated to one of eight height classes: >0-0.5 m; >0.5-1 m; >1-1.5 m; >1.5-2 m; >2-3 m; >3-4 m; >4-5 and >5 m (Friedel, 1987). Tree data were standardized to tree equivalents/ha (1 TE = 1 tree, 1.5 m high) (Teague et al., 1981).
Woody-Grass Layer Interaction
This part of the study was carried out at Did-Tyura Ranch which is 17 km Northeast of Yabello town. Sites were selected from bottomland and upland soils. The bottomland and upland represent grasslands with scattered trees dominated by A. drepanolobium and A. brevispica, respectively. Approximately 2 ha of land were demarcated in each site. The sites were excluded from grazing for two years before the experiment started. Ten mature trees of A. drepanolobium (bole height >4 m; mean canopy area 31.7±5.4 m2) and A. brevispica (bole height >3 m; mean canopy area 22.6±3.6 m2) were selected. Each tree was fenced with wood in a radius of 13 m from the tree trunk in the dry season in December 2000. The grass layer surrounding each tree was divided into two main habitats: sub-canopy and open habitats. Four sites: >0-1 m, >1-3 m, >3-5 m and >5-9 m from the tree trunk were established in areas dominated by A. drepanolobium. Sites ≤3 m represented sub-canopy habitat whereas those >3 m were open habitats. For A. brevispica area, 3 sites: >0-1 m, >1-3 m and >6-9 m were surveyed where a distance >6 m represented the open habitat. The distance >3-6 was not included because this could not be clearly identified as canopy zones or open habitats. Grass shoots were harvested to stubble height from four 0.25 m2 quadrats in both the sub-canopy and open sites of both tree species giving a total of 280 quadrats. The quadrats were oriented in the four directions. The harvested materials were separated into species, oven dried to a constant weight at 60°C and weighed using digital weighing scale to determine dry matter (DM) yield and species composition. Sampling was done late in the growing season (April, 2001).
Species Identification and Classification
Most grasses were identified in the field. For the remaining species, plants with full inflorescences and other vegetative parts were collected and taken to the national herbarium of Ethiopia at Addis Ababa University for identification. Grasses were classified into desirability groups following the procedures of Tainton et al. (1980) and Vorster (1982). Accordingly, the species were grouped into (1) highly desirable species: those which occur in rangelands in good condition and which decrease with overgrazing (decreasers), (2) desirable species: those which occur in rangeland in good condition and increase with moderate over grazing (increasers IIa) and (3) less desirable species: those which occur in rangeland in good condition and increase with severe/extreme overgrazing (increasers IIb and IIc) (Table 2). In addition, species were grouped according to their life forms and their abundance (Solomon, 2003).
Data Analysis
A one-way analysis of variance (ANOVA) was used to examine variation in density of A. brevispica and A. drepanolobium along a distance gradient from water points, while densities between land uses were compared using a simple t-test. Mean differences were considered significant at p<0.05. One-way ANOVA was also used to analyse data on grass DM yield. All data were analysed using General Linear Model (GLM) of SAS (1999). Treatment means were separated using the PDIFF option of the least squares means statement of the GLM procedure of SAS (1999).
RESULTS
Density of Woody Plants
The total density of A. brevispica (TE ha-1) was greater (p<0.05) on communal land than on the ranch. There were more seedlings (>0-0.5 m) and saplings (>0.5-3 m) on communal land than in the ranch. For mature trees, the density of height class >3-4 m was significantly (p<0.05) greater on communal land than in the ranch, while the height class >4 m was 55% less on the communal land (Table 1). The greatest total density of A. brevispica (343 TE ha-1) occurred in the areas nearest to water where saplings were particularly more abundant. All the study sites had more A. brevispica saplings than trees or seedlings. Mature trees were the least abundant in the communal area but seedlings were most limited at the ranch.
For A. drepanolobium, total density was greater (p<0.05) on communal land than in the ranch. The population of seedlings (>0-1 m) and trees (>3-4 m) was higher (p<0.05) in the ranch than the communal land. In contrast, there were more saplings (>1-3 m) on communal land (Table 1). Saplings had the largest proportion of the total density followed by seedlings on communal land whereas the proportion of seedling was largest in the ranch. The total density of A. drepanolobium varied significantly (p<0.01) with the nearest site to the water having greatest value (571 TE ha-1). Similarly, the density of each height class decreased away from the water.
