
S.K. Biswas
Department of Plant Pathology, Chandra Shekhar Azad University of Agriculture and Technology, Kanpur 208002, Uttar Pradesh, India
LiveDNA: 91.16360
Mohd. Rajik
Department of Plant Pathology, Chandra Shekhar Azad University of Agriculture and Technology, Kanpur 208002, Uttar Pradesh, India
Muneeshwar Sharma
Shere-Kashmir University of Agricultural and Sciences, Jammu and Kashmir, India
Prem Naresh
Department of Plant Pathology, Chandra Shekhar Azad University of Agriculture and Technology, Kanpur 208002, Uttar Pradesh, India
Upesh Kumar
Department of Plant Pathology, Chandra Shekhar Azad University of Agriculture and Technology, Kanpur 208002, Uttar Pradesh, India
Kishan Lal
Department of Plant Pathology, Chandra Shekhar Azad University of Agriculture and Technology, Kanpur 208002, Uttar Pradesh, India
Ravindra Singh
Department of Plant Pathology, Chandra Shekhar Azad University of Agriculture and Technology, Kanpur 208002, Uttar Pradesh, India
Plant Pathology Journal
Year: 2016 | Volume: 15 | Issue: 2 | Page No.: 57-64







ABSTRACT
Disease response to spot blotch by different varieties of wheat namely K 307, K 9644, K 9162, K 9351, K 9465 and HD 2985 revealed that all the varieties give variable response on disease severity. Among the varieties, minimum disease severity was recorded in K 9351 with the value of 30.07% at 65 days of plant. Biochemical analysis of leaves of different varieties of wheat revealed that the variable amount of phenol and soluble protein content. The maximum with 31.50 and 28.60 mg g–1 of soluble protein and 3.10 and 2.85 mg g–1 of total phenol content was found in K 9351 at 20 and 65 days of plant, respectively. The correlation co-efficient between disease severity and soluble protein and total phenol content at 65 days of wheat varieties revealed that there was negative correlation showing (r) -0.974 and -0.953. The regression equation of soluble protein and disease severity showed that higher regression value, lower disease incidence. It is also cleared that increase amount of total phenol and protein content both are associated with decreases of disease severity and both the parameter are reduced with the increase with age of plant. The RAPD analysis of different wheat varieties revealed that variable number and size of fragments are found among the varieties. The highest numbers of fragments are found in variety K 9351 which is 13. The presence or absence of fragment and their variable size might be the responsible for morphological and biochemical variability along with resistance response to spot blotch.
PDF Abstract XML References Citation
Received: October 09, 2015;
Accepted: January 30, 2016;
Published: March 15, 2016
How to cite this article
S.K. Biswas, Mohd. Rajik, Muneeshwar Sharma, Prem Naresh, Upesh Kumar, Kishan Lal and Ravindra Singh, 2016. Variable Disease Response to Spot Blotch in Different Eat Varieties and it Assessment at Biochemical and Genetics Level. Plant Pathology Journal, 15: 57-64.
DOI: 10.3923/ppj.2016.57.64
URL: https://scialert.net/abstract/?doi=ppj.2016.57.64
DOI: 10.3923/ppj.2016.57.64
URL: https://scialert.net/abstract/?doi=ppj.2016.57.64
INTRODUCTION
Wheat (Triticum aestivum L.) is one of the most important and widely cultivated crops in the world which is mainly used for human consumption and support nearly 35% of the world populations. It is grown on 8307 thousand ha with total production of 21.708 million t and an average yield of 2.615 t ha–1 (Anonymous, 2002). Global demand for wheat is growing at approximately 2% per year, twice the current rate of grain in genetic yield potential (Skovmand and Reynolds, 2000). In India, 95% grown wheat is hexaploid and used for the preparation of bread and other bakery products (Chaurasia et al., 1999). It is nutritious, easy to store and transport and can be processed into various types of food. The demand for wheat is expected to grow faster than any other major agricultural crop. To meet the needs of the growing world population, the forecast demand for the year 2020 varies between 840 (Rosegrant et al., 1995) and 1050 million t (Kronstad, 1997). Due to land limitations, the enhancement of wheat production must come from higher absolute yields, which can only be met by the concerted action of scientists involved in diverse agricultural disciplines and in particular by increased efforts in plant breeding (Braun et al., 1998). More than 350 cultivars have been released during the last four decades but only 250 varieties are notified by government of India. Most of the varieties are available for cultivations under different agro-climatic conditions (Kumar et al., 2011). But resistant varieties are scan to spot blotch. Globally an estimated 25 million ha of wheat cultivated land is affected by spot blotch disease (Van Loon et al., 1998; Snyder and Hansen, 1940). The widely applied rice-wheat cropping system of south Asia provides favorable environment for the survival and multiplication of foliar blight pathogens because rice serves as a host for spot blotch fungi and rice stable plays the role as a substrate for the fungi after rice harvest (Kumawat et al., 2008; Shetty and Ahmad, 1980). But disease severity response to spot blotch varies from varieties to varieties (Ahmad and Hossian, 2005; Porter and Smith, 1982; Sinha et al., 1991; Nasser et al., 1990). The disease has a special significance in eastern Gangetic plains of South Asia that includes India, Nepal and Bangladesh (Joshi et al., 2007). It has emerged as a serious yield limiting problem in north western part of India also (Singh et al., 1993). The pathogen causes pre- and post-emergence damping off, seedling blight and root-rot but the major yield losses are due to foliar blight phase of D. sorokiniania. The pathogen is responsible for 3-20% reduction in yield under different agro-climatic conditions (Nema and Joshi, 1971). A multi-location trial conducted on yield losses indicated 2.72-36.24% loss in yield (Parashar et al., 1995). The survival of the pathogen is mainly in seeds, soil and crop residue. Several factors like heat stress, soil water deficient, light texture soil and host physiology have generally been reported to enhance the development of pathogen. Therefore, the study was undertaken in the present investigation.
