
Ann Afamefuna Jerry Mofunanya
Department of Botany, University of Calabar, PMB 1115, Calabar, Nigeria
Ayodeji Timothy Owolabi
Department of Botany, University of Calabar, PMB 1115, Calabar, Nigeria
LiveDNA: 234.12392
Ani Nkang
Department of Botany, University of Calabar, PMB 1115, Calabar, Nigeria
Plant Pathology Journal
Year: 2016 | Volume: 15 | Issue: 3 | Page No.: 86-94







ABSTRACT
Background: Telfairia occidentalis is pot-herb, cultivated extensively in Southern Nigeria. Methodology: Infection of the crop by Telfairia mosaic virus (TeMV) is characterized by severe mosaic and leaf malformation. Activities of polyphenol oxidase (PPO), peroxidase (POD), glucose-6-phosphahte dehydrogenase (G6PDH) and 6-phosphogluconate dehydrogenase (6PGDH) were evaluated spectrophotometrically using the appropriate wavelength for each of the enzymes, in two ecotypes of the vegetable infected with TeMV over a period of 120 Days After Inoculation (DAI). Results: From an apparent low level of activities at 14 and 30 DAI, there was a general increase in PPO activity in infected leaf samples of ecotype A compared to healthy controls. In ecotype B, PPO activity was highest at 14 DAI, followed by a decline and exhibited similar level of activity at later inoculation times. The POD activity was highest in ecotypes A and B at 60 and 90 DAI, respectively. For G6PDH, a rise in the activity in both ecotypes from 14-60 DAI was followed by a rather stable activity as disease progressed with percentage difference as higher as 107.3%. The activity of 6GPDH was highest at 30 DAI in both ecotypes. While the activity of the enzyme remained unchanged at later assay times in ecotype A, its activity was characterized by fluctuations in ecotype B. Values obtained for all the enzymes were significantly higher (p<0.05) in infected leaf tissues compared to their corresponding controls except for 6GPDH at later post inoculation times (60-120 DAI). Conclusion: Comparatively, the activities of all the enzymes in ecotype A, which has been found to be more susceptible to the TeMV were general higher than observed for ecotype B. It is thus suggested that the observed increases in the activity of these enzymes aided the disease severity engendered by TeMV, in the two ecotypes of T. occidentalis rather than disease mitigation as has been reported for some plant-virus combinations.
PDF Abstract XML References Citation
Received: April 02, 2016;
Accepted: May 16, 2016;
Published: June 15, 2016
How to cite this article
Ann Afamefuna Jerry Mofunanya, Ayodeji Timothy Owolabi and Ani Nkang, 2016. Time Course Evaluation of the Activities of Some Enzymes in Telfairia Mosaic Virus Infected Ecotypes of Telfairia occidentalis Hook f. Plant Pathology Journal, 15: 86-94.
DOI: 10.3923/ppj.2016.86.94
URL: https://scialert.net/abstract/?doi=ppj.2016.86.94
DOI: 10.3923/ppj.2016.86.94
URL: https://scialert.net/abstract/?doi=ppj.2016.86.94
INTRODUCTION
Telfairia occidentalis Hook. f., a member of the family Cucurbitaceae and commonly called fluted pumpkin is an important leafy vegetable in South Eastern Nigeria. Infection of the plant by Telfairia mosaic virus (TeMV), results in severe mosaic, shoestring and severe leaf malformation1,2. Induction of polyphenol oxidases and peroxidases has been implicated in disease resistance against plant pathogens, particularly in hypersensitive reactions characterized by programmed cell death, attributed to the production of Reactive Oxygen Species (ROS)3-6. Glucose-6-phosphate dehydrogenase (G6PDH EC 1.1.1.49) and 6-phosphogluconate dehydrogenase (6PGDH EC 1.1.1.44), on the other hand are two important enzymes in the Oxidative Pentose Phosphate (OPP) pathway for the generation of NADPH molecules which are important in many pathways, including the gluthatione-ascobate cycle essential in the defense against oxidative stress in plants7 and for the generation of adenosine triphosphate (ATP) molecules through the cytochrome system, which drive the biosynthetic processes needing energy8, including the polymerization of bases necessary for viral RNA synthesis. In plants according to Hauschild and Schaewen9, G6PDH isoenzymes are present in the cytosol and in plastids. The actions of the isoenzymes invariably lead to the formation of ribulose-5-phosphate. Ribose-5-phosphate, a derivative of ribulose-5-phosphate is believed to be raw materials for incorporation into viral genome10,11.
