
Adillah Ibrahim
Department of Biosciences, Faculty of Science, University of Mauritius, 80837 Reduit, Mauritius
Nawsheen Taleb-Hossenkhan
Department of Biosciences, Faculty of Science, University of Mauritius, 80837 Reduit, Mauritius
LiveDNA: 222.18548
Plant Pathology Journal
Year: 2017 | Volume: 16 | Issue: 3-4 | Page No.: 121-129







ABSTRACT
Background and Objective: Control of late blight disease of potato and tomato, caused by the oomycete Phytophthora infestans (P. infestans), remains a highly challenging task in almost all potato and tomato growing areas around the world. This is partly due to the high degree of polymorphism in this pathogen’s genome, which has been reported to rapidly evolve via host jumps followed by specialization. Late blight remains a major problem in Mauritius where fungicide use is less common on potato and certified seed programs are not well established. This study aimed at investigating the genetic identity of P. infestans isolates in Mauritius collected during the late blight seasons from 2012-2014 using mitochondrial haplotyping, PCR-based mating type analysis and random amplified polymorphic DNA (RAPD). Materials and Methods: Seven isolates were collected from distinct geographical areas around the island, cultured by baiting with potato disks and subsequently isolated on antibiotic-amended pea agar for further analysis. Mitochondrial haplotyping was carried out using four previously characterized polymorphic regions of the mitochondrial genome, mating type analysis was carried out using a CAPS assay and RAPD fingerprinting was carried out using 40 random oligomers. Results: All of the isolates were shown to belong to mitochondrial type II and mating type A2 and no A1 mating type was detected. Codified data from RAPD fingerprinting was used to construct a consensus dendogram using the UPGMA method, which grouped isolates into 3 distinct clusters but all isolates were shown to be genetically distinct from each other and there was no indication of geographical clustering. Conclusion: These results reveals a much wider diversity among the existing populations of P. infestans found on the island than was previously thought. It infers that blight isolates may be gaining access to the island from several different geographical sources and underlines the importance of stricter screening procedures on imported seed potato and tomato being imported onto the island.
PDF Abstract XML References Citation
Received: August 28, 2017;
Accepted: October 09, 2017;
Published: March 01, 2018
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Adillah Ibrahim and Nawsheen Taleb-Hossenkhan, 2017. Genotypic Characterization of Phytophthora infestans from Mauritius using Random Amplified Polymorphic DNA (RAPD), Mitochondrial Haplotyping and Mating Type Analysis. Plant Pathology Journal, 16: 121-129.
DOI: 10.3923/ppj.2017.121.129
URL: https://scialert.net/abstract/?doi=ppj.2017.121.129
DOI: 10.3923/ppj.2017.121.129
URL: https://scialert.net/abstract/?doi=ppj.2017.121.129
INTRODUCTION
Late blight has been reported in Mauritius since the 1970’s and the latest devastating outbreak occurred in 2004, with disastrous economic consequences for the production of potato on the island. With an annual consumption of around 23,000 t, Mauritius aims at achieving self-sufficiency in potato production but pests and diseases remain one of the major obstacles to overcome and late blight ranks among the most important potato diseases reported by planters each year during the winter season. Phenotypic markers like specific virulence phenotypes on plants and fungicide sensitivity have been used to characterize populations of P. infestans. However, since phenotypic markers are under selection pressure and are thus not neutral markers, their use in analysis of P. infestans populations is limited. P. infestans isolates can be discriminated using relatively few reliable markers1. Allozyme alleles at glucose 6-phosphate isomerase (Gpi-1) and peptidase (Pep-1) loci have been useful codominant genetic markers for P. infestans population characterization. Selectively neutral markers and mating type are used in studies of P. infestans as they can help to discriminate between sexual and asexual populations2. Research on the genetic structure from the Del Fuerte Valley has shown that data on metalaxyl sensitivity indicate that there is a correlation between metalaxyl sensitivity and allozyme markers and RFLP analysis with the probe RG57 can provide further discrimination between genotypes within an allozyme genotype2.
