
Rita Noveriza
Indonesian Agency for Agricultural Research and Development (IAARD), Indonesian Spices and Medicinal Crops Research Institute, Jl. Tentara Pelajar No. 3, Bogor 16111, Indonesia
LiveDNA: 62.11226
Gede Suastika
Faculty of Agriculture, Bogor Agricultural University, Jl. Kamper-Darmaga Campus, Bogor 16680, Indonesia
Sri Hendrastuti Hidayat
Faculty of Agriculture, Bogor Agricultural University, Jl. Kamper-Darmaga Campus, Bogor 16680, Indonesia
LiveDNA: 62.16401
Keiko T. Natsuaki
Laboratory of Tropical Plant Protection, Department of International Agricultural Development, Tokyo University of Agriculture, 1-1-1 Sakuragaoka, Setagaya-ku, Tokyo 156-8502, Japan
Plant Pathology Journal
Year: 2016 | Volume: 15 | Issue: 2 | Page No.: 65-74







ABSTRACT
Patchouli belonging to the family Lamiaceae is one of the important aromatic plants that originated from the Philippines. Telosma mosaic virus (TeMV) is one of the dominant types of viruses which attack Indonesian patchouli causing mosaic symptoms on the leaves. Only few research publications were conducted in Indonesia, hence this study was conceptualized in the information on the genome structure of the virus was not enough. The nucleotides sequence encoding the Coat Protein (CP) of TeMV-I (Indonesia isolate) was cloned and determined to complete information about this virus and designed an original primers for sequencing of the genes of NIb and NIa of TeMV-I. A part of genome structure of TeMV was sequenced from patchouli plants showing mosaic symptoms by using universal primer from the genus Potyvirus, then cloned to pGEM-T vector. Sequence analysis was done with program Sequence Scanner and CLC Sequence Viewer. The result showed that newly primer set for NIa and NIb region was design and cloned to TeMV-I. It was determined a sequence of 3805 nucleotides (nt) or a polypeptide chain of 1193 amino acid (aa) at the 3’-terminal region of the genome of TeMV. This contained the 3’-terminal part of the NIa (nts 1-1182), the NIb (nts 1183-2733), the coat protein (nts 2734-3549) and the 3’-UTR (nts 3550-3805). The stop codon (TAA) was followed by a 3’-untranslated region (3’-UTR) of 253 nt. The sequence analysis and development of specific primers for TeMV will be of great help in the detection and protection of this virus.
PDF Abstract XML References Citation
Received: December 07, 2015;
Accepted: February 08, 2016;
Published: March 15, 2016
How to cite this article
Rita Noveriza, Gede Suastika, Sri Hendrastuti Hidayat and Keiko T. Natsuaki, 2016. Molecular Cloning and Sequencing of Telosma Mosaic Virus
(TeMV) Causing Mosaic Disease on Patchouli. Plant Pathology Journal, 15: 65-74.
DOI: 10.3923/ppj.2016.65.74
URL: https://scialert.net/abstract/?doi=ppj.2016.65.74
DOI: 10.3923/ppj.2016.65.74
URL: https://scialert.net/abstract/?doi=ppj.2016.65.74
INTRODUCTION
Patchouli (Pogostemon cablin) belonging to the family Lamiaceae is one of the important aromatic and medicinal plants that originated from the Philippines (Sreedevi et al., 2009) but also reported a native from Indonesia (Hasegawa et al., 1992). The dry leaves of patchouli on steam distillation yield an essential oil called the patchouli oil (Sreedevi et al., 2009) which is used as a natural fixative in the aromatic industry (Hasegawa et al., 1992) and as main active ingredients of more than 30 common traditional Chinese medicines. Studies on the main active constituents of patchouli oil namely patchouli alcohol and pogostone, showed that they can be used as medicine (Zhang et al., 2006). They also possess anti-insecticidal, antifungal and bacteriostatic properties (Zhang et al., 2002) and were proved to inhibit neurotoxic activityof β-amyloid peptide (Huang et al., 2008) and to act as Reactive Oxygen Species (ROS) scavenger in oxidant-induced cell death of human neuroglioma cells (Kim et al., 2010).
