
M.M. Nemat Alla
Department of Botany, Faculty of Science at Damietta, Mansoura University, Damietta, Egypt
Y.M. Shabana
Department of Plant Pathology, Faculty of Agriculture, Mansoura University, Egypt
M.S. Serag
Department of Botany, Faculty of Science at Damietta, Mansoura University, Damietta, Egypt
N.M. Hassan
Department of Botany, Faculty of Science at Damietta, Mansoura University, Damietta, Egypt
M.M. El-Hawary
Department of Botany, Faculty of Science at Damietta, Mansoura University, Damietta, Egypt
Research Journal of Botany
Year: 2007 | Volume: 2 | Issue: 4 | Page No.: 165-175







ABSTRACT
An attempt was performed to achieve four granular formulations from two isolates of Fusarium oxysporum (Foxy I and Foxy II) for biocontrol of Orobanche ramosa and Orobanche crenata in tomato and faba bean; two formulations from microconidia (PM I and PM II) and two from chlamydospores (PC I and PC II). Doses of all formulations (0.5, 0.75, 1.0 and 1.25 g kg-1 soil) differentially reduced number of emerged shoots, shoot height and shoot dry weight of both Orobanche species with a remarkable increase in disease incidence on the parasitic plant, the highest dose giving the most severe symptoms. On the other hand, Orobanche-infected tomato and faba bean plants showed significant reductions in shoot height and root length as well as in shoot and root dry weights concomitant with significant increases in H2O2 content and superoxide dismutase (SOD) activity and significant decreases in ascorbic acid (AsA) content and activities of catalase (CAT), guaiacol peroxidase (GPX) and glutathione-S-transferase (GST). However, application of the highest dose of Pesta fungi overcame, to a great extent, the changes in host growthand oxidative stress parameters so that levels were similar to those of normally grown plants, the preparation PC II had the most pronounced effect. These findings showed that considerable control of the two species of Orobanche could be obtained by the formulations of the biocontrol agents, particularly at the highest dose (1.25 g kg-1 soil).
PDF Abstract XML References
How to cite this article
M.M. Nemat Alla, Y.M. Shabana, M.S. Serag, N.M. Hassan and M.M. El-Hawary, 2007. Granular Mycoherbicides Formulation of Fusarium oxysporum for Orobanche Biocontrol Mitigate Oxidative Stress and Growth Reduction in Host Species. Research Journal of Botany, 2: 165-175.
URL: https://scialert.net/abstract/?doi=rjb.2007.165.175
URL: https://scialert.net/abstract/?doi=rjb.2007.165.175
INTRODUCTION
Parasitic angiosperms are a taxonomically diverse group of plants that invade host plant tissues and remove resources via a specialized structure known as the haustorium. Through the haustorium, carbon, water and mineral nutrients are withdrawn, often at the expense of host growth and vigour (Watling and Press, 2001). The host selectivity of these plants is mediated by chemical signals, including germination stimulants and haustorial inducing factors (Shen et al., 2006). Parasitic weeds of the genus Orobanche represents a serious threat to a wide range of economically important crops. Orobanche crenata parasitizes major legume crops while Orobanche ramosa, which is closely related to aegyptiaca, attacks mainly Solanaceae (Rubiales et al., 2004). Due to infection with biotic agents such as Orobanche, oxidative stress could be arisen in host plants with a consequent production of Reactive Oxygen Species (ROS). Moreover, during O2 reduction, cells continuously produce ROS. They have been implicated in damaging cells. To protect cellular membranes against the harmful ROS levels, plants developed defense antioxidants (Aravind and Prasad, 2005). Antioxidants are crucial for plant defense against oxidative stress (Gomez et al., 2004). However, ROS accumulation is crucial to plant development as well as defense (Pavet et al., 2005). Because of the intimate host-parasite relationship and the anatomical-physiological connections, Orobanche is a particularly difficult target for selective chemical control (Kleifeld et al., 1998). Parasitic weeds are one of the major intractable regional problems and require biotechnological solutions (Gressel et al., 2004). Biological control using fungal pathogens could provide a possible solution because of the high specificity of the fungal pathogens used as biocontrol agents (Boari and Vurro, 2004; Elzein et al., 2006). Indigenous, weed-specific fungal pathogens can be developed and used as safe and effective bioherbicides (Charudattan, 2001). This study was an attempt to formulate granular Pesta from Fusarium oxysporum and to evaluate their role in both Orobanche biocontrol and mitigation of Orobanche-induced growth reduction and oxidative stress of host plants, tomato and faba bean.