A global marker database exists for P. infestans where information on mitochondrial DNA (mtDNA) haplotypes, allozyme genotypes and fungicide sensitivity are compiled3. The database contains information for isolates of potato and tomato late blight pathogen originating from 41 locations and contains information for 1,776 for one or more of the following markers: Restriction fragment length polymorphism (RFLP) “fingerprint”, mating type, dilocus allozyme genotype mitochondrial DNA haplotype, sensitivity to the fungicide metalaxyl and virulence4. Isolates in each country were analysed by a multilocus genotype approach and were given a code based on the International Organization for Standardization (ISO) two-letter code and a unique number. Most of the genetic diversity represented in the database is associated with North America and Europe and this partly reflects the global structure of this important plant pathogen. The presence of both mating types and associated sexual recombination in Mexico and Europe has also contributed to a large number of genotypes, found in those areas4. The database also shows a geographic imbalance in research activity because very little sampling and marker analyses have been done outside North America, Europe and a few locations in Asia. Additional information on the P. infestans populations from Africa, South America and Asia could assist research in the development of improved disease management strategies as information obtained from pathogen population studies in the past has led to recommendations for refinement or modification of disease management approaches. Knowledge of pathogen population structure can also play a key role in selection and deployment of durable host resistance.
Two major migration events are thought to account for the worldwide spread of P. infestans. In the first migration, it was suggested that the pathogen migrated in the mid 19th century from Mexico to the US and then to Europe. The pathogen population in central Mexico is the only one in which more than 25 bands have been identified by the DNA fingerprint profiles using RFLP probe RG57 and 4 and 7 alleles at the allozyme loci peptidase (Pep) and Glucose-6-phosphate isomerase (Gpi), respectively. However recent analysis strongly supports a South American (Andean) origin5. Phytophthora infestans first appeared in Europe in the 1840’s, probably imported in seed tubers6. It is believed that a single strain of the pathogen (US-1) migrated from Mexico to Europe via North America in the early 1840’s and it was suggested that isolates derived from the US-1 genotype was the only genotype introduced to most locations around the world7,8. A second migration occurred out of Mexico to Western Europe in the late 1970’s7 which brought new genotypes of both A1 and A2 mating types to Europe9. The new genotype spread quickly and by the 1980’s the A2 mating type was present in almost all locations in Europe, this new genotype displaced the old clonal lineage in Europe and proved to be more aggressive with greater fitness than the old lineage6,10. The old population consisted exclusively of the A1 mating type and Ib mitochondrial haplotype, whereas the new population comprised of isolates of both A1 and A2 mating types and Ia and IIa mitochondrial haplotypes as well as previously undetected allozyme genotypes and a much greater diversity of DNA fingerprints8.
Late blight was first reported in the continent of Africa in 194111. Limited knowledge is available on the existing P. infestans populations in Sub Saharan African (SSA) countries. It has been reported that the US-1 lineage (mating type A1) is the dominant lineage in SSA countries including Tanzania, Kenya, Uganda, Rwanda, Burundi, Malawi, Mozambique and South Africa and no mating type A2 has been detected in those areas yet but the situation is different in Northern African countries like Ethiopia, where the US-1 lineage seems to have been replaced by a population characterised by mitochondrial haplotype Ia12,13. It was concluded that it is unlikely that the more virulent clonal genotypes of Northern Africa may have reached the SSA countries. In Mauritius the pathogen was discovered in 1937 and one isolate collected in 1999 and characterized at Cornell University showed that it belonged to the US-1 lineage. However, isolates collected in the late blight seasons of 2012 and 2013 exhibited a wide range of allozyme patterns at both the Gpi and Pep loci14, clearly showing a wide diversity of prevailing strains that now exist on the island. This study aimed at investigating the mitochondrial haplotype, mating type and RAPD profiles of these isolates in order to get a more comprehensive picture of the genetic identity of isolates from Mauritius.
MATERIALS AND METHODS
Sample collection, isolation and maintenance: Isolates were collected from infected potato fields over seven geographical locations during the blight seasons of 2012, 2013 and 2014. This study spans over 3 years because late blight outbreaks mostly go unreported on the island and the number of isolates collected from reported outbreaks can therefore be relatively small. Therefore, in order to get a sufficient number of isolates to provide scientific validity to the study, isolates were collected over 3 years. This provides absolutely novel data as no previous data is available on the genotypic characteristics of P. infestans from this part of the world. The locations included the East (Bambous), the Central Plateau (Saint Pierre, Glen Park, La Marie), the South West (Nouvelle France) and South (Mare Longue, Plaine Sophie) of the island. The isolates were coded following the location from where they were isolated as follows: LM, BB, GP, SP, NF, ML and PS. For each isolate, P. infestans infected leaves from potato fields were collected and placed in a closed plastic bag with a moist piece of cotton wool which was then placed at 18°C overnight to favor sporulation. Small areas showing sporulating lesions were then placed under potato slices of known susceptible potato cultivars in a petri dish. Dishes were incubated at 18°C for 1 week, until abundant sporulation could be observed on the surface of the potato slices. The mycelium was then transferred to pea agar amended with an antibiotic mix (Vancomycin, Polymixin B, Ampicillin, Rifampicin, PCNB, Benlate). Isolates were subsequently maintained by sub-culturing on pea agar with the same composition.