The TeMV is one of the dominant types of viruses which attack Indonesian patchouli causing mosaic symptoms on the leaves of patchouli. This virus infection leads to decrease production of patchouli leaves and oil levels in the leaves. Nucleotides of coat protein this virus are submitted to Gene Bank with accession number AB699343.1, AB699342.1, AB699341.1, AB699340.1, AB699339.1, AB699338.1 and AB699131.1 (Noveriza et al., 2012a, b). As patchouli plants can only be propagated by stem cuttings, this method of propagation can accelerate the spread of the virus in patchouli production (Hartono and Subandiyah, 2006). Only few studies were conducted in Indonesia, hence this study was conceptualized in the information on the genome structure of the virus was not enough, the nucleotides sequence encoding the Coat Protein (CP) of TeMV-I (Indonesia isolate) was cloned and determined to complete information about this virus and designed an original primers for sequencing of the genes of NIb and NIa of TeMV-I.
MATERIALS AND METHODS
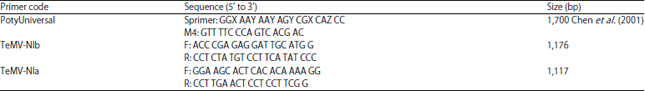
Virus isolates were obtained from patchouli plants collected in Indonesia and directly detected using a set of Poty Universal primers (Table 1). The NIa and NIb specific primers (Table 2) were designed based on the alignment of complete sequence of TeMV from Vietnam (ABI34612.1). Total plant RNAs were extracted from samples using Trizol® Reagent (Invitrogen, USA) as recommended by the manufacturer. First-strand cDNA were synthesized using ReverTra Ace-alpha-® kit (TOYOBO, Japan) according to the manufacturer’s instructions and the oligo d(T) primer M4T (5’-GTT TTC CCA GTC ACG AC (T)15-3’) as initial primer.
A degenerate primer (Sprimer: 5’-GGX AAY AAY AGY CGX CAZ CC-3’, x = A, G, C or T; Y = T or C; Z = A or G) was designed based on the amino acid sequence motif GNNSGQP according to Chen et al. (2001). Second-strand cDNAs of Coat Protein (CP) of TeMV were synthesized by polymerase chain reaction using TaKaRa Ex TaqTM (Takara Biomedicals, Japan) as recommended by the manufacturer. The PCR contained 5 μL template cDNA, 25 pmol of each amplification primer (M4: 5’-GTT TTC CCA GTC ACG AC-3’ and sprimer) (Table 2), 2 μL dNTP mixture, 2.5 μL 10xEx TaqTM Buffer and 0.1 μL TaKaRa Ex TaqTM in a total of 50 μL. Amplifications were for 30 cycles each of 0.5 min at 94°C, 1 min at 47°C and 2 min at 72°C with final extension 10 min at 72°C. The PCR products were examined by electrophoresis in 1.5% agarose gel. Gel extraction of PCR product was done using a Wizard@SV Gel and PCR clean up system following the manufacturer’s protocols.
The PCR fragments were cloned into the pGEM-T vector (Promega) following the manufacturer’s protocols and then isolated and purified by Labopass Plasmid Mini Kit (Hokkaido System Science, Japan).
| Table 1: | Sequences of primers used to amplify second-strand cDNA of telosma mosaic virus (TeMV) |
 | |
| Table 2: | DNA amplification of patchouli plant samples from Indonesia |
 | |
Additional primers were designed to internal sequences for sequencing. At least three independent PCR clones of each virus isolates were sequenced with the DNA analysis using Automate Sequencer (Applied Biosystems 3130 xl Genetic Analyzer, Japan). Sequence analysis was done with program Sequence Scanner version 1.0 and CLC Sequence Viewer version 6.6.2 (http://www.clcbio.com/ products/latest-improvements-sequence-viewer/).
RESULTS
The DNA fragments of the expected size of 3’terminal of RNA genomes of telosma mosaic virus (TeMV) were amplified from 3 tested Indonesian samples. They were amplified using primers TeMV-NIb and TeMV-NIa (Table 2).