From the previous studies, the influence of virus infection on OPP pathway enzymes and some pathogenic-related enzymes and their activities in infected plants have been documented. While, a few cases of a decrease in their activities have been reported, majority of the reports pointed to increased activities of these enzymes in virus infected plants. Diaz-Vivancos et al.12 and Hernandez et al.13 reported that Plum Pox Virus (PPV) caused a decrease in some antioxidants enzymes but an increase in the class I ascorbate peroxidase (APX), peroxidase (designated as POX in the study) and polyphenol oxidase (PPO) enzymes in Prunus species. Hernandez et al.14 observed that in leaves of the very susceptible peach cultivar GF305, PPV infection brought about an increase in class I APX, POX, NADH-POX and PPO activities. They also reported that in susceptible apricot cultivar real fino, PPV produced a decrease in apoplastic POX and SOD activities, whereas a strong increase in PPO was observed. However, in the resistant apricot cultivar stark early orange, a rise in class I APX as well as a strong increase in POX and SOD activities was noticed in the apoplastic compartment. They also observed that long-term PPV infection produced an oxidative stress in the apoplastic space in apricot and peach plants as a result of the increase in hydrogen peroxide (H2O2) contents in this compartment. El-Fahaam et al.15 also reported considerable higher PPO and peroxidase (POD) activities in lettuce infected by Lettuce Mosaic Virus (LMV) 10 and 15 days after inoculation. Devanathan et al.16 have also observed high peroxidase activity in Banana Bunchy Top Virus (BBTV) infected cultivars of banana. Similarly, Meena et al.17 observed enhanced POD and PPO activities in Capsicum annuum L., infected with an unnamed geminivirus in diseased leaves as compared to healthy leaves. Arora et al.18 have also reported enhanced activities of POD and PPO in mothbean infected by Mothbean Yellow Mosaic Virus (MYMV). Comparison of superoxide dismutase (SOD), catalase (CAT), guaiacol peroxidase (GPX) and ascorbate peroxidase (APX) profiles of healthy and infected leaves of Impatients hawkerii and I. walleriana susceptible to Tomato Spotted Wilt Virus (TSWV)19 showed that long-term infection did not cause major changes in I. hawkerii but caused induction of several APX and GPX isoforms, increase in Mn-SOD and APX and a decrease in CAT activity in I. walleriana. Activities of enzymes involved in the detoxification of ROS (catalase, glutathione reductase, peroxidase and superoxide dismutase (SOD) were examined by Clarke et al.20 in the leaves of Phaseolus vulgaris L. var. "Top crop" treated with plant hormones and infected with a non-lesion-forming isolate of white clover mosaic virus (WClMV). The activities of catalase, glutathione reductase and SOD were reported to rapidly decline after infection while, peroxidase activity was enhanced. These changes were believed to occur before WClMV replication. Riedle-Bauer21 who studied oxidative stress in CMV-infected Cucumis sativus and zucchini yellow mosaic virus-infected Cucurbita pepo plants found virus-enhanced peroxidation via formation of ROS.
Sindelar and Sindelarova22 showed that the activity of 6PGDH (an enzyme that catalyzes the dehydrogenation of 6-phosphogluconate to ribulose-5-phosphate) was higher during the entire period in PVY-infected tissues of tobacco plants infected in a time course experiment compared to the values obtained for healthy control plants in both crude homogenate and partially purified enzyme preparations. Similarly, Sindelar et al.10 also demonstrated marked increase in the activities of G6PDH isoenzymes in potato infected by PVY. Tecsi et al.23 reported that infection of Cucumber pepo L. by Cucumber Mosaic Virus (CMV) produced a stimulatory effect on the capacity for OPP pathway, G6PDH and 6PGDH. Sindelar and Sindelarova11 reported that the activities of G6PDH were markedly increased in locally and systemically infected leaves of tobacco (Nicotiana tabacum L.) by Potato Virus Y (PVY), CMV, Potato Virus X (PVX), Tobacco Rattle Virus (TRV) and Turnip mosaic virus (TYMV) and the time courses of the activity were found to be linearly correlated with those of virus multiplication. They also observed that in leaves infected with PVY, the activity time courses of the crude and the partially purified G6PDH were found to be coincident. Sindelarova et al.24 found that there were slight increases in the activities of G6PDH and 6PGDH among other enzymes, in the thylakoid/envelope and stroma fractions of chloroplasts isolated from leaf tissues of susceptible tobacco plants infected with PVY. Infection with Tobacco Mosaic Virus (TMV) was also reported to produce increase in the activities of chloroplasts G6PDH and 6PGDH in both thylakoid/envelope and stroma fractions.
Previous studies to determine the effect of the TeMV on the chlorophyll content and photosynthetic capabilities of the two ecotypes of Telfairia occidentalis used in this study2 and on the growth characteristic of the ecotypes25 have been carried out. In this study, a time course evaluation of the activities of G6PDH, 6PGDH, polyphenol oxidase (PPO) and peroxidase (POD) in two ecotypes of T. occidentalis infected by TeMV, a potyvirus prevalent on the vegetable in South Western and Eastern Nigeria was carried out.
MATERIALS AND METHODS
Source of virus and propagation: The virus used in this study was the very isolate described by Shoyinka et al.1 generously supplied by Dr. H.J. Vetten of the Federal Biological Research Centre for Agriculture and Forestry (BBA) Braunschweig, Germany. The TeMV in leaf tissues stored under liquid nitrogen was reactivated by triturating the sample in cold disodium phosphate (Na2HPO4) buffer 0.03 M, pH 8.00 in pre-cooled sterilized pestle and mortar. The inoculum was applied by the conventional leaf-rub method (mechanical inoculation) with cotton swab onto Nicotiana benthamianum pre-dusted with 800 mesh carborundum. The inoculated leaves were rinsed with water and left for symptom development. Subsequent inoculations were carried out on T. occidentalis in which the virus was propagated in the screen-house at 23±3°C.