MATERIALS AND METHODS
Plant Materials and Growth Conditions
This study was conducted in Botany Department, Faculty of Science at Damietta, Mansoura University, Egypt during the year 2006. Surface-disinfected seeds of tomato (Supper strain B) and faba bean (Giza 3) were used. Root exudate was produced by sowing ten seeds in plastic pots filled with 5 kg of sand. After 14 days, the root system was washed, grouped together and completely immersed in 200 mL water in a 250 mL glass flask. The flasks were then wrapped in black polyethylene. Plants were incubated for three days while water content of the flasks was held constant. Afterwards, plants were removed and the liquid content of the flasks was stored at -20°C until use.
Seeds of O. aegyptiaca/ramosa and O. crenata were germinated in vitro in order to determine the seed amounts needed to achieve a concentration of approximately 10,000 germinating seeds per pot (2 kg of soil). For the in vitro germination, surface-sterilized Orobanche seeds were placed on small glass fiber filter paper discs in petri dishes containing one layer of witted filter paper, sealed with parafilm and incubated for seven days at 20°C in the dark. The small discs were then transferred to other petri dishes with exudate. After ten days, the percentage of germinated seeds was determined using a binocular microscope.
For pot experiments, Orobanche seeds (60 mg to give 10000 seeds) were sprinkled onto the soil surface of plastic pots (25 cm diameter x 20 cm height) filled up to 2/3 of their height with 1:1 (v:v) sand:clay soil. Inoculum was added and mixed into the soil together with the seeds. The pots were then filled with soil and host plant seeds were sown. Fourteen days after sowing, plants were thinned out to three per pot. The pots were irrigated as required.
Granular Formulation of Mycoherbicides
About 300 infected Orobanche plants were collected from faba bean and tomato fields. Shoots were carefully collected and stored in a refrigerator. Small pieces of infected shoots were surface-disinfected, rinsed with sterile water, blotted dry and placed on Potato Dextrose Agar (PDA) plates supplemented with 200 ppm chloramphenicol and 100 ppm streptomycin sulfate. The plates were incubated at room temperature until fungal mycelium grew out of the plant pieces. The mycelium was transferred to fresh PDA plates to obtain pure cultures. One hundred and 69 fungal isolates obtained from infected shoots from infested fields were tested for their pathogenicity to Orobanche. Only two fungi isolated from diseased Orobanche shoots, classified as F. oxysporum isolate I (Foxy I) and isolate II (Foxy II), inhibited Orobanche germination. Microconidia and chlamydospores of both isolates were used to achieve prepare granular formulations for biocontrol of Orobanche. The most suitable formulations of Fusarial species for biocontrol of Orobanche are those that contain microconidia and chlamydospores as these remain viable in the soil for long periods and can re-germinate. Foxy isolates were grown on Potato Dextrose Broth (PDB) for six days on a reciprocating shaker at 125 strokes per min (spm) at room temperature to produce microconidia or for 20 days at 100 spm to produce chlamydospores. The contents of culture flasks were blended and the number of spores was determined using a haemocytometer. The resulting suspensions were centrifuged at 4,000 x g for 10 min and adjusted to 3.5x108 and 5x108 microconidia mL-1 for Foxy I and Foxy II, respectively and to 2.5x107 and 2.7x107 chlamydospores mL-1.
The two types of spores were used as active ingredients for the formulations along with three adjuvants (sucrose, yeast extract and glycerol). Dough was prepared by blending 38 g semolina, 4 g kaolin, 6 g yeast extract, 2 g sucrose, 20 mL spore suspension and 2 mL glycerol. The dough was then rolled through a pasta machine, folded and extruded several times. A 1 mm thick sheets were produced, air-dried at room conditions, ground and sieved. Four formulations resulted from both spore types of both isolates. Microconidia of Foxy I and Foxy II gave rise to PM I and PM II, chlamydospores gave rise to PC I and PC II.