A sequence of 3,805 nucleotides (nt) at the 3’-terminal region of the genome of Indonesian TeMV (TeMV-I) was determined (Fig. 1). This contained the 3’-terminal part of the NIa (nts 1-1182), the NIb (nts 1183-2733), the coat protein (nts 2734-3549) and the 3’-UTR (nts 3550-3805). Analysis showed that the sequence was very similar (85% identical nucleotides) to only one reported data of Telosma mosaic virus (ABI34612.1) (hereafter TeMV-V), genus Potyvirus from Telosma cordata in Vietnam.
The aa sequence homologies of the TeMV-I among TeMV-V and the previously reported species in the genus Potyvirus suggest that the relationship among them represent similar (91.1% identical to TeMV-V and 72.9-76.9% identical to bean common mosaic virus (BCMV), CABMV, PWP, SMV and watermelon mosaic virus (WMV). Here, are inserted 6 amino acids (AMAAGL) in sequence number 352.
DISCUSSION
Nucleotides of coat protein telosma mosaic virus (TeMV) are submitted to GenBank with accession number AB699343.1, AB699342.1, AB699341.1, AB699340.1, AB699339.1, AB699338.1 and AB699131.1 (Noveriza et al., 2012a, b) but that nucleotides of coat protein were not complete yet.
 | |
| Fig. 1: | Nucleotides sequence of the NIa (1182 nt), NIb (1550 nt), CP (815 nt) and 3’-UTR (253 nt) encoding gene of TeMV-I and deduced aa sequence. Thick underline and asterisk indicate 3’-UTR region and stop codon, respectively |
So need to find the full length coat protein of those sequences for complete information of that species in Indonesia. This study is conducted using cloning technique to find genome structure of TeMV from patchouli in Indonesia.
 | |
| Fig. 2: | Comparison of the aa sequence of TeMV-I poplyprotein (1183 aa; NIa, NIb and CP) with those of other potyvirus. Abbreviations for the viruses and sources of sequences are as described in the references 3082 aa, TeMV-aaV, Ha et al. (2008); 3169 aa, BCMNV-AK, Larsen et al. (2005); 3053 aa, CABMV-AZ, Mlotshwa et al. (2002); 3086 aa, PWV-AUST, Wylie and Jones (2011); 3067 aa, SMV-SKor, Seo et al. (2009) and 3217 aa, WMV-Fran, Desbiez and Lecoq (2004). Identical aa residues with TeMV-aaI are indicated by a black highlight and different aa, when compared to TeMV-aaI, are denoted. Hyphens are used to indicate deletions introduced for best alignment. Yellow and green highlight are used to indicate a conserve sites in CP and NIb Potyvirus genome, respectively |
Nucleotide sequence of 3’ terminal of RNA genome obtained and compared with sequences available in the GenBank indicated that this sequence belongs to the genus Potyvirus and species of telosma mosaic virus (TeMV), because of having 85% nucleotide identity over the whole genome of TeMV from Vietnam (Ha et al., 2008). This virus also reported in Thailand, it cause mosaic and fruit woodiness disease on passion fruit (Chiemsombat et al., 2014). According to King et al. (2011) that the current species demarcation criterion is either 85% nucleotide identity over the whole genome or 85% amino acid identity over the full length coat protein.
For further discussion, 3’ terminus of RNA genome of TeMVI (length 3,805 nts or 1,183 aa) was sequenced. These sequences consist of a part of NIa gene (394 aa), NIb gene (517 aa), CP gene (272 aa) and 3’ UTR (353 nts). According to Zheng et al. (2008), there were 17 conserved sites in genome of Potyvirus while four of them were identified by earlier worker. Of the previously identified sites, NIIENGV found by Nicolas and Laliberte (1991) ("ATNIIENG") ranked third, QMKAAA found by Langeveld et al. (1991) and Pappu et al. (1993) ranked 5th, GNNSGQ site identified by Gibbs and Mackenzie (1997) (PV2) in 1997 ranked 7th and the MVWCIE/DNG site identified by Langeveld et al. (1991) and Pappu et al. (1993) ("WCIEN") ranked 12th. The amino acid sequences of CP gene of TeMV-I and TeMV-V have 6 conserved sites, EN/DTERH, QMKAAA, YAFDFYE, MVWCIE/DNG, WV/TMMDGD/E/N and P/R/AYMPRYG (Fig. 2), which are generally owned by the Potyvirus group. On NIb gene of TeMV-I and TeMV-V genome, there were 6 conserved sites, CVDDFN, A/SMI/VES/AWG, GNNSGQ, GQPSTVVD, FTAAPL/ID/E and DGSQ/RFDA (Fig. 2).