Grass Layer Composition
A total of 23 grass species were identified in the study areas (Table 2). Cenchrus ciliaris and C. aucheri were dominant around A. drepanolobium in the vertisol sub-canopy and open habitats. Pennisetum mezianum was present only in the open habitat where T. triandra occurred rarely.
| Table 1: | Density distribution of Acacia brevispica and Acacia drepanolobium under two land use systems and along a distance gradient from a water source |
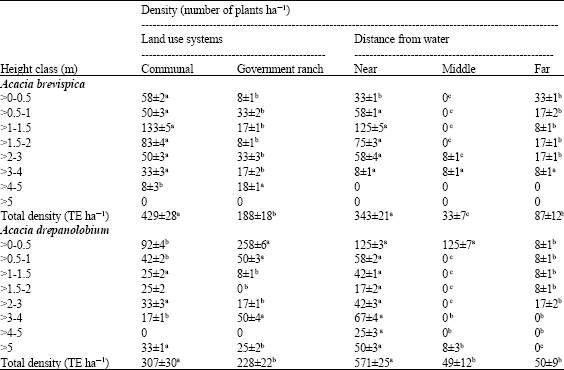 | |
| Means in the same row with different superscripts are significantly different at p≤0.05 | |
For red soil grasslands surrounding A. brevispica, T. triandra was the dominant sub-canopy species, while Andropogon sp., C. aucheri, Eustachys paspaloides, Heteropogon contortus and P. turgidium were common. Dominant grasses in the open habitat included C. aucheri, P. turgidum and L. flavida. Cenchrus ciliaris, C. aucheri, T. triandra andropogon sp., E. paspaloides and P. turgidium are highly palatable species (Table 2).
Grass Yield around A. brevispica
Total DM production was significantly greater (p<0.001) in the sub-canopy nearest (>0-1 m) to the tree trunks than the furthest sites from the trunk (>1-3 and >6-9 m, respectively). For highly desirable species, lowest DM yield for C. ciliaris and C. aucheri were recorded in the sub-canopy zone nearest to the trunk (>0-1 and 1-3 m, respectively). The production of E. paspaloides and P. turgidum was statistically similar in the sub-canopy and open habitat (Table 3). Greatest production (p<0.001) for T. triandra occurred below the canopy nearest to the trunk. DM yield for all highly desirable species together was 32 and 27% higher in the nearest sub-canopy zone (>0-1 m) than the sites at >1-3 and >6-9 m, respectively. Of the species with intermediate desirability, H. contortus attained the highest value in the nearest sub-canopy zone, whereas L. nutans had maximum production in the open habitats. When all intermediately desirable species were considered together, locations adjacent to the trunk had 97% greater (p<0.001) total DM yield than open habitat. Of less desirable species, L. flavida had greater (p≤0.05) production in open areas. For all the less desirable species, sites on the outside edge of crowns had higher production (p<0.01) than those adjacent to trunks (Table 3).