Seed sowing and inoculation with TeMV: Seeds of T. occidentalis obtained from Akparabong in Ikom local government area of cross river state (designated as ecotype A) and another seed lots obtained from Eket in Eket local government area of Akwa Ibom state, Nigeria (designated as ecotype B) were sown (1 seed/bag) in 16 cm diameter polyethylene bags. For each ecotype, there were 30 seedlings arranged in a randomized block design in 5 plots (six seedlings in each plot). The treatments were randomized within the plots and inoculation of the plants was mechanical. Inoculated plants were rinsed and left for symptom development.
Enzyme extraction: Enzyme extraction was carried out as described by Nkang26. About 2 g fresh leaf tissues from healthy and the TeMV infected ecotypes of T. occidentalis obtained randomly from the experimental plots (at the chosen assay days) were homogenized in 10 mL extraction buffer in a sterilized pestle and mortar. The extraction buffer consisted of 100 mM mixed monobasic potassium phosphate salt (KH2PO4) and dibasic potassium phosphate (K2HPO4), to which 4 g phenolic adsorbent polyvinyl polypyrrolidone (PVPP) was added. The buffer was then stirred adequately using a magnetic stirrer and adjusted to a final pH of 7.2. All extractions were carried out at 4°C. The homogenate was filtered through cheesecloth and the filtrate centrifuged at 5000 rpm for 5 min. The supernatant was stored on ice block in boxes and used as crude enzyme source in assaying for POD and PPO.
Enzyme purification: Two milliliter of crude enzyme extract was added 2 mL of cold acetone. The mixture was allowed to stand for 5 min and then centrifuged at 5000 rpm. The precipitate was suspended in 1 mL of cold assay buffer (KH2PO4 and K2HPO4) as partially purified enzyme filtrate. To this was added 0.8 g of ammonium sulphate [(NH4)2SO4] and stirred for 10 min on a magnetic stirrer. This was followed by centrifugation at 5000 rpm for 5 min after the addition of 2 g of (NH4)2SO4. The precipitate was suspended in 0.5 mL of the assay buffer and used for POD and PPO assays.
Determination of the G6PDH EC 1.1.1.49 and 6PGDH EC 1.1.1.44 activities: All enzyme assays were carried out at 30°C spectrophotometrically following the reduction of NADP at 340 nm as described by Nkang and Chandler27. The assay mixture contained the following: 100 mL triethanolamine hydrochloride-NAOH (pH 7.5), 10 mL MgCl2, 10 mL NADP, 10 mL 6-phosphogluconate or glucose 6-phosphate and 5 mL of the crude enzyme preparation. Two milliliter of the assay mixture was added 0.1 mL of 0.83 mM 6-phosphogluconate or 0.1 mL of 0.83 mM of glucose 6-phosphogluconate 0.1 mL of 12.5 mM MgCl2, 0.1 mL of 0.625 mM NADP and 0.1 mL of crude enzyme preparation were added to start the reaction in a final volume of 2.4 mL and read spectrophotometrically at 430 nm for both infected and healthy leaf samples of both ecotypes. Readings were taken after 4 min. All G6PDH activities were corrected for the possible complicating effect of 6PGDH on the G6PDH assays using the method of Goslinc and Ross28.
Polyphenol oxidase (PPO) assay: About 2 mL of the assay buffer (50 mM mixed phosphate salts, pH 7.2) in a cuvette was added 0.1 mL of 10 mM dihydroxyphenylalanine (DOPA) and 0.1 mL the enzyme preparation. The reaction was started with the addition of 0.1 mL of 10 mM H2O2 in a final volume of 3.3 mL. The absorbance of the mixture was measured after 1 min at a wavelength29 of 470 nm. The polyphenol oxidase activity was calculated using an extinction coefficient of 1433 mM cm–1 for quinone product30.
Determination of peroxidase (POD EC 1.11.1.7) activity: Assay for peroxidase activity was carried out as described by Nkang and Chandler27. The reaction mixture consisted of 50 mM mixed buffer containing 0.7 g KH2PO4 and K2HPO4 pH 7.2. The assay mixture contained in a final volume of 3.3 mL, made up of 3 mL of assay buffer and 0.1 mL of 10 mM guaiacol and 0.1 mL of enzyme preparation. The reaction was initiated with the addition of 0.1 mL of 10 mM H2O2. Absorbance readings were taken spectrophotometrically after 1 min at 436 nm with the assay buffer as blank26. The peroxidase activity was calculated using extinction co-efficient of 6.39 mol–1 cm–1 for guaiacol dehydrogenation product31.
Data analysis: Values presented were activities of the enzymes in unit per fresh weight (μg–1 FW). Data obtained were analyzed using independent t-test. Results were also expressed as percentage differences and differences between means were determined at 5% probability level.
RESULTS
Effect of TeMV on PPO activity in ecotypes A and B: The results showed that from apparently low levels at 14 and 30 DAI when the values obtained were 2.89±0.0002 and 2.99±0.0001, respectively, there was an increase in the activity of PPO in TeMV infected leaf samples in ecotype A as the disease progressed. Irrespective of the stage of plant growth at the time of assay, values obtained for infected samples were significantly higher (p<0.05) than the corresponding healthy samples. Percentage differences were particularly high as the plant aged (72.8% at 90 DAI and 74.6% at120 DAI) (Table 1).