When the amino acid sequence of the 3 terminal of RNA genome of TeMV was compared with 6 species in the genus Potyvirus, it was proved that there was a mutation in NIa gene (Nia-protease) of TeMV-I at QAMAAGLP (1115-1133 nt, Fig. 1). There were six additional amino acids at the end of NIa gene (AMAAGL, Fig. 2). The NIa-Pro is a trypsin-like cysteine proteinase responsible for the remaining of seven cleavage events in the C-terminal which was two thirds of the polyprotein (Guo et al., 2011). So NIa-protease gene (Nia-pro) plays an important role in multiplication of the virus. If the gene is isolated, it can be used in resistance to viral gene. Though TeMV is relatively a new reported virus, the occurrence of this virus in distinct countries like Vietnam, Indonesia and Thailand. In this regards, the sequence analysis and development of specific primers for TeMV in this study will be of great help in the detection and protection of this virus.
CONCLUSION
Telosma mosaic virus (TeMV) Indonesian isolate was clone and sequence using newly primer set for NIa and Nib region. It was determined a sequence of 3805 nucleotides (nt) or a polypeptide chain of 1193 amino acid (aa) at the 3’-terminal region of the genome of TeMV. This contained the 3’-terminal part of the NIa (nts 1-1182), the NIb (nts 1183-2733), the coat protein (nts 2734-3549) and the 3’-UTR (nts 3550-3805). The stop codon (TAA) was followed by a 3’-untranslated region (3’-UTR) of 253 nucleotides. The sequence analysis and development of specific primers for TeMV in this study will be of great help in the detection and protection of this virus.
ACKNOWLEDGMENTS
We wish to thank to Ms. Hiroko Otsu and Ms. Saki Yoshida for their help. This work was partially supported by the research fund from Laboratory of Tropical Plant Protection at Department of International Agricultural Development in Tokyo University of Agriculture, Japan and the scholarship from Indonesian Agency for Agricultural Research and Development (IAARD), Ministry of Agriculture, Indonesia.
REFERENCES
- Chen, J., J. Chen and M.J. Adams, 2001. A universal PCR primer to detect members of the Potyviridae and its use to examine the taxonomic status of several members of the family. Arch. Virol., 146: 757-766.
CrossRefDirect Link - Chiemsombat, P., S. Prammanee and N. Pipattanawong, 2014. Occurrence of Telosma mosaic virus causing passion fruit severe mosaic disease in Thailand and immunostrip test for rapid virus detection. Crop Prot., 63: 41-47.
CrossRefDirect Link - Desbiez, C. and H. Lecoq, 2004. The nucleotide sequence of Watermelon mosaic virus (WMV, Potyvirus) reveals interspecific recombination between two related potyviruses in the 5' part of the genome Arch. Virol., 149: 1619-1632.
CrossRefDirect Link - Gibbs, A. and A. Mackenzie, 1997. A primer pair for amplifying part of the genome of all potyvirids by RT-PCR. J. Virol. Methods, 63: 9-16.
PubMedDirect Link - Guo, B., J. Lin and K. Ye, 2011. Structure of the autocatalytic cysteine protease domain of potyvirus helper-component proteinase. J. Biol. Chem., 286: 21937-21943.
CrossRefDirect Link - Ha, C., S. Coombs, P.A. Revill, R.M. Harding, M. Vu and J.L. Dale, 2008. Design and application of two novel degenerate primer pairs for the detection and complete genomic characterization of potyviruses. Arch. Virol., 153: 25-36.