DNA extraction: The DNA was extracted using the GeneJet Plant Genomic DNA Purification kit (Thermo Fisher Scientific, Waltham, Massachusetts, United States) following the manufacturer’s instructions. One hundred mg of fresh mycelium tissue was used and the final elution was performed using 100 mL of elution buffer. The genomic DNA extracted was then analyzed on a 1% agarose gel in TBE buffer. DNA samples were subjected to spectrophotometric analysis (JP Selecta UV Spectrophotometer Model UV-2005, Abrera, Barcelona, Spain) at 260 and 280 nm to determine yield and purity. Triplicate samples were subsequently used for mitochondrial haplotyping, mating type analysis and RAPD analysis to ensure reproducibility and viability of results.
Mitochondrial haplotyping: Mitochondrial haplotypes were determined by amplification of DNA of each isolate using primers designed for four specific polymorphic regions of the mitochondrial genome of P. infestans following the original protocol designed by Griffith and Shaw15. After amplification with primer sets F1/R1, F2/R2, F3/R3 and F4/R4, bands corresponding to the correct amplicon size were excised using a ZymocleanTM Gel DNA Recovery Kit (ZYMO RESEARCH, Irvine, California, United States) before being subjected to restriction digestion with enzymes Cfo I, Msp I and EcoRI, which yielded banding patterns used for classification into mtDNA haplotypes Ia, Ib, IIa and IIb. Type II differs from Type I by possessing an extra 1kb insert of DNA. Type I may be further differentiated into Ia and Ib, the latter possessing an extra MspI restriction site. Type II can be differentiated into Type IIa and IIb with IIb possessing an extra Cfo1 site.
Mating type analysis: Mating type analysis was performed using a PCR-based assay using CAPS markers (Cleaved Amplified Polymorphic Sequences) previously identified by Judelson et al.16 following the method devised by Mazakova et al.17, with slight modifications as follows: 2 μL of template DNA were used, the number of cycles was increased from 29-35 and an annealing temperature of 55°C was used. Fragments from both mating types (A1 and A2) were amplified using the W16-1 (5 -ACACGACAACGATATAAATGTA-3 and W16-2 (5 GCGTAATGTAGCGTAACAGCTCTC-3 ) primers, which amplify sequences linked to the mating type locus, in the P. infestans genome. Subsequent digestion with the restriction enzyme BsuR I (Hae III) allows differentiation between mating type I and II.
Rapid Amplified Polymorphic DNA (RAPD) analysis: To estimate the genetic diversity of P. infestans isolates collected from 2012-2014, the extracted DNA from the isolates were subjected to rapid amplified polymorphic DNA (RAPD) analysis. RAPD primers were selected by screening genomic DNA using four sets of ten random decamer primers:
OPA 1-10, OPB 1-10, OPE 1-10 and OPL 1-10 (Operon Technologies Inc., Alameda, California, United States) to identify primers that exhibited polymorphisms and yielded consistent, easily scorable banding patterns. The DNA amplification was performed in the Thermo Fisher Scientific ProFlex™ PCR System (Thermo Fisher Scientific, Waltham, Massachusetts, United States with one cycle of 98°C for 2 min, followed by 35 cycles of 98°C for 30 sec, 35°C for 30 sec and 72°C for 1 min and a final cycle at 72°C for 5 min. Reactions were carried out in 25 μL volumes containing 1XDNA polymerase Buffer, 0.2 mM of each dNTP, 2.0 μM primer, 0.2 U Taq DNA polymerase and 5 μL of DNA template at concentration 30-50 ng μL–1. Amplification products were separated in a 1% agarose gel, visualized under 302 nm UV light and photographed. RAPD bands were manually scored and analyzed as binary data, with 1 representing the presence and 0 representing the absence of a band at a particular location in each lane. Bands were assumed to be independent and those of identical size were assumed to have identical sequences.