For ecotype B, the results showed that infected plants generally exhibited increased PPO activity compared to the healthy at all stages of plant growth. The highest activity was recorded at 14 DAI with a value of 2.96±0.0003. Subsequent determinations revealed a decline in the activity up till 60 DAI with value of 2.80±0.0002 and a relatively same level of activity of the enzyme in later stages of plant growth (Table 1). Statistically, the values obtained when compared to the corresponding healthy samples were significantly higher (p<0.05) with percentage differences ranging from 19.7- 26.1. Generally, activity of PPO in ecotype A was found to be higher compared to that recorded for ecotype B after 30 DAI.
Effect of TeMV on POD activity in ecotypes A and B: There was a rise in the activity of POD in ecotype A from 14 DAI with a value of 275.35±0.0006 up till 60 DAI when activity was highest with a mean value of 418.11±0.0005. This was followed by a slight decline in the enzyme activity as the plant aged. Percentage differences ranged from 40.0-76.7. All the values obtained for the infected samples were significantly higher (p<0.05) than the corresponding healthy samples (Table 2).
For ecotype B, the results (Table 2) showed that there was a progressive increase in the activity of POD in the ecotype from 14 DAI with a mean activity value of 251.33±0.0006 till 90 DAI when the highest activity of 371.57±0.0002 was obtained. Thereafter, there was a decline in the enzyme activity. Compared to the healthy samples, the enzyme activity was found to be significantly higher (p<0.05) in TeMV infected leaves irrespective of the age at time of assay.
| Table 1: | Time course activity of polyphenol oxidase (PPO) in Telfairia occidentalis inoculated with Telfairia mosaic virus (TeMV) |
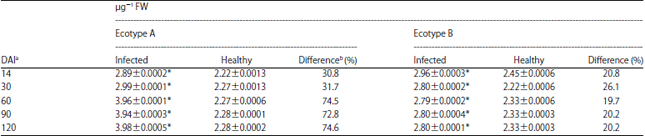 | |
| Values are means of 3 readings, *Significantly different (p<0.05), aDays after inoculation and bPercentage difference was obtained by expressing the difference between the value for the control and inoculated as a percentage of the control | |
| Table 2: | Time course activity of peroxidase (POD) activity in Telfairia occidentalis inoculated with Telfairia mosaic virus (TeMV) |
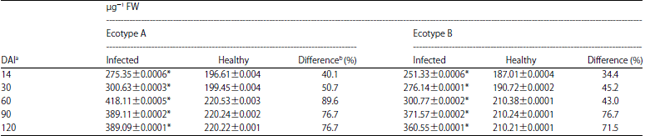 | |
| Values are means of 3 readings, *Significantly different (p<0.05), aDays after inoculation and bPercentage difference was obtained by expressing the difference between the value for the control and inoculated as a percentage of the control | |
| Table 3: | Time course activity of glucose-6-phosphate dehydrogenase (G6PDH) activity in two ecotypes of Telfairia occidentalis inoculated with Telfairia mosaic virus (TeMV) |
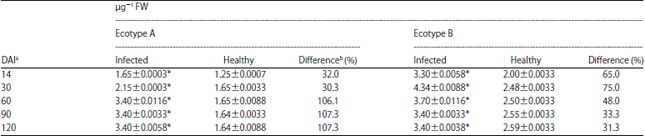 | |
| Values are means of 3 readings, *Significantly different (p<0.05), aDays after inoculation and bPercentage difference was obtained by expressing the difference between the value for the control and inoculated as a percentage of the control | |
Percentage differences ranged from 34.4-71.5. Similar to the activity of PPO, the results showed that the activity of POD was generally higher in ecotype A than in ecotype B.
Effect of TeMV on G6PDH in ecotype A and B: The results showed that there was an increase in the activity of the enzyme in ecotype A from a mean value of 1.65±0.0003 obtained at 14 DAI to 3.40±0.0116 recorded at 60 DAI (Table 3). Thereafter, there was no increase or decrease in the activity of the enzyme with prolonged infection. The values obtained for infected leaf samples were significantly higher (p<0.05) than their corresponding healthy controls at all post-inoculation times. Percentage differences were between 30.3 at 14 DAI and 107.3 at 90 DAI. For ecotype B, there was a general increase in the activity of G6PDH in infected leaf samples compared to the healthy control, irrespective of the age of plant at time of assay (Table 3). The highest peak of activity was recorded at 30 DAI with an activity value of 4.34±0.0088 followed by a decline to 3.70±0.0016 at 60 DAI and a lowered but sustained higher level of activity at 90 and 120 DAI. All the values obtained for the infected samples were significantly higher (p<0.05) compared to the corresponding values for the controls. Percentage increases compared to healthy controls ranged from 31.3% at 120 DAI to 75.0% at 30 DAI. The results also showed that the activity of the enzyme was generally higher in ecotype A than observed for ecotype B.