Evaluation of Mycoherbicides Formulations
Plastic pots were filled up to 2/3 the height with sand/clay (1/1, v/v) soil and divided into groups. Sixty milligram of O. crenata or O. aegyptiaca/ramosa seeds together with the different fungal preparations at 0.0, 0.5, 0.75, 1.0 and 1.25 g kg-1 soil were well mixed with the sub layer. Five faba bean seeds were placed in O. crenata infested pots and covered with an additional 3 cm thick layer of soil. Faba bean seedlings were thinned to 3 per pot 14 days after sowing. The O. aegyptiaca/ramosa infested pots were planted with 3 tomato seedlings. Disease incidence on Orobanche and the number of emerged shoots were determined two months after planting. In addition, the Orobanche shoot height and shoot dry weight were determined. Meanwhile, an evaluation of host plants growth and oxidative stress signals was performed.
Determination of H2O2
Extraction was carried out in 200 mM perchloric acid and centrifuged at 5000 x g for 10 min. The supernatant was neutralized with 4 M KOH and centrifuged at 3000 x g for 5 min. An aliquot (0.2 mL) of the supernatant was loaded on 1 mL column of anion exchange resin and eluted with 0.8 mL of distilled water. H2O2 was assayed in 12.5 mM 3-dimethylaminobenzoic acid in 375 mM phosphate buffer pH 6.5, 1.3 mM 3-methyl-2-benzothiazolinone hydrazone and 0.25 units horseradish peroxidase (Okuda et al., 1991). The reaction was initiated by the addition of peroxidase and the increase in absorbance at 590 nm was monitored for 3 min.
Determination of Ascorbate (AsA)
Extraction was performed in 62.5 mM phosphoric acid, centrifuged at 12000x g for 20 min and filtered through a 0.5 μm Millipore filter. The filtrate was loaded onto an ion exclusion column (300x7.8 mm) connected to analytical HPLC system and eluted with 4.5 mM H2SO4 at a flow rate of 0.5 mL min-1. The elution of AsA was detected at 245 nm (Ahn et al., 1999).
Assays of Superoxide Dismutase (SOD), Catalase (CAT) Guaiacol Peroxidase (GPX) and Glutathione-S-Transferase (GST) Activities
All extraction steps were carried out at 4°C. SOD (EC 1.15.1.1) was extracted in 50 mM phosphate, pH 7.8, 0.1% (w/v) bovine serum albumin, 5.5 mM AsA and 8 mM β-mercaptoethanol. SOD was assayed in 50 mM phosphate, pH 7.8, 9.9 mM L-methionine, 0.057 mM nitroblue tetrazolium (NBT), 0.025% (w/v) Triton X-100 and 0.1 mM riboflavin by using the photochemical NBT method in terms of SOD’s ability to inhibit reduction of NBT to form formazan by superoxide (Beyer and Fridovich, 1987). The photoreduction of NBT was measured at 560 nm.
CAT (EC 1.11.1.6) was extracted in 50 mM phosphate buffer, pH 7 and 1 mM dithiothreitol. CAT was evaluated spectrophotometrically by determining the consumption of H2O2 at 240 nm in 50 mM phosphate buffer, pH 7.5 and 200 mM H2O2 (Aebi, 1984).
GPX (EC 1.11.1.7) was extracted in 220 mM Tris-HCl, pH 7.4, 250 mM sucrose, 50 mM KCl, 1 mM MgCl2, 160 mM β-mercaptoethanol and 0.57 mM phenyl methyl sulphonyl fluoride. GPX was assayed in 20 mM acetate, pH 5, 30 mM H2O2 and 2 mM guaiacol. The absorption at 470 nm was recorded and the activity was calculated using the extinction coefficient of 26.6 mM-1 cm-1 (Ranieri et al., 1997).
GST (EC 2.5.1.18) was extracted in 100 mM Tris-HCl, pH 7.5, 2 mM EDTA, 14 mM β-mercaptoethanol and 7.5% (w/v) polyvinylpolyprollidone, centrifuged at 15000 x g for 15 min, ammonium sulfate was added to 80% saturation and the protein pellets were collected (Dixon et al., 1995). GST was assayed in 100 mM phosphate, pH 6.5, 5 mM GSH and 1 mM CDNB. The absorbance at 340 nm was measured and the activity was calculated by the extinction coefficient E = 9.6 mM-1cm-1 (Ando et al., 1988).
Protein content was determined spectrophotometrically by reaction with Commassie Brilliant Blue G according to Bradford (1976). All values reported herein are means of at least six replications from two independent experiments. The full data were statistically analyzed using the Least Significant Difference (LSD) test at 5% level (Snedecor and Cochran, 1980).