| Table 2: | Life forms, ecological groupings and botanical composition (%DM) of grasses in the study sites |
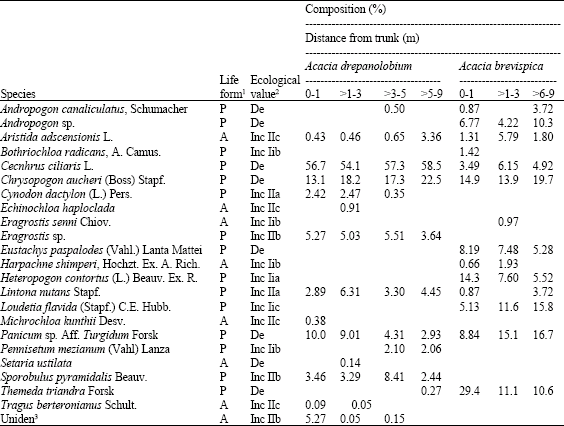 | |
| 1A: Annual, P: Perennial; 2De: Decreaser, Inc IIa: Increaser IIa, Inc IIb: Increaser IIb, Inc IIc: Increaser Iic; Dominant (>15%), Common (>5-15%), Present (1-5%), rare (≤ 1%). 3Uniden: Unidentified species | |
| Table 3: | Dry matter production (kg ha-1±SE) of grasses along the radius from the trunk of Acacia brevispica |
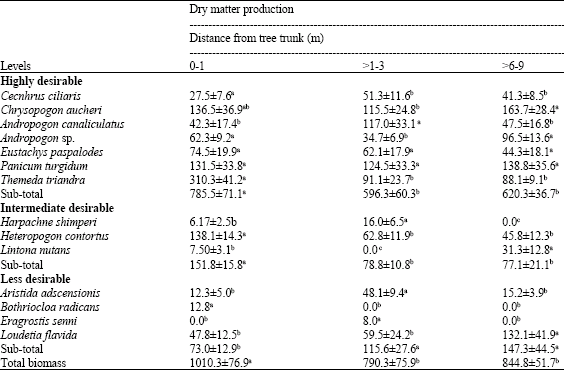 | |
| Means in the same row with different superscripts are significantly different at p≤0.05 | |
| Table 4: | Dry matter production (kg ha-1±se) of grasses along the radius from the trunk of Acacia drepanolobium |
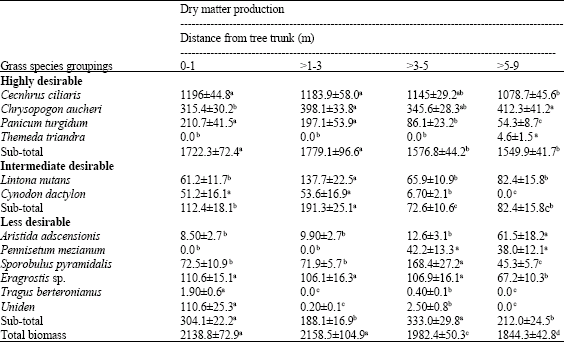 | |
| Means in the same row with different superscripts are significantly different at p≤0.05 | |
Grass Yield Around A. drepanolobium
The greatest (p<0.001) total DM yield occurred in the sub-canopy zone (>0-3 m), while the lowest value was obtained in the open habitat. Of the highly desirable species, C. ciliaris and P. turgidum had the greatest production in the sub-canopy zone (>0-3 m). The yield of C. aucheri was not significantly affected by distance from the trees. Dry matter yield of all the highly desirable species was greater (p<0.01) in the sub-canopy than open habitats. For intermediately desirable species, production of L. nutans did not show a particular trend although the highest value was obtained in one of the sub-canopy sites. Cynodon dactylon had significantly greatest production in the site closest to the trunk. When all intermediately desirable species were combined, DM yield was highest (p<0.001) in the sub-canopy zone. For less desirable species, production of S. pyramidalis and Eragrostis sp. was significantly lowest (p≤0.05) in the open habitat where DM yield of A. adscensionsis was highest. When less desirable species were added together, production in the sub-canopy and open habitats differed significantly (p<0.01) but this was not consistent (Table 4).
DISCUSSION
The current study indicated that saplings and total densities of A. brevispica and A. drepanolobium were higher on communal land than on the ranch. This observation could be due to their difference in terms of grazing and browsing pressure. Stocking rate at the ranch was estimated to be between 0.12 heads ha-1 for the years 1988-1998 (Solomon et al., 2007), which was lower than on the communal land (Desta and Coppock, 2002). Heavy grazing pressure reduces grass cover enabling the woody plants to get a competitive edge for available moisture and soil nutrient on soil surface (Solomon et al., 2007).
Populations of A. brevispica and A. drepanolobium were highest in the sites nearest to the water. This is consistent with the reports of Coppock (1994) and Solomon et al. (2007) which revealed that the two species may be key indicators of rangelands that have been subjected to high grazing and browsing pressure over long period of time (Coppock, 1994; Solomon et al., 2007). In Borana rangelands, areas close to water are subject to high livestock densities compared to distant grazing points and hence concentration of nutrients close to water points. When the herbaceous layer is depleted around the water point, import of nutrients by grazers and browsers (via dung and urine) most likely increases a macro-and micro element concentration (Moleele and Perkins, 1998). This process, when repeated over a long period, should eventually contribute to establishment of dense woody plants around watering points. More specifically, high browse pressure around water points could favour encroachers at the expense of non-encroachers. Animals browse selectively (Victor, 1981) in such a way that while there may be many woody species on offer it is usual for a limited number of them to form a significant proportion of their diet (Moleele and Perkins, 1998). Hence, palatable woody species without any protection against browsing will not be able to survive the high browsing pressure close to the water points (Moleele, 1994), while species such as A. brevispica and A. drepanolobium are not extensively browsed since they are protected by thorns. As a result, emergence and the establishment of these plants are favoured in conducive environment characterised by low competition from non-encroachers and the herbaceous layer (Moleele, 1994).