Effect of TeMV on 6GPDH activities in ecotypes A and B: The results of TeMV on 6GPDH are presented in Table 4. For ecotype A, the infected plants exhibited a general increase in 6GPDH activity at all stages of plant growth compared to healthy samples. The highest activity was observed 30 DAI with a value of 2.01±0.0013. For later inoculation stages, however, the activity of the enzyme remained unchanged and percentage differences ranged from 10.1-32.3%. The results also showed that the activities of the enzymes in infected plants were only significantly higher (p<0.05) than in healthy when assays were carried out between 14-30 DAI. For ecotype B, from an initial mean activity 3.32±0.001 at 14 DAI, there was an increase in the enzyme activity to a mean value of 4.02±0.003 30 DAI. As infection became prolonged there was a progressive decrease in the enzyme activity. The values obtained for infected samples were however, significantly higher than the corresponding values for the controls at all assay dates. Percentage differences ranged from 22.9 at 120 DAI to 67.7 at 14 DAI. The results also showed that there was higher 6GPDH activity in ecotype B than in A.
| Table 4: | Time course activity of 6-phosphogluconate dehydrogenase (6GPDH) activity in two ecotypes of Telfairia occidentalis inoculated with Telfairia mosaic virus (TeMV) |
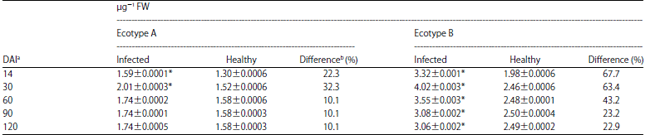 | |
| Values are means of 3 readings, *Significantly different (p<0.05), aDays after inoculation and bPercentage difference was obtained by expressing the difference between the value for the control and inoculated as a percentage of the control | |
DISCUSSION
Time course evaluation of the activities of PPO, POD, G6PDH and 6GPDH enzymes in two ecotypes of T. occidentalis were studied over a period of 120 days after inoculation with TeMV. The enzyme activities were measured spectrophotometrically, using appropriate wavelength for each of the enzymes assayed.
The result of this study showed that inoculation of both ecotypes engendered significant increase in PPO activity in TeMV-infected ecotypes of T. occidentalis compared to the healthy controls at all post-inoculation times. Similar observations have been reported in several host plant-virus combinations including lettuce infected with LMV15, banana infected by BBTV16, Prunus species infected with PPV12, P. vulgaris var. top crop infected with WCLMV and tomato infected with Tomato Yellow Leaf Curl Virus (TYLCV)32.
The results also showed that there was significant increase in POD activity in TeMV infected T. occidentalis in both ecotypes compared to the healthy controls at all the different post-inoculation times. Increased peroxidase activity in response to viral infection has similarly been reported in several host-virus combinations including soybean infected by Soybean mosaic virus (SoyMV)33, Cucumber Mosaic Virus (CMV) and Zucchini Yellow Mosaic Virus (ZYMV) infected Cucumis sativus and Cucurbita pepo plants, respectively21 and peaches and apricots infected with PPV13,34; tomato and bell pepper infected with Tobacco Mosaic Virus (TMV) and tomato mosaic tobamovirus35, Beans Yellow Mosaic Virus (BYMV) infection of beans36, Tomato Yellow Leaf Curl Virus (TYLCV) infected tomato plants37, tobacco infected with pepper mild mottle virus (PMMoV)38 and cotton inoculated with cotton leaf curl burewala virus (CLCuBuV)39. The results obtained in the study however, contrast with that of Hernandez et al.13 who reported a drop in the activity of POD in real fino, a susceptible apricot cultivar to PPV and also the report of Garcia-Ibarra et al.40 who observed a decrease of about 34% in the activity of POD in apricot infected with Apple Chlorotic Leaf Spot Virus (ACLSV).
Involvement of antioxidant enzymes (PPO and POD) in disease modulation in virus infected plants has been suggested. Hernandez et al.13 reported that the enhanced antioxidant enzyme activity induced by PPV in apricot cultivar Stark Early Orange (cv. SEO, a resistant varieties), among other factors was responsible for its resistance to the virus, while reduced activities of antioxidant enzymes observed in susceptible cultivar real fino was also said to be responsible for chlorosis symptom observed. Riedle-Bauer21 had earlier inferred that induction of peroxidase isoforms was responsible for growth reduction and malformation in C. sativus and Cucurbita pepo infected by CMV and ZYMV, respectively. Ashfaq et al.41 opined that differential reaction of Mash-80 (susceptible) and CM-2002 (resistant) varieties of blackgram (Vigna mungo) to UCLV was as result of differences in the level of peroxidase production and therefore, submitted that peroxidases might be involved in resistance/susceptibility to UCLV. Garcia-Ibarra et al.40 reported that enhanced activity of antioxidant enzymes protected the apricot variety "GF305" against Apple Chlorotic Leaf Spot Virus (ACLSV) (a variety that was highly susceptible to the virus) after a long post infection time. On the other hand, the induction of irreversible symptom development in banana infected by BBTV has been attributed to significant changes in enzyme levels of PPO, POX, among others antioxidant enzymes42.
Polyphenol oxidases (PPOs) are generally believed to be important in the initial stage of plant defense following damage to membrane causing the release of phenols and their subsequent oxidation to quinines43 which are lethal to pathogens44. Enhanced POD activity has been implicated as one of the first enzymes responding and providing fast defense against plant pathogens45,46, a function attributed to their involvement in lignification, suberization and polymerization of hydroxyl-proline-rich glycoproteins, regulation of cell wall elongation and wound healing47,48.