Statistical analysis: The data was analyzed with the computer software NCSS v10. Hierarchical clustering was performed using the unweighted pair-group method with arithmetic averages (UPGMA) option for linkage type and the Euclidean option for distance method and dendrograms showing the relationships among the RAPD patterns were then generated from these matrices.
RESULTS
Mitochondrial haplotyping: High-molecular weight DNA was successfully obtained from all seven isolates. For all amplifications carried out, the amplicons corresponding to the correct sizes were excised from the agarose gel and purified before being subjected to restriction digest with the appropriate restriction enzyme (Fig. 1). Amplification with primer set F1/R1 yielded the expected 1118 bp amplicon (P1), which was digested with Cfo I. Amplification with primer set F3/R3 yielded the expected 1308 bp amplicon (P3), which was digested with EcoR I. Amplification with primer set F4/R4 yielded the expected 964 bp amplicon (P4), which was digested with EcoR I. Amplification with primer set F2/R2 did not yield results. The RFLP results all indicated that the seven tested DNA samples belonged to mitochondrial haplotype II: The P1 and the P3 products remained undigested after an overnight incubation with Cfo I and EcoR I, respectively and P4 product yielded fragments of 350 and 600 bp, respectively (Fig. 2).
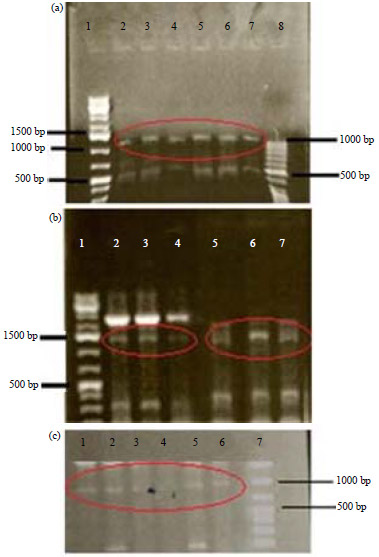 | |
| Fig. 1(a-c): | PCR products for mitochondrial haplotyping, (a) PCR amplified products obtained with primer pair F1/R1: Lane 1: 1 Kb ladder, Lane 2-8: DNA fragments of size of about 1118 bp obtained for isolates LM, BB, GP, SP, NF, ML, PS, (b) PCR results for amplification of P3 region of the mitochondrial genome with primer set F3/R3: Lane 1: 1 Kb ladder, Lane 2 to 7: DNA fragment of size of about 1308 bp obtained with isolates LM, BB, GP, SP, NF, ML and (c) PCR profile after amplification of P4 regions with primer set F4/R4: Lane 1 - 6: products of size of about 964 bp obtained with isolates LM, BB, GP, SP, NF, ML, Lane 7: 1 Kb ladder |
Mating type analysis: This assay generates 600 bp bands from both mating types and upon digestion with Hae III, the A1 mating type generates two bands of 550 and 600 bp, respectively, as the A1 mating type is heterozygote for this marker and the A2 mating type generates a single 550 bp band, as the A2 mating type is homozygote for this marker.
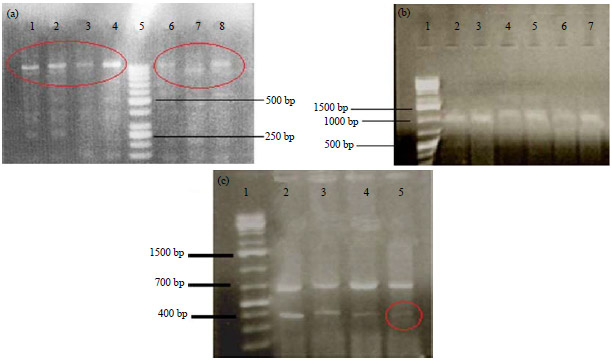 | |
| Fig. 2(a-c): | PCR-RFLP results for mitochondrial haplotyping, (a) Restriction profile of P1 region with enzyme CfoI: Lane 1-4: Undigested products of size of about 1118 bp with isolates LM, BB, GP, SP, Lane 5: 50 bp ladder, Lanes 6-8: Fragments of size of about 1118 bp for isolates NF, ML, PS, (b) Restriction profile of P3 region with enzyme EcoR I: Lane 1: 1 Kb ladder, Lanes 2-7: Undigested products of size of about 1308 bp with isolates LM, BB, GP, SP, NF, ML and (c) Restriction profile of P4 region with enzyme EcoR I: Lane 1: 1 Kb ladder, Lanes 2-5: Fragments of about 361 bp and 603 bp for isolates LM, BB, GP, SP |
 | |
| Fig. 3(a-b): | PCR products for mating type analysis, (a) PCR product from amplification using W16-1/W16-2 primer pairs, Lane 1: 1 Kb ladder, Lanes 2-3: Band of size 702 bp obtained for isolates LM, BB, Lane 6: Band of size 702 bp obtained for isolate GP and (b) PCR product from amplification using W16-1/W16-2 primer pairs, Lane 1: 1 Kb ladder, Lanes 2-5: Band of size 702 bp obtained for isolates SP, NF, ML, PS |
In this study, a PCR product of 700 bp was obtained after amplification with primer pairs W16-1 and W16-2 (Fig. 3) and after digestion with Hae III, a single digestion product of 650 bp was obtained (Fig. 4) for all the seven P. infestans isolates tested. Estimation of the band sizes was performed using the software My Image Analysis (Thermo Fisher Scientific) in order to obtain precise band sizes. The results obtained here can be considered to indicate the presence of mating type A2 in the Mauritian isolates.