Most study sites showed a dominance by saplings, which has been reported in many encroached rangelands (Childes and Walker, 1987; Solomon et al., 2007). In the context of this study, this can be explained in terms of age of plants and/or retarded growth of individual woody plants due to spatial and temporal competition for water and nutrients between closely spaced plants (Riginos and Young, 2007). A dominance by saplings could be an indication of the potential for long-term increase. However, smaller size classes can be effectively controlled by burning provided there is enough grass fuel load to burn the bushes (Solomon, 2003).
Grass Species Composition and Distribution
Grass distribution in the study area corresponded partially with earlier studies (Angassa and Baars, 2000; Angassa, 2005). In this study, Cenchrus ciliaris and C. aucheri made up the bulk of the grass layer in the sub-canopy and open habitat of A. drepanolobium. Chrysopogon aucheri, P. turgidium and T. triandra formed a significant proportion in both habitats surrounding A. brevispica. Angassa (2005) reported that C. ciliaris was one of the most abundant species in the Borana rangeland, in contrast to Solomon et al. (2007), who reported a low occurrence in similar soil types. The high proportion of C. aucheri and T. triandra (Table 2) agrees with reports of Angassa (2005) and Solomon et al. (2007). Cenchrus ciliaris and C. aucheri are known as key valuable forage species in Borana (Coppock, 1994).
Grass Yield
Both A. brevispica and A. drepanolobium sub-canopy habitats had higher DM yield of highly desirable and intermediately desirable grasses and there was higher total DM production in the sub-canopy than open habitats. Conversely, the yield of less desirable grasses showed the highest values in open habitats. The higher yield of DM in the sub-canopy habitat can be attributed to a few high yielding species growing in association with tree crowns; i.e., T. triandra and H. contortus beneath A. brevispica; C. ciliaris and P. turgidium beneath A. drepanolobium. Studies that reported a higher sub-canopy grass yield have also shown one or two dominant, high yielding grass species growing in association with tree crowns e.g., Panicum maximum and Digitaria eriantha (Smit and Swart, 1994) or T. triandra (Moyo and Campbell, 1998). The pattern in DM yield of C. aucheri and E. paspaloides did not show a preference for sub-canopy or open habitats. Dry matter yield of C. ciliaris, T. triandra and P. turgidium differed between the two habitats, but the nature of the relationship was specific to tree species. Similarly, study by Smit and Swart (1994) also found a different reaction of individual grass species to the sub-canopy beneath Acacia erubescens Welw.ex Oliv. and Combretum apiculatum Sond compared with open habitat.
The current study does not support the results of Angassa and Beyene (2003) who reported that Borana rangelands have deteriorated due to invasion by A. brevispica and A. drepanolobium which was associated with increased production of unpalatable Pennisetum mezianum and P. stramineum. Pastoralists in this region associated such encroachment with reduced milk production, lowered calving rates and reduced livestock survival during drought (Oba et al., 2000). Gemedo et al. (2006) associated increased cover of woody plants in Borana rangelands with reduced grazing capacity. The authors have advocated the use of fire to control bush encroachment in the semi-arid Borana rangelands.
The current study, however, demonstrated that isolated trees of A. brevispica and A. drepanolobium enhanced grass productivity in the sub-canopy compared to the open habitats. This suggests that total grass production and livestock production may be favoured by the presence of such woody plants to certain density. Therefore, the use of such non-selective control methods in areas dominated by mature A. brevispica and A. drepanolobium may cause an undesirable shift in the yields of highly desirable and desirable grasses. If the intent is to obtain maximum grass yield and desired composition, bush control through selective thinning might be used in heavy stands of Acacias instead of attempting to clear completely.