The result of this study suggests that rather than disease mitigation, the enhanced production of PPO and POD in T. occidentalis TeMV combination could have contributed to the disease severity (apparently arising from prevention of oxidation of phenolics into quinones by PPO and detoxification of peroxides by POD) as symptoms (mosaic and leaf malformation) elicited in the host plant became more and more severe consequent upon prolonged infection.
The results of this study also revealed that activities of G6PDH and 6GPDH were significantly increased in both ecotypes compared to healthy controls at all post-inoculation times. This result is consistent with earlier observation of increased activities of G6PDH and 6GPDH in different plant-virus combinations10,11,23,24. The involvement of both G6PDH and 6PGDH, particularly the former, in plant defense mechanism has been suggested49. The G6PDH is believed to be a key enzyme for the maintenance of redox potential in cell50 and leads the NADPH production through OPP pathway which is important in gluthatione reduction. Reduced gluthatione, according to Singh et al.51, protects the cell from oxidative damage by destroying hydrogen peroxide (H2O2) and peroxidase free radicals. Both G6PDH and 6PGDH are important enzymes in OPP pathway through which NADPH molecules are generated.
Besides enhancing susceptibility of T. occidentalis to TeMV as could be inferred in this study, increased activities of G6PDH and 6PGDH could also be attributed to attempt by the activities of some enzymes in TeMV infected ecotypes of Telfairia occidentalis infected plant to meet the increasing demand for energy (ATP) for viral genome replication and virus coat protein synthesis and to compensate for the expected turn-down of processes such as photosynthesis, glycolysis, increased respiration and lowered sugar level often associated with virus infection of plants2,25.
CONCLUSION
| • | Generally, there were significant increases in the activities of the PPO, POD, G6PDH and 6PGDH enzymes compared to their corresponding healthy controls at all post inoculation times |
| • | Activities of these enzymes were also found to be higher in ecotype A than ecotype B irrespective of the time of assay |
| • | Overall, the results of this study suggest that increased activities of these enzymes could have been responsible, among other factors for the disease severity associated with TeMV infection of the vegetable rather than disease mitigation as have been reported for some plant-virus combinations |
| • | Moreover, increased activities of all the enzymes in ecotype A did not confer any defense advantage on the ecotype relative to ecotype B with less PPO, POD, G6PDH and 6PGDH activities |
ACKNOWLEDGMENT
The authors are grateful to the Department of Botany of the University of Calabar, Calabar, Nigeria for providing the facilities to carry out the study.
REFERENCES
- Shoyinka, S.A., A.A. Brunt, D. Lesemann, G. Thottappilly and R.J. Lastra, 1987. The occurrence, properties and affinities of Telfairia mosaic virus, a potyvirus prevalent in Telfairia occidentalis (Cucurbitaceae) in South Western Nigeria. J. Phytopathhol., 119: 13-17.
Direct Link - Mofunanya, A.A.J., A.T. Owolabi and A. Nkang, 2014. Effects of Telfairia mosaic virus (TeMV) on the chlorophyll content and photosynthetic capabilities of two ecotypes of Telfairia occidentalis Hook. f. (Fluted Pumpkin). Int. J. Plant Pathol., 5: 54-62.
CrossRefDirect Link - Lamb, L. and R.A. Dixon, 1997. The oxidative burst in plant disease resistance. Annu. Rev. Plant Physiol. Plant Mol. Biol., 48: 251-275.
CrossRefPubMedDirect Link - De Gara, L., M.C. de Pinto and F. Tommasi, 2003. The antioxidant systems vis-a-vis reactive oxygen species during plant-pathogen interaction. Plant Physiol. Biochem., 41: 863-870.
CrossRefDirect Link - Mohase, L., A.J. de Westhuizen and Z.A. Pretorius, 2011. Involvement of reactive oxygen species generating enzymes and hydrogen peroxide in the rust resistance response of sunflower (Helianthus annuus L.). South Afr. J. Plant Soil, 28: 64-68.
CrossRefDirect Link - Ngadze, E., D. Icishahayo, T.A. Coutinho and J.E. van der Waals, 2012. Role of polyphenol oxidase, peroxidase, phenylalanine ammonia lyase, chlorogenic acid and total soluble phenols in resistance of potatoes to soft rot. Plant Dis., 96: 186-192.
CrossRefDirect Link - Potters, G., L.D. Gara, H. Asada and N. Horemans, 2002. Ascorbate and glutathione: Guardians of the cell cycle, partners in crime? Plant Physiol. Biochem., 40: 537-548.
CrossRefDirect Link - Hauschild, R. and A. von Schaewen, 2003. Differential regulation of glucose-6-phosphate dehydrogenase isoenzyme activities in potato. Plant Physiol., 133: 47-62.
CrossRefDirect Link - Sindelar, L., M. Sindelarova, N. Cerovska and M. Hanusova, 1990. Changes in ribonuclease and glucose-6-phosphate dehydrogenase activities during PVY-RNA biosynthesis in potato leaf discs. Biologia Plantarum, 32: 119-127.
CrossRefDirect Link - Sindelar, L. and M. Sindelarova, 2002. Glucose-6-phosphate dehydrogenase/6-phosphogluconate dehydrogenase ratio and the glucose-6-phosphate, 6-phosphogluconate and fructose-6-phosphate contents in tobacco plants infected with potato virus Y. Biol. Plantarum, 45: 575-580.