Random amplified polymorphic DNA (RAPD) analysis: Out of 40 primers screened, 30 primers (OPA 1-10, OPE 1-10 and OPL 1-10) generated reproducible amplification products and were used in the RAPD analysis. Each RAPD-PCR was performed in triplicate and a total of 140 reproducible fragments were obtained with primer set OPA, 133 with set OPE and 84 with set OPL, respectively, producing a total of 51 monomorphic and 300 polymorphic bands from the repeated amplifications. The size of the amplified products generated by each primer varied from 194-2957. Raw RAPD data was converted into binary data using the computer software NCSS v10 to generate similarity matrices based on simple matching coefficients and a dendogram was generated by hierarchical clustering. Distance values ranged from 1.0-1.6. The cophenetic correlation which resulted between the original distances and those that resulted from the cluster configuration was 0.92 and delta values were as follows: Delta(0.5): 0.049519 and Delta(1.0): 0.059688. Cluster analysis using the unweighted pair-group method with arithmetic averages (UPGMA) algorithm separated the isolates into 3 major groups (A, B and C). Group B formed the major group with 5 isolates (PS, ML, SP, NF and BB) while group A and C consisted each of one individual, GP and LM, respectively (Fig. 5). These results show that RAPD groups did not correlate to the geographical origins of the isolate as only isolates PS and ML belong were collected from geographically close regions.
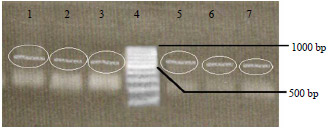 | |
| Fig. 4: | Restriction enzyme digestion using fast digest BsuR I (Hae III) of the PCR products amplified from P.infestans DNA with primer pair W16 1/ 2 : Lanes 1-3: Band sizes of 650 bp obtained with isolates LM, BB, GP, Lane 4: 100 bp ladder, Lane 5-7: Band sizes of 650 bp obtained with isolates SP, NF, ML |
A geographical clustering would have grouped together isolates GP, LM, ML and PS into one cluster, with isolates SP and NF as outliers, based on geographical locations.
DISCUSSION
In this study, mitochondrial DNA haplotypes were determined by amplification of the P1 (1,118 bp), P2 (1,070 bp), P3 (1,308 bp) and P4 (964 bp) regions of the mitochondrial genome, using oligonucleotides primers F1/R1, F2/R2, F3/R3 and F4/R4 and methods described by Griffith and Shaw15. PCR products were digested with enzymes CfoI (P1), MspI (P2) and EcoRI (P3 and P4). Digestion yielded restriction patterns by which the isolates could be classified into four haplotypes: Ia, Ib, IIa and IIb and Ic15. The main polymorphism in P. infestans is the presence (type II) or absence (type I) of a 2 kb insertion sequence. PCR products were successfully obtained for P1, P3 and P4, which all allowed discrimination between mitochondrial type I and type II. The results clearly indicate that all isolates tested belong to type II. Further distinction into IIa and IIb could not be completed due to the difficulty in obtaining PCR product P2. Gavino and Fry18 carried out a comprehensive survey of Phytophthora infestans mitochondrial diversity worldwide and report that only two mtDNA haplotypes, Ib and IIb were found to be tightly associated with particular clonal lineages, with haplotype Ib being very closely linked with the US-1 lineage or its variants (collected from the United States, Peru, Netherlands and the Philippines) and haplotype IIb closely linked with the US-6 lineage (collected primarily in the North Western part of North America).