RESULTS
Only two fungi isolated from diseased Orobanche shoots, classified as F. oxysporum isolate I (Foxy I) and isolate II (Foxy II), inhibited Orobanche germination. Microconidia and chlamydospores of both isolates were used for achieving formulations as Pesta (PM I and PM II from microconidia while PC I and PC II from chlamydospores). As shown in Table 1, the number of emerged and the total number of Orobanche shoots were decreased after the application of Pesta formulations.
| Table 1: | Effect of granular Pesta formulations from microconidia and chlamydospore (PM and PC) of two Fusarium oxysporum isolates (I and II) on emerged shoot number, shoots height, shoot dry weight and disease incidence in Orobanche ramosa and Orobanche crenata |
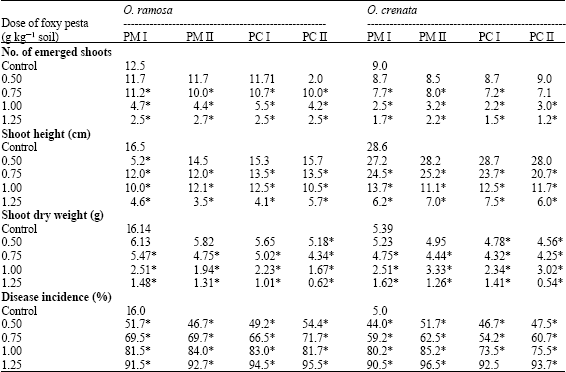 | |
Values are mean of at least six determinations from two independent experiments. *: Values are significantly different at 5% level with respect to untreated control | |
However, low doses had, in general, no significant effects on the number of emerged shoots of both Orobanche species. Increasing Pesta dose retarded Orobanche emergences so that high doses resulted in much diminutions in emerged shoots. Moreover, Pesta significantly reduced shoot height of both Orobanche species compared with their respective controls. The magnitude of reduction increased with increasing doses. In addition, Orobanche shoot dry weight was significantly decreased by all doses of the applied Pesta formulations, the magnitude of decrease was greatest with the highest dose. Nevertheless, PC II seemed to be the most effective formulation. The trend of response to the formulations was most likely similar in both Orobanche species. The emerged shoots of the Pesta-treated Orobanche exhibited disease symptoms at emergence, which continued up to the flowering stage. Disease symptoms consists of inhibition of Orobanche germination, wilting, necrosis and shoot curvature of those plants that did emerge. High doses mostly prevented flowering and fruit production of Orobanche. Treatments with either of the different formulations highly enhanced Disease Incidence (DI) of the emerged Orobanche shoots either with faba bean or with tomato. DI appeared to be mostly related to the doses of formulations. Treatments with either of the different formulations highly enhanced Disease Incidence (DI) of the emerged Orobanche shoots either with faba bean or with tomato. DI appeared to be mostly related to the formulation doses.
Regarding growth parameters of host plants, Table 2 shows that Orobanche reduced shoot height, root length as well as shoot and root dry weights of tomato and faba bean as compared with normally grown plants. However, the Orobanche-induced reduction in host growth was ameliorated by application of Foxy Pesta formulations. Shoot height and root length of Orobanche-infected tomato and faba bean seemed to be mitigated by all types of formulations to become comparable to control levels.