CrossRefPubMedDirect Link - Hasegawa, Y., K. Tajima, N. Toi and Y. Sugimura, 1992. An additional constituent occurring in the oil from a patchouli cultivar. Flavor Fragrance J., 7: 333-335.
CrossRefDirect Link - Huang, X.W., L. Bai, F.H. Xu and Y.J. Wu, 2008. Inhibitory activities of patchouli alcohol on neurotoxicity of β-amyloid peptide. Med. J. Chin. People Liberation Army, 24: 338-340.
Direct Link - Kim, H.W., S.J. Cho, B.Y. Kim, S.I. Cho and Y.K. Kim, 2010. Pogostemon cablin as ROS scavenger in oxidant-induced cell death of human neuroglioma cells. Evidence-Based Complement. Altern. Med., 7: 239-247.
CrossRefDirect Link - Langeveld, S.A., J.M. Dore, J. Memelink, A.F.L.M. Derks, C.I.M. van der Vlugt, C.J. Asjes and J.F. Bol, 1991. Identification of potyviruses using the polymerase chain reaction with degenerate primers. J. Gen. Virol., 72: 1531-1541.
CrossRefDirect Link - Larsen, R.C., P.N. Miklas, K.L. Druffel and S.D. Wyatt, 2005. NL-3 K strain is a stable and naturally occurring interspecific recombinant derived from Bean common mosaic necrosis virus and Bean common mosaic virus. Phytopathology, 95: 1037-1042.
CrossRefDirect Link - Mlotshwa, S., J. Verver, I. Sithole-Niang, T. van Kampen, A. van Kamme and J. Wellink, 2002. The genomic sequence of cowpea aphid-borne mosaic virus and its similarities with other potyviruses. Arch. Virol., 147: 1043-1052.
CrossRefPubMedDirect Link - Nicolas, O. and J.F. Laliberte, 1991. The use of PCR for cloning of large cDNA fragments of turnip mosaic potyvirus. J. Virol. Methods., 32: 57-66.
CrossRefDirect Link - Noveriza, R., G. Suastika, S.H. Hidayat and U. Kartosuwondo, 2012. Potyvirus associated with mosaic disease on patchouli (Pogostemon cablin (Blanco) Benth.) plants in Indonesia. J. Int. Soc. Southeast Asian Agric. Sci., 18: 131-146.
Direct Link - Pappu, S.S., R. Brand, H.R. Pappu, E.P. Rybicki, K.H. Gough, M.J. Frenkel and C.L. Niblett, 1993. A polymerase chain reaction method adapted for selective amplification and cloning of 3' sequences of potyviral genomes: Application to dasheen mosaic virus. J. Virol. Methods, 41: 9-20.
CrossRefDirect Link - Seo, J.K., H.G. Lee, H.S. Choi, S.H. Lee and K.H. Kim, 2009. Infectious in vivo transcripts from a full-length clone of Soybean mosaic virus strain G5H. Plant Pathol. J., 25: 54-61.
Direct Link - Sreedevi, S.C., Y.R. Hegde and S.K. Prashanthi, 2009. Survey for fungal diseases of patchouli in Karnataka. Biomed, 4: 109-112.
Direct Link - Wylie, S.J. and M.G.K. Jones, 2011. The complete genome sequence of a Passion fruit woodiness virus isolate from Australia determined using deep sequencing and its relationship to other potyviruses. Arch. Virol., 156: 479-482.
CrossRefDirect Link - Zhang, Y., J.C. Zhang, Y. Chen, M.S. Yang, H. Cao and P.G. Xiao, 2006. Current progresses in pharmacognosy, chemistry and pharmacology of Pogostemon cablin. Chin. Tradit., 37: 786-790.
Direct Link - Zheng, L., P.J. Wayper, A.J. Gibbs, M. Fourment, B.C. Rodoni and M.J. Gibbs, 2008. Accumulating variation at conserved sites in potyvirus genomes is driven by species discovery and affects degenerate primer design. PloS One, Vol. 3.
CrossRefDirect Link - Hartono, S. and S. Subandiyah, 2006. Pemurnian dan deteksi serologi Patchouli mottle virus pada tanaman nilam. J. Perlindungan Tanaman Indonesia, 12: 74-82.
Direct Link