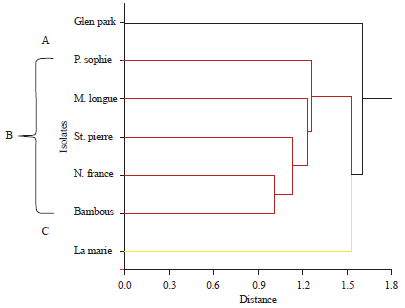 | |
| Fig. 5: | Dendogram generated by cluster analysis of different genotypes of P.infestans in Mauritius using random amplified polymorphic DNA (RAPD) loci |
| Table 1: | Multilocus genotypes of 7 Phytophthora infestans isolates from Mauritius after compilation of results from phenotypic and genotypic data |
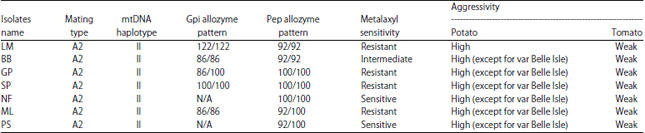 | |
The other mtDNA haplotypes were found in association with many different nuclear genomes across many countries, in both the A1 and A2 mating types, with no particular geographic or genetic backgrounds. In Mauritius, genetic characterization of P. infestans has been carried out on only one isolate collected in 2000 and it was found to be of the US-1 lineage. It is clear that, in the last 16 years, the island has become host to a much wider diversity of P. infestans strains, which have most probably displaced the old US-1 clonal lineage, as has been the case in many other parts of the world.
In order to determine mating type, this study made use of the CAPS assay previously described by Mazakova et al.17, instead of the conventional pairing method. The disadvantage of the pairing method is that it relies on the availability of tester isolates and oospore production may also be hampered by other factors such as aged cultures, various substrates, physical wounding, reduced metabolic activity of isolates on artificial media, exact incubation temperatures, light intensity and air flow over the petri dishes. The existence of a molecular method that directly targets polymorphism in the mating type locus is therefore more advantageous and provides a more robust and reliable way to ascertain mating type, without the absolute requirement for viable tester isolates and reproducing exact conditions for formation of oospores on artificial media. The W16-1/2 primers, originally designed by Judelson et al.16 are unique to the mating type locus of the P. infestans genome only and we can hence safely assume that the CAPS assay is a reliable way to ascertain mating type in P. infestans and provides a useful alternative to the conventional pairing method. The assay proved that all the seven isolates tested belonged to mating type A2. One Mauritian P. infestans isolate was characterized as belonging to the US-1genotype, which is characterized by the A1 mating type and it is not known for how long the A2 mating type has been present in Mauritius. However, it is clear that the A2 mating type is now widely distributed in almost all countries where potato and tomato is grown and in at least one other country, Bolivia, the P. infestans population has been reported to be clonal, A2 mating type and host-specific, which seems to be the case for the Mauritian isolates tested in this study as well. Since the A1 mating type had been previously discovered on the island, it is now important to continue monitoring the mating type status of local isolates to aid in the prediction of the extent of sexual recombination and thus the risk of long-lived oospores serving as primary inoculum sources within the island itself.
Although the reliability of the RAPD method continues to be debated, RAPD markers have shown to be successful for studying the diversity of P. infestans populations, as reported by Punja et al.19 for isolates in Canada, Zhu et al.20 and Li et al.21 for Chinese isolates, Mahuku et al.22 for Canadian isolates, Tosun et al.23 for Turkish isolates and Atheya et al.24 for Indian isolates. The RAPD profiles of the 7 tested P. infestans isolates used in this study revealed a total of 140 reproducible fragments with primer set OPA, 133 with set OPE and 84 with set OPL, respectively with 51 monomorphic and 300 polymorphic bands obtained from the repeated amplifications. Analysis of the results showed that no two isolates have the same RAPD profile. Hierarchical clustering using a phenetic approach categorized the isolates into 3 groups: A, B and C. The majority of isolates clustered together, irrespective of their geographical locations, forming group B (PS, ML, SP, NF and BB). Group A comprised of isolate GP only and group C contained isolate LM only.