Increased sub-canopy herbaceous production is often associated with leguminous shrubs, especially Acacia (Teague, 1984). Several hypotheses have been suggested to explain this phenomenon. Firstly, high grass production is assumed to be related to elevated soil nutrient status and improved physical (e.g., compaction) characteristics (Friedel, 1987) under tree canopies. Contributions from leaf litter (Ola-Adams and Egunjobi, 1992), bird droppings and dung of animals resting under trees (Teague and Smit, 1992) remains a possible source of soil enrichment. Nitrogen-fixation due to microbial activities in soil under leguminous trees has also been mentioned (Högberg, 1986). Decreased loss of nutrients to leaching or tighter recycling of nutrients by tree roots may also be important (Belsky et al., 1989). Microclimate changes (shading, cooling) are also known to increase herbaceous yield in some cases (Wilson et al., 1986). In contrast, in the USA, McPherson and Wright (1990) and Ansley and Rasmussen (2005) reported a severe negative impact of Juniperus sp. on the sub-canopy herbaceous and soil characteristics.
Secondly, stimulation of sub-canopy herbaceous production is assumed to be related to root and crown morphologies of woody plants. In this case, the most marked contrasts include those with tap roots, tall boles and elevated open crowns having positive or minor effects and those with lateral roots, short boles and low spreading crowns having the most negative effects (Coppock, 1994). Examples of diverse interactions among plants on the Borana rangelands include the apparently positive or neutral effects of A. tortilis (Forsk.) Hayne (Coppock, 1994) and A. drepanolobium (Tamene, 1990) on herbaceous layer as well as the negative effects of A. horrida (L.) Willd and A. seyal Del. (Coppock, 1994). In the current study, although the woody species have short (A. brevispica) and tall (A. drepanolobium) boles, extensive lateral and tap roots as well as low spreading crowns, they exhibit a positive effect on the sub-canopy grass yield. This implies that the contribution these plants make to soil enrichments would prevail over the negative or minor effects associated with root and crown morphologies. Indeed, the possible mechanisms observed in the current study require future investigation.
CONCLUSION
Significant differences were seen in the densities of A. brevispica and A. drepanolobium between the communal grazing land and a government ranch as well as between the near and furthest sites from water points. This is consistent with the differences in grazing and browsing pressure. Saplings had higher densities in sites subjected to higher grazing pressure. Dry matter yield for highly desirable and desirable grasses as well as for total grasses showed indications of higher production in the sub-canopy than open habitats. This study investigated the effect of mature isolated trees on their surrounding grass layer productivity. Indeed, research is required to determine the effect of smaller trees of these species on the productivity of the grass layer. Changes in total tree density may alter sub-canopy grass production and composition. Therefore, additional field and modelling experiments are needed to ascertain the extent to which cover and/or the density dynamics may alter grass layer productivity. Research is also required to identify determinants of increased grassland productivity in the sub-canopy of A. brevispica and A. drepanolobium.
REFERENCES
- Amarasekare, P., 2003. Competitive coexistence in spatially structured environments: a synthesis. Ecol. Lett., 6: 1109-1122.
CrossRef - Ansley, R.J. and G.A. Rasmussen, 2005. Managing native invasive juniper species using fire. Weed Technol., 19: 517-522.
CrossRefDirect Link - Angassa, A., 2005. The ecological impact of bush encroachment on the yield of grasses in Borana rangeland ecosystem. Afr. J. Ecol., 43: 14-20.
CrossRefDirect Link - Angassa, A. and R.M.T. Baars, 2000. Ecological condition of encroached and non-encroached rangelands in Borana Ethiopia. Afr. J. Ecol., 38: 321-328.
CrossRef - Belsky, A.J., 1994. Influence of trees on savanna productivity: Tests of shade, nutrients and tree-grass competition. Ecology, 75: 922-932.
CrossRefDirect Link - Belsky, A.J., R.G. Amundson, J.M. Duxbury, S.J. Riha, A.R. Ali and S.M. Mwonga, 1989. The effects of trees on their physical, chemical and biological environments in a semi-arid savanna in Kenya. J. Applied Ecol., 26: 1005-1024.