CrossRefDirect Link - Diaz-Vivancos, P., M. Rubio, V. Mesonero, P.M. Periago, A.R. Barcelo, P. Martinez-Gomez and J.A. Hernandez, 2006. The apoplastic antioxidant system in Prunus: Response to long-term plum pox virus infection. J. Exp. Bot., 57: 3813-3824.
CrossRefDirect Link - Hernandez, J.A., P. Diaz-Vivancos, M. Rubio, E. Olmos, A. Ros-Barcelo and P. Martinez-Gomez, 2006. Long-term plum pox virus infection produces an oxidative stress in a susceptible apricot, Prunus armeniaca, cultivar but not in a resistant cultivar. Physiologia Plantarum, 126: 140-152.
CrossRefDirect Link - Hernandez, J., P. Diaz-Vivancos, M. Rubio, E. Olmos, M. Clemente, A. Ros-Barcelo and P. Martinez-Gomez, 2007. Plum pox virus (PPV) infection produces an imbalance on the antioxidative systems in Prunus species. Acta Phytopathologica Entomologica Hungarica, 42: 209-221.
CrossRefDirect Link - El-Fahaam, Y.M., G.I. Fegla, E.E. Wagih and H.A. El-Karyoni, 1990. Biochemical changes in lettuce plants infected with lettuce mosaic virus. J. King Saud Univ. Agric. Sci., 2: 271-277.
Direct Link - Devanathan, M., M. Ramaiah, A.R. Sundar and M. Murugan, 2005. Changes of peroxidase and polyphenol oxidase in bunchy top nana virus infected and healthy cultivars of banana. Ann. Plant Physiol., 19: 114-116.
Direct Link - Arora, R., U. Joshi, P. Gupta and J. Singh, 2009. Effect of Yellow mosaic virus on pathogenesis related enzymes and chlorophyll content in mothbean (Vigna aconitifolia). Acta Phytopathologica Entomologica Hungarica, 44: 49-60.
CrossRefDirect Link - Milosevic, S., A. Simonovic, A. Cingel, S. Jevremovic, S. Todorovic, B. Filipovic and A. Subotic, 2012. Response of antioxidative enzymes to long-term Tomato spotted wilt virus infection and virus elimination by meristem-tip culture in two Impatiens species. Physiol. Mol. Plant Pathol., 79: 79-88.
CrossRefDirect Link - Clarke, S.F., P.L. Guy, D.J. Buritt and P.E. Jameson, 2002. Changes in the activities of antioxidant enzymes in response to virus infection and hormone treatment. Physiologia Plantarum, 114: 157-164.
CrossRefPubMedDirect Link - Riedle-Bauer, M., 2000. Role of reactive oxygen species and antioxidant enzymes in systemic virus infections of plants. J. Phytopathol., 148: 297-302.
CrossRefDirect Link - Sindelar, L. and M. Sindelarova, 1987. Changes in the activity of phosphogluconate dehydrogenase and its regulation in tobacco infected with PVY. Biol. Plantarum, 29: 150-154.
CrossRefDirect Link - Tecsi, L., A.M. Smith, A.J. Maule and C.R. Leegood, 1996. A spatial analysis of physiological changes associated with infection of cotyledons of marrow plants with cucumber mosaic virus. Plant Physiol., 111: 975-985.
PubMedDirect Link - Sindelarova, M., L. Sindelar, N. Wilhelmova and D. Prochazkova, 2005. Changes in key enzymes of viral-RNA biosynthesis in chloroplasts from PVY and TMV infected tobacco plants. Biologia Plantarum, 49: 471-474.
CrossRefDirect Link - Mofunanya, A.A.J., A.T. Owolabi and A. Nkang, 2015. Comparative study of the effect of telfairia mosaic virus (TEMV) on the growth characteristics of two ecotypes of Telfairia occidentalis (hooker fil). Int. J. Virol., 11: 54-65.
CrossRefDirect Link - Nkang, A. and G. Chandler, 1989. Changes during germination in rainforest seeds with orthodox and recalcitrant viability characteristics. J. Plant Physiol., 134: 9-16.
CrossRefDirect Link - Goslinc, P.G. and J.D. Ross, 1979. Characterization of glucose-6-phosphate dehydrogenase and 6-phosphogluconic acid dehydrogenase from hazel cotyledons. Phytochemistry, 18: 1441-1445.
CrossRefDirect Link - Kahn, V., 1983. Multiple effects of hydrogen peroxide on the activity of avocado polyphenol oxidase. Phytochemistry, 22: 2155-2159.
CrossRefDirect Link - Jimenez, M. and F. Garcia-Carmona, 1995. pH-induced hysteresis of latent broad bean polyphenol oxidase. Phytochemistry, 40: 373-376.
CrossRefDirect Link - Wang, C. and Y. Fan, 2014. Eugenol enhances the resistance of tomato against tomato yellow leaf curl virus. J. Sci. Food Agric., 94: 677-682.
CrossRefDirect Link - Zhuang, B.C., B. Xu and L. Liao, 1993. Change of superoxide dismutase, peroxidase and storage protein in soyabean leaves after inoculation with soyabean mosaic virus. Acta Phytopathologica Sinica, 23: 261-265.