The seven local P. infestans isolates tested in this study were previously characterized in terms of pathogenic fitness on potato and tomato hosts, metalaxyl sensitivity and isozyme analysis at the Gpi and Pep loci14 and now a complete multilocus phenotypic and genotypic profile can be drawn for each isolate (Table 1). When all this data is taken into consideration and codified to generate similarity matrices based on simple matching coefficients and generate a dendogram by hierarchical clustering, the same phenetic relationship is found as the one generated using RAPD data only. The seven isolates were found to cluster into 3 major groups (A, B and C). The major cluster (B) consisted of 5 isolates namely, BB, NF, SP, PS and ML. Cluster A and C consist each of one isolate, GP and LM, respectively. Cluster analysis showed that all the 7 isolates, although similar, possessed unique genetic constitutions. No evident correlation could be made between the isolates and the genetic markers. No correlation could also be found between geographical locations and genotypes present there. For example isolate GP, LM, ML and PS did not cluster together, although the four isolates were collected from the central region of the island. When the genotypic profile of each of the seven isolates in this study is compared to characterized isolates in other countries, it appears that isolate SP has exactly the same characteristics as US isolates US-11 and US-13, but the other Mauritian isolates resemble other reported isolates at only a few loci. Further research is warranted to differentiate between mitochondrial haplotypes IIa and IIb among Mauritian isolates before a definitive comparison can be made with other existing isolates. It is interesting to note that some relatively rare Gpi and Pep profiles have been revealed in these isolates (LM- Gpi: 122/122 and Pep: 92/92, ML- Gpi: 86/86 and Pep: 92/100), thereby possibly supporting the existence of previously uncharacterized lineages among the Mauritian P. infestans population.
CONCLUSION
This study reports, for the first time, the genotypic characteristics of Phytophthora infestans isolates currently causing blight outbreaks on the island of Mauritius in the Southern Indian Ocean region. We here report that all seven isolates collected over a period of three potato growing seasons all have unique genetic characteristics and are not genetically identical, as might have been previously thought for the small geographical area which the island covers. All isolates were found to be of mitochondrial type II, mating type A2 and RAPD fingerprinting data showed the isolates to be genetically distinct from each other. The information generated in this study will allow stakeholders involved in potato and tomato production and disease control to better understand the causal factors and epidemiology of the disease in order to predict the effectiveness and durability of disease management practices over the long term.
SIGNIFICANCE STATEMENTS
This study presents, for the first time, the genetic identity and characteristics of Phytophthora infestans isolates, the causal agent of potato and tomato late blight disease worldwide, from the Southern Indian Ocean region. Prior to this study, no information was available regarding the genotypic characteristics of P. infestans from this region of the world and neither the origin nor the migration patterns of this devastating pathogen could be inferred with reference to the Southern Indian Ocean region. This study reveals a very wide diversity among the existing populations of P. infestans found on Mauritius island and have important epidemiological consequences for the production of potato and tomato in Mauritius.
REFERENCES
- Cooke, D.E.L. and A.K. Lees, 2004. Markers, old and new, for examining Phytophthora infestans diversity. Plant Pathol., 53: 692-704.
CrossRefDirect Link - Jaime-Garcia, R., R. Trinidad-Correa, R. Felix-Gastelum, T.V. Orum, C.C. Wasmann and M.R. Nelson, 2000. Temporal and spatial patterns of genetic structure of Phytophthora infestans from tomato and potato in the Del Fuerte valley. Phytopathology, 90: 1188-1195.
CrossRefDirect Link - Jaimasit, P. and W. Prakob, 2010. Characterization of Phytophthora infestans population in potato crops from Chiang Mai and tak provinces. J. Agric. Technol., 1: 117-125.
Direct Link - Forbes, G.A., S.B. Goodwin, A. Drenth, P. Oyarzun, M.E. Ordonez and W.E. Fry, 1998. A global marker database for Phytophthora infestans. Plant Dis., 82: 811-818.
CrossRefDirect Link - Le, V.H., X.T. Ngo, M.B. Brurberg and A. Hermansen, 2008. Characterisation of Phytophthora infestans populations from Vietnam. Aust. Plant Pathol., 37: 592-599.
CrossRefDirect Link - Lehtinen, A., A. Hannukkala, T. Rantanen and L. Jauhiainen, 2007. Phenotypic and genetic variation in Finnish potato-late blight populations, 1997-2000. Plant Pathol., 56: 480-491.