Direct Link - Belsky, A.J., S.M. Mwonga, R.G. Amundson, J.M. Duxbury and A.R. Ali, 1993. Comparative effects of isolated trees on their undercanopy environments in high- and low-rainfall savannas. J. Applied Ecol., 30: 143-155.
Direct Link - Chesson, P., 2000. Mechanisms of maintenance of species diversity. Ann. Rev. Ecol. Syst., 31: 343-366.
Direct Link - Childes, S.L. and B.H. Walker, 1987. Ecology and dynamics of the woody vegetation on the Kalahari sands in Hwange National Park, Zimbabwe. Plant Ecol., 72: 118-128.
Direct Link - Coppock, D.L., 1994. The Borana Plateau of Southern Ethiopia: Synthesis of Pastoral Research, Development and Change, 1980-91. ILCA. International Livestock Centre for Africa, Addis Ababa, Ethiopia, 9290532831.
Direct Link - Dalle, G., B.L. Maass and J. Isselstein, 2006. Encroachment of woody plants and its impact on pastoral livestock production in the Borana lowlands, Southern Oromia, Ethiopia. Afr. J. Ecol., 44: 237-246.
CrossRefDirect Link - Desta, S. and D.L. Coppock, 2002. Cattle population dynamics in the southern Ethiopia rangelands, 1980-97. J. Range Manag., 55: 439-451.
Direct Link - Hogberg, P., 1986. Nitrogen-fixation and nutrient relations in savanna woodland trees (Tanzania). J. Applied Ecol., 23: 675-688.
Direct Link - McPherson, G.R. and H.A. Wright, 1990. Effect of cattle grazing and Juniperus pinchottii canopy cover on herb cover and production in west Texas. Am. Midland Naturalist, 123: 144-151.
Direct Link - Moleele, N.M. and J.S. Perkins, 1998. Encroaching woody plant species and boreholes: is cattle density the main driving factor in the Olifants Drift communal grazing lands, South-eastern Botswana?. J. Arid Environ., 40: 245-267.
CrossRef - Oba, G., E. Post, P., P.O. Syvertsen and N.C. Stenseth, 2000. Bush cover and range condition assessments in relation to landscape and grazing in Southern Ethiopia. Landscape Ecol., 15: 535-546.
CrossRefDirect Link - Ola-Adams, B.A. and J.K. Egunjobi, 1992. Effects of spacing on litterfall and nutrient contents in stands of Tectona grandis Linn. F. and Terminalia superba Engl. and Diels. Afr. J. Ecol., 30: 18-32.
CrossRef - Riginos, C. and T.P. Young, 2007. Positive and negative effects of grass, cattle and wild herbivores on Acacia saplings in an African savanna. Oecologia, 153: 985-995.
CrossRef - Sankaran, M., J. Ratnam and N.P. Hanan, 2004. Tree-grass coexistence in savannas revisited-insights from an examination of assumptions and mechanisms invoked in existing models. Ecol. Lett., 7: 480-490.
CrossRef - Smit, G.N. and J.S. Swart, 1994. Influence of leguminous and non‐leguminous woody plants on the herbaceous layer and soil under varying competition regimes in mixed Bushveld. Afr. J. Range Forage Sci., 11: 27-33.
CrossRefDirect Link - Tainton, N.M., P.J. Edwards and M.T. Mentis, 1980. A revised method for assessing veld condition. Proc. Grassland Soc. Southern Afr., 15: 37-42.
CrossRefDirect Link - Solomon, T., H.A. Snyman and G.N. Smit, 2007. Rangeland dynamics of Southern Ethiopia: (2) Assessment of woody vegetation structure in relation to land use and distance from water in semi-arid Borana rangelands. J. Environ. Manage., 85: 443-452.
Direct Link - Vorster, M., 1982. The development of the ecological index method for assessing veld condition in the Karoo. Proc. Grassld Soc. Southern Afr., 17: 84-89.
CrossRefDirect Link - Wilson, J.R., V.R. Catchpoole and K.L. Weier, 1986. Stimulation of growth and nitrogen uptake by shading a rundown green panic pasture on brigalow clay soil. Trop. Grasslands, 20: 134-143.
Direct Link - Angassa, A. and F. Beyene, 2003. Current range condition in southern Ethiopia in relation to traditional management strategies: The perceptions of Borana pastoralists. Trop. Grasslds, 37: 53-59.
Direct Link