Direct Link - Hernandez, J.A., J.M. Talavera, P. Martinez-Gomez, F. Dicenta and F. Sevilla, 2001. Response of antioxidative enzymes to plum pox virus in two apricot cultivars. Physiologia Plantarum, 111: 313-321.
CrossRefDirect Link - Madhusudhan, K.N., B.M. Srikanta, M.D. Shylaja, H.S. Prakash and H.S. Shetty, 2009. Changes in Antioxidant Enzymes, Hydrogen Peroxide, Salicylic Acid and Oxidative Stress in Compatible and Incompatible Host-Tobamovirus Interaction. J. Plant Interact., 4: 157-166.
CrossRefDirect Link - Radwan, D.E.M., K.A. Fayez, S.Y. Mahmoud and G. Lu, 2010. Modifications of antioxidant activity and protein composition of bean leaf due to Bean yellow mosaic virus infection and salicylic acid treatments. Acta Physiologiae Plantarum, 32: 891-904.
CrossRefDirect Link - Dieng, H., T. Satho, A. Abu Hassan, A.T. Aziz and R.E. Morales et al., 2011. Peroxidase activity after viral infection and whitefly infestation in juvenile and mature leaves of Solanum lycopersicum. J. Phytopathol., 159: 707-712.
CrossRefDirect Link - Hakmaoui, A., M.L. Perez-Bueno, B. Garcia-Fontana, D. Camejo, A. Jimenez, F. Sevilla and M. Baron, 2012. Analysis of the antioxidant response of Nicotiana benthamiana to infection with two strains of Pepper mild mottle virus. J. Exp. Bot., 63: 5487-5496.
CrossRefDirect Link - Siddique, Z., K.P. Akhtar, A. Hameed, N. Sarwar, Imran-Ul-Haq and S.A. Khan, 2014. Biochemical alterations in leaves of resistant and susceptible cotton genotypes infected systemically by cotton leaf curl Burewala virus. J. Plant Interact., 9: 702-711.
CrossRefDirect Link - Garcia-Ibarra, A., M.J. Clemente-Moreno, G. Barba-Espin, P. Diaz-Vivancos and M. Rubio et al., 2011. Changes in the antioxidative metabolism induced by Apple chlorotic leaf spot virus infection in peach [Prunus persica (L.) Batsch]. Environ. Exp. Bot., 70: 277-282.
CrossRefDirect Link - Ashfaq, M., M.A. Khan, N. Javed, S.M. Mughal, M. Shahid and S.T. Sahi, 2010. Effect of urdbean leaf crinkle virus infection on total soluble protein and antioxidant enzymes in blackgram plants. Pak. J. Bot., 42: 447-454.
Direct Link - Anuradha, C., R. Selvarajan, S. Vasantha and G.S. Suresha, 2015. Biochemical characterization of compatible plant virus interaction: A case study with bunchy top virus-banana host-pathosystem. Plant Pathol. J., 14: 212-222.
CrossRefDirect Link - Li, L. and J.C. Steffens, 2002. Overexpression of polyphenol oxidase in transgenic tomato plants results in enhanced bacterial disease resistance. Planta, 215: 239-247.
CrossRefDirect Link - Mohamed, H., A.A. El-Hady, M. Mansour and A.E.R. El-Samawaty, 2012. Association of oxidative stress components with resistance to flax powdery mildew. Trop. Plant Pathol., 37: 386-392.
CrossRefDirect Link - Sulman, M., G. Fox, A. Osman, A. Inkerman, P. Williams and M. Michalowitz, 2001. Relationship between total peroxidase activity and susceptibility to black point in mature grain of some barley cultivars. Proceedings of the 10th Australian Barley Technical Symposium, September 16-20, 2001, Canberra, ACT, Australia.
- Mohammadi, M. and H. Kazemi, 2002. Changes in peroxidase and polyphenol oxidase activities in susceptible and resistant wheat heads inoculated with Fusarium graminearum and induced resistance. Plant Sci., 162: 491-498.
CrossRefDirect Link - Yoshida, K., P. Kaothien, T. Matsui, A. Kawaoka and A. Shinmyo, 2003. Molecular biology and application of plant peroxidase genes. Applied Microbiol. Biotechnol., 60: 665-670.
CrossRefDirect Link - Maksimov, I., N. Troshina, O. Surina and E. Cherepanova, 2014. Salicylic acid increases the defense reaction against bunt and smut pathogens in wheat calli. J. Plant Interact., 9: 306-314.
CrossRefDirect Link - Scharte, J., H. Schon and E. Weis, 2005. Photosynthesis and carbohydrate metabolism in tobacco leaves during an incompatible interaction with Phytophthora nicotianae. Plant Cell Environ., 28: 1421-1435.
CrossRefDirect Link - Farhud, D.D. and L. Yazdanpanah, 2008. Glucose-6-phosphate dehydrogenase (G6PD) deficiency. Iran. J. Public Health, 37: 1-18.
Direct Link - Singh, S., A. Anand and P.K. Srivastava, 2012. Regulation and properties of glucose-6-phosphate dehydrogenase: A review. Int. J. Plant Physiol. Biochem., 4: 1-19.
Direct Link