| Table 2: | Effect of granular Pesta formulations from microconidia and chlamydospore (PM and PC) of two Fusarium oxysporum isolates (I and II) on shoot height and root length as well as shoot and root dry weight of tomato and faba bean infected with Orobanche ramosa and Orobanche crenata, respectively |
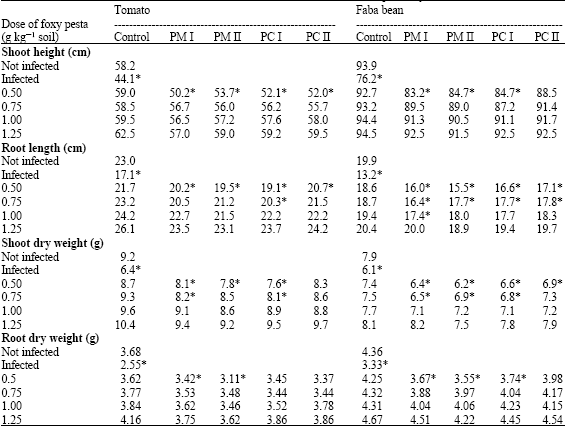 | |
Values are mean of at least six determinations from two independent experiments. *: Values are significantly different at 5% level with respect to untreated control | |
| Table 3: | Effect of granular Pesta formulations from microconidia and chlamydospore (PM and PC) of two Fusarium oxysporum isolates (I and II) on ascorbate (AsA) and H2O2 contents in shoots and roots of tomato and faba bean infected with Orobanche ramosa and Orobanche crenata, respectively |
 | |
Values are mean of at least six determinations from two independent experiments. *: Values are significantly different at 5% level with respect to untreated control | |
| Table 4: | Effect of granular Pesta formulations from microconidia and chlamydospore (PM and PC) of two Fusarium oxysporum isolates (I and II) on activity of superoxide dismutase (SOD) in shoots and roots of tomato and faba bean infected with Orobanche ramosa and Orobanche crenata, respectively |
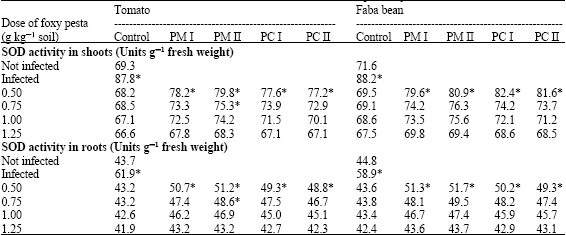 | |
Values are mean of at least six determinations from two independent experiments. *: Values are significantly different at 5% level with respect to untreated control | |
The effect of PC II was greater compared to the other formulations. Similar increases were also detected in shoot and root dry weights. Such reduction in symptoms became more pronounced at higher doses of biocontrol formulations and plants treated with the highest doses were indistinguishable from controls.
The H2O2 content in shoots and roots of the Orobanche-infected tomato and faba bean plants were significantly higher than in the control (Table 3). The application of Pesta formulations reduced the magnitude of the Orobanche-induced accumulation. The reduction in H2O2 accumulation was greater with higher doses of the formulations and the PC II formulation was the most effective.
In contrast to H2O2, there was a significant drop in the contents of AsA in tomato and faba bean plants due to Orobanche infection. The application of all formulations resulted in significant increases in AsA content of the infected plants to reach mostly the contents of the healthy non-infected plants. The magnitude of increase in AsA content was proportional to the increase in Pesta doses; the higher the Pesta dose was, the greater was the enhancement of AsA content.
| Table 5: | Effect of granular Pesta formulations from microconidia and chlamydospore (PM and PC) of two Fusarium oxysporum isolates (I and II) on activity of catalase (CAT), guaiacol peroxidase (GPX) and glutathione-S-transferase (GST) in shoots and roots of tomato and faba bean infected with Orobanche ramosa and Orobanche crenata, respectively |
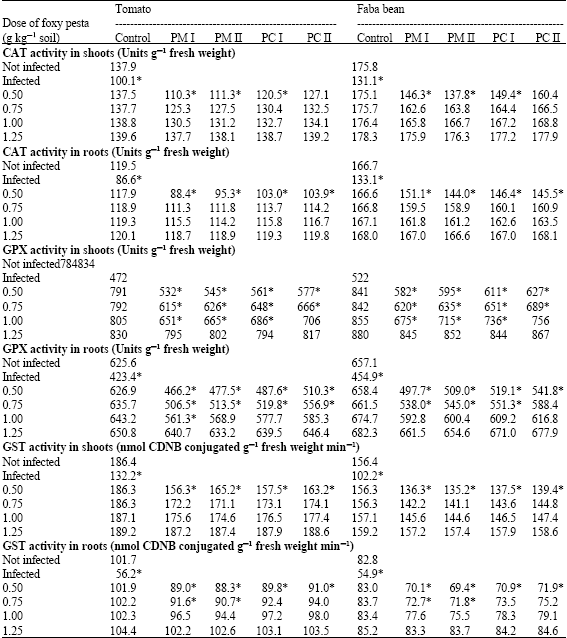 | |
Values are mean of at least six determinations from two independent experiments. *: Values are significantly different at 5% level with respect to untreated control | |
SOD activity was significantly higher in shoots and roots of the Orobanche infected tomato and faba bean plants than in the control plants (Table 4). The application of Pesta formulations significantly decreased the enhancement of SOD activity. The reduction of theinduced increases in SOD activity was related to dosage of the formulations and at the higher doses values were not significantly different from uninfected control plants.