CrossRefDirect Link - May, K.J. and J.B. Ristaino, 2004. Identity of the mtDNA haplotype(s) of Phytophthora infestans in historical specimens from the Irish potato famine. Mycol. Res., 108: 471-479.
CrossRefDirect Link - Goodwin, S.B., B.A. Cohen and W.E. Fry, 1994. Panglobal distribution of a single clonal lineage of the Irish potato famine fungus. Proc. Nat. Acad. Sci. USA., 91: 11591-11595.
PubMedDirect Link - Widmark, A.K., B. Andersson, A. Cassel‐Lundhagen, M. Sandstrom and J.E. Yuen, 2007. Phytophthora infestans in a single field in southwest Sweden early in spring: Symptoms, spatial distribution and genotypic variation. Plant Pathol., 56: 573-579.
CrossRefDirect Link - Wangsomboondee, T. and J.B. Ristaino, 2002. Optimization of sample size and DNA extraction methods to improve PCR detection of different propagules of Phytophthora infestans. Plant Dis., 86: 247-253.
CrossRefDirect Link - McLeod, A., S. Denman, A. Sadie and F.D.N. Denner, 2001. Characterization of South African isolates of Phytophthora infestans. Plant Dis., 85: 287-291.
CrossRefDirect Link - Mukalazi, J., E. Adipala, T. Sengooba, J.J. Hakiza, M. Olanya and H.M. Kidanemariam, 2001. Metalaxyl resistance, mating type and pathogenicity of Phytophthora infestans in Uganda. Crop Prot., 20: 379-388.
CrossRefDirect Link - Taleb-Hossenkhan, N. and A. Ibrahim, 2015. Pathogenic fitness, Metalaxyl sensitivity and Isozyme analysis of Phytophthora infestans isolates from Mauritius. Plant Pathol. J., 14: 113-122.
CrossRefDirect Link - Griffith, G.W. and D.S. Shaw, 1998. Polymorphisms in Phytophthora infestans: Four mitochondrial haplotypes are detected after PCR amplification of DNA from pure cultures or from host lesions. Applied Environ. Microbiol., 64: 4007-4014.
Direct Link - Judelson, H.S., L.J. Spielman and R.C. Shattock, 1995. Genetic mapping and non-Mendelian segregation of mating type loci in the oomycete, Phytophthora infestans. Genetics, 141: 503-512.
Direct Link - Mazakova, J., V. Taborsky, M. Zouhar, P. Rysanek, E. Hausvater and P. Dolezal, 2006. Occurrence and distribution of mating types A1 and A2 of Phytophthora infestans (Mont.) de Bary in the Czech Republic. Plant Prot. Sci., 42: 41-48.
Direct Link - Gavino, P.D. and W.E. Fry, 2002. Diversity in and evidence for selection on the mitochondrial genome of Phytophthora infestans. Mycologia, 94: 781-793.
CrossRefDirect Link - Punja, Z.K., H. Forster, I. Cunningham and M.D. Coffey, 1998. Genotypes of the late blight pathogen (Phytophthora infestans) in British Columbia and other regions of Canada during 1993-1997. Can. J. Plant Pathol., 20: 274-282.
CrossRefDirect Link - Li, B.J., X. Lv, Q.H. Chen, C.Z. Lan, J. Zhao, R.Z. Qiu and Q.Y. Weng, 2009. Population genetic diversity of Phytophthora infestans from China as revealed by SSRs and RAPDs. Chin. J. Agric. Biotechnol., 6: 161-170.
CrossRefDirect Link - Mahuku, G., R.D. Peters, H.W. Platt and F. Daayf, 2000. Random amplified polymorphic DNA (RAPD) analysis of Phytophthora infestans isolates collected in Canada during 1994 to 1996. Plant Pathol., 49: 252-260.
CrossRefDirect Link - Tosun, N., A. Yildrim, H. Turkusay and B. Tanyolac, 2007. Genetic variation among Phytophthora infestans (tomato blight) isolates from western Turkey revealed by Inter Simple Sequence Repeat (ISSR) and random amplified polymorphic DNA (RAPD) markers. Pak. J. Bot., 39: 897-902.
Direct Link - Atheya, I., B.P. Singh, S.K. Chakrabarti and D. Pattanayak, 2005. Genetic diversity and differentiation of Indian isolates of Phytophthora infestans as revealed by RAPD analysis. Indian J. Exp. Biol., 43: 817-823.
Direct Link