Infection with Orobanche led to decreases in activities of CAT, GPX and GST in shoots and roots of tomato and faba bean plants as compared to the control plants (Table 5). Pesta application overcame the infection-induced inhibition of the enzyme activity. The release of the enzymes activities from inhibition was related to the dose of the Pesta formulations and became comparable with those of controls at the higher levels.
DISCUSSION
In the present study, two isolates of F. oxysporum showed pathogenicity to O. crenata and O. aegyptiaca/ramosa. Microconidia and chlamydospores of both isolates were used to achieve granular mycoherbicide formulations for biocontrol of Orobanche. Severe reductions were obtained in shoot emergence of O. crenata and O. ramosa, respectively by application of all Foxy Pesta formulations particularly at the highest dose. Similar reductions were observed in shoot height and shoot dry matter. On the contrary, disease incidence of Orobanche was highly accelerated upon application of Pesta. In this connection, Shabana et al. (2003) reported that application of Pesta formulations containing microconidia and chlamydospores of F. oxysporum f. sp. orthoceras (FOO) as a bioherbicide for O. cumana resulted in a reduction in Orobanche biomass and increase in disease severity. Moreover, Pesta granules reduced the emergence of O. cumana shoots (Müller-Stöver et al., 2004). Similar results were also observed by Müller-Stöver and Kroschel (2005) on O. crenata by inoculation with Ulocladium botrytis. The reduction of Orobanche growth by Foxy isolates may be due to the production of toxic metabolites (Zonno and Vurro, 2002; Müller-Stöver and Kroschel, 2005).
Growth of the host species (faba bean and tomato) was significantly reduced by Orobanche infection. The Orobanche-induced growth inhibition of tomato and faba bean due to parasitism might result from depletion of host nutrition. However, these reductions appeared to be counterbalanced following the application of the formulated Pesta. The biocontrol agents did not only prevent germination but also attacked developed shoots overcoming, therefore, any malfunction in host metabolism probably caused by Orobanche infection. Consequently a repair in host metabolism might support new syntheses and consequently normal and even vigor growth. In this context, Müller-Stöver and Kroschel (2005) found that dry matter accumulation of faba bean was significantly increased by treatment with U. botrytis. Also, Shabana et al. (2003) observed an increase in sunflower dry weight as a result of treatments with FOO. Therefore, Pesta not only overcame the Orobanche-induced reductions in host growth but also seemed to serve as amendments, to some extent, supplying host species with good conditions for growth. The inhibition in Orobanche growth following Pesta application accompanied with recoveries in host growth could point to the suitability of the formulations as mycoherbicides for Orobanche biocontrol with no negative side-effects on host growth.
The stress imposed in tomato and faba bean by Orobanche could generate oxidative stress resulted from both infection and normal growth, a state that could be indicated by the accumulation of H2O2 and the decline of AsA. Aerobic organisms depend on O2 as electron acceptor in electron transfer reactions. During O2 reduction, cells continuously produce ROS (Mittler, 2002). ROS are involved in molecular damage in plants (Noctor and Foyer, 1998). Aerobic organisms evolve numerous antioxidants to minimize the adverse effects of ROS (Durmus and Kadioglu, 2005; Hassan and Nemat Alla, 2005; Nemat Alla et al., 2007). The accumulation of H2O2 and the decline of AsA could be considered as indices of oxidative stress. However, H2O2 accumulation is also very important to trigger defense mechanisms against pathogens. ROS accumulation is crucial to plant development as well as defense (Foyer and Noctor, 2005; Pavet et al., 2005). Signals of oxidative stress could be evidenced from the abnormal changes in activities of antioxidative stress enzymes (Nemat Alla and Hassan, 2007; Nemat Alla et al., 2007). Within a cell, SOD constitutes the first line of defense against ROS (Alscher et al., 2002). SOD dismutates the superoxide radicals into H2O2 (Noctor and Foyer, 1998). H2O2 is detoxified by GPX or CAT and sometimes by GST which may act as peroxidase. However, under unstressed conditions, the formation and removal of ROS are in balance. Therefore, the defense system, with increased ROS formation under stress conditions, can be overwhelmed. Detoxification reactions must involve right balance between the formation and detoxification of ROS.
The increases in SOD activity of the Orobanche infected plants concomitant with the inhibited activities of CAT, GPX and GST could explicate the great accumulation of H2O2. Thus, the accumulated H2O2 might arise from dismutation of superoxide radical with no enhanced detoxification routes. Under these conditions where CAT, GPX and GST were inhibited, the cell is not competent to scavenge H2O2. On the other hand, application of Pesta formulation overcame the stress inducers and consequently could terminate and moreover eliminate the stress circumstances. Therefore, AsA content and activities of CAT, GPX and GST showed enhancements supporting more detoxification of H2O2 with a subsequent decline in its accumulation. AsA increases H2O2 detoxification through a played role by the AsA-glutathione (AsA-GSH) cycle. This cycle is an important and powerful detoxifying mechanism in the plant cells (Ma and Cheng, 2003) in which H2O2 is reduced to water by peroxidase on the expense of AsA oxidation (Mittler, 2002; Aravind and Passad, 2005; Nemat Alla et al., 2007). AsA is regenerated again by monodehydroascorbate reductase and GSH-dependent dehydroascorbate reductase coupled with glutathione reductase to maintain GSH levels for elimination of ROS (Nagalakshmi and Prasad, 2001). These antioxidants are crucial for plant defense against oxidative stress (Gomez et al., 2004; Nemat Alla and Hassan, 2007). In addition, CAT, GPX and GST help in H2O2 scavenge by catalyzing its reduction to water (Nagalakshmi and Prasad, 2001; Nemat Alla and Hassan, 2006; Nemat Alla et al., 2007).
The present results clearly revealed that application of Pesta formulations particularly at 1.25 g kg-1 soil to Orobanche infected plants retarded Orobanche growth and simultaneously retracted the Orobanche-induced reductions in host plants and moreover delayed oxidative stress. These findings could suggest that an increase in the defense of host plants was developed by Pesta application against the stress elicited by the infection. Antioxidative mechanism seemed to be modified as activities of SOD, CAT, GPX and GST as well as contents of AsA were greatly improved following the application of Pesta formulations. As a result, H2O2 and certainly other ROS could be eliminated. Therefore, it could be suggested that these formulations have positive influences not only because of overcoming the infection effect but also of improving plant metabolites and antioxidative defense mechanism.
The present findings, moreover, could suggest a high virulence of chlamydospores of isolate II in performing the Pesta against Orobanche. Anyway, H2O2 levels and SOD activity were decreased by Pesta to reach those of normally grown non-infected plants. In the mean time, AsA content and activities of CAT, GPX and GST raised. The retractions in oxidative stress of host plants by Foxy Pesta in addition to the recoveries of growth reductions could suggest that plants seemed more healthy to withstand even with the presence of Orobanche. These findings conclude that the application of Foxy formulations particularly PC II at 1.25 g kg-1 soil seemed to scavenge the negative effects of parasitism and appeared efficient to mitigate oxidative stress and growth reduction in tomato and faba bean resulted from Orobanche infection.
REFERENCES
- Alscher, R.G., N. Erturk and L.S. Heath, 2002. Role of superoxide dismutases (SODs) in controlling oxidative stress in plants. J. Exp. Bot., 53: 1331-1341.
Direct Link - Aravind, P. and M.N.V. Passad, 2005. Modulation of cadmium-induced oxidative stress in Ceratophyllum demersum by zinc involves ascorbate-glutathione cycle and glutathione metabolism. Plant Physiol. Biochem., 43: 107-116.
Direct Link - Boari, A. and M. Vurro, 2004. Evaluation of Fusarium spp. and other fungi as biological control of broomrape (Orobanche ramose). Biol. Control, 30: 212-219.
Direct Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Durmus, N. and A. Kadioglu, 2005. Spermine and putrescine enhance oxidative stress in maize leaves. Acta Physiol. Plant., 27: 515-522.
CrossRefDirect Link - Elzein, A., J. Kroschel and V. Leth, 2006. Seed treatment technology, An attractive delivery system for controlling root parasitic weed Striga with mycoherbicide. Biocontrol Sci. Tech., 16: 3-26.
CrossRefDirect Link - Foyer, C.H. and G. Noctor, 2005. Redox homeostasis and antioxidant signaling: A metabolic interface between stress perception and physiological responses. Plant Cell, 17: 1866-1875.
CrossRefDirect Link - Gomez, L.D., G. Noctor, M.R. Knight and C.H. Foyer, 2004. Regulation of calcium signaling and gene expression by glutathione. J. Exp. Bot., 55: 1851-1859.
CrossRefDirect Link - Gressel, J., A. Hanefi, G. Head, W. Mareses and A.B. Obilane et al., 2004. Major heterofore intractable biotic constraints to African food security that may be amenable to novel biotechnological solutions. Crop Prot., 23: 661-689.
Direct Link - Hassan, N.M. and M.M. Nemat Alla, 2005. Oxidative stress in herbicide-treated broad bean and maize plants. Acta Physiol. Plant, 27: 429-438.
Direct Link - Ma, F. and L. Cheng, 2003. The sun exposed peel of apple fruit has higher xanthophylls cycle dependent thermal dissipation and antioxidants of the ascorbate glutathione pathway than shaded peel. Plant Sci., 165: 819-827.
Direct Link - Mittler, R., 2002. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci., 7: 405-410.
CrossRefPubMedDirect Link - Stover, D.M, H. Thomas, J. Sauerborn and J. Kroschel, 2004. Two granular formulations of Fusarium oxysporum f. sp. orthoceras to mitigate sunflower broomrape Orobanche cumana. Biocontrol, 49: 595-602.
CrossRef - Nagalakshmi, N. and M.N.V. Prasad, 2001. Responses of glutathione cycle enzymes and glutathione metabolism to copper stress in Scenedesmus bijugatus. Plant Sci., 160: 291-299.
CrossRefPubMedDirect Link - Nemat Alla, M.M. and N.M. Hassan, 2007. Changes of antioxidants and GSH associated enzymes in isoproturon treated maize. Acta Physiol. Plant., 29: 247-258.
CrossRef - Nemat Alla, M.M., A.M. Badawi, N.M. Hassan, Z.M.E. Bastawisy and E.G. Badran, 2007. Induction of glutathione and glutathione associated enzymes in butachlor tolerant plant species. Am. J. Plant Physiol., 2: 195-205.
CrossRef - Noctor, G. and C.H. Foyer, 1998. Ascorbate and glutathione keeping active oxygen under control. Annu. Rev. Plant Physiol. Plant Mol. Biol., 49: 249-279.
PubMed - Okuda, T., Y. Masuda, A. Yamanaka and S. Sagisaka, 1991. Abrupt increase in the level of hydrogen peroxide in leaves of winter wheat is caused by cold treatment. Plant Physiol., 97: 1265-1267.
Direct Link - Pavet, V., E. Olmos, G. Kiddle, S. Mowla and S. Kumar et al., 2005. Ascorbic acid deficiency activates cell death and disease resistance responses in Arabidopsis. Plant Physiol., 139: 1291-1303.
CrossRef - Ranieri, A., A. Castagna, G. Lorenzini and G.F. Soldatini, 1997. Changes in thylakoid protein patterns and antioxidant levels in two wheat cultivars with different sensitivity to sulphur dioxide. Environ. Exp. Bot., 37: 125-135.
CrossRef - Rubiales, D., C. Alcantara and J. Sillero, 2004. Variation in resistance to Orobanche crenata in species of Cicer. Eur. Weed Res. Soc., Weed Res., 44: 27-32.
Direct Link - Shen, H. , W. Ye, L. Hong, H. Huang, Z. Wang, X. Deng, Q. Yang and Z. Xu, 2006. Progress in parasitic plant biology host selection and nutrient transfer. Plant Biol., 8: 175-185.
PubMed - Watling, J.R. and M.C. Press, 2001. Impacts of infection by parasitic angiosperms on host photosynthesis. Plant Biol., 3: 244-250.
Direct Link - Zonno, M.C. and M. Vurro, 2002. Inhibition of germination of Orobanche ramosa seeds by fusarium toxins. Phytoparasitica, 30: 519-524.
CrossRef - Shabana, Y.M., D. Muller-Stover and J. Sauerborn, 2003. Granular pesta formulation of Fusarium oxysporum f. sp. Orthoceras for biological control of sunflower broomrape: Efficacy and shelf life. Biol. Control, 26: 189-201.
CrossRef