MATERIALS AND METHODS
Collection of wheat genotypes: The popular varieties of wheat namely K-307, K-9644, K-9162, K-9351, K-9465 ands HD-2985 are commonly used by the farmers around the vicinity of Kanpur. The seeds of all varieties are collected from rabi cereals, Chandra Shekhar Azad University of Agriculture and Technology, Kanpur for conducting the present investigation.
Field trial: The field trial was conducted at Agriculture Research Farm of C.S. Azad University of Agriculture of Technology, Kanpur-208002 to evaluate variable disease response to spot blotch in different wheat varieties and it assessment at biochemical and genetics level. Recommended agronomical practices were followed. The experiment was laid out Randomizes Block Design (RBD) with three replications. The observations on disease severity were taken 65 days age of plant. The leaf samples were collected at 20 and 65 days of plant for estimation of total soluble protein, total phenol content in wheat leaves and also Random Amplified Polymorphic DNA (RAPD) analysis.
Measurement of disease severing: Disease observations were recorded at 65 days of sowing. The total disease leaf area in different varieties of wheat was calculated from 50 randomly selected leave of diseased plant. The sketch of each leaf was drawn on a graph paper (mm) and area covered by leaf was measured. The average of all the reading of 50 leaves was calculated to get leaf area in cm3. Similarly, disease area of total leaf area was calculated. Leaf with no sign of infection received a score of zero while those with highest received score of IV. Similarly, leave with 1-25, 26-50 and 51-75, area, covered with spot received a score of I, II, III, respectively. The disease severity of plants was calculated by the following formula (Chenulu and Singh, 1964).
Biochemical studies
Soluble protein estimation: The soluble protein contents were determined by Lowry et al. (1951) with slight modification. Wheat leaves from different varieties were harvested, washed with distilled water several times and blotter dried before protein extraction. A quantity of 1 g of each leaf sample was cut into small pieces and grinded in pestle and mortar as 1:5 ratio of leaves extraction solution. The suspension was centrifuged at 12000 rpm for 30 min at 4°C. The supernatant was collected and used for protein estimation. The working standard solution was pipette out 0.2, 0.4, 0.6 and 1.0 mL and was put into series of test tubes. A quantity of 0.2, 0.4, 0.6 and 1.0 mL of the sample extract was also pipette out and kept into another series of test tube. Then volume in all the tubes was made up to 1 mL with distilled water. Two tube with 1 mL of water each was served as the blank. Later on, 5 mL of solution C was mixed well and incubated at room temperature for 10 min. Thereafter, 0.5 mL of Folin Ciocaltu Reagent (FCR) was mixed well immediately and incubated at room temperature in dark for 30 min. The absorbance at 660 nm against the blank was read and standard graph was drawn to calculate the amount of protein in sample.
Phenol estimation: The accumulation of phenols in different wheat varieties was estimated as the method developed by Bray and Thrope (2006) with slight modification. In this method, the total phenol estimation was carried out with Folin Ciocaltu Reagent (FCR) which was measured at 650 nm calorimetrically. For estimations, 1 g of leaf sample of different wheat varieties was ground in pestle and mortar subsequently by adding in 10 times volume of 80% ethanol. It was then centrifuged to homogenate the suspensions at 10,000 rpm for 20 min. Supernatant was separated and the residue was re-extracted five times volume with of 80% ethanol. Centrifuged and the supernatants were pooled. The supernatant was evaporated to dryness and residue was dissolved in 5 mL of distilled water. Different aliquots 0.2, 0.4 and 0.6, 1.0 and 1.5 mL were pipette out into test tubes and the volume in each tube was make up to 3 mL with distilled water. Subsequently, 0.5 mL of folin ciocaltu reagent was added and after 3 min, 2 mL of 20% Na2CO3 solution in each tube was thoroughly mixed. The tubes were placed in boiling water for one minute and then cooled. Then absorbance at 650 nm against a reagent blank was measured using Ultra Violet Visible (UV-VIS) spectrophotometer and the standard curve using different concentration on catechol was prepared. From the standard curve the concentration of phenol in the test sample was determined and expressed as mg phenols per gram of sample material.
RAPD analysis
Isolation and purification of total genomic DNA
Isolation of DNA: Genomic DNA from six different wheat varieties was isolated as described by Doyle and Doyle (1987) with slight modification.
About 100 mg of wheat tender leaves were harvested and quickly ground in 750 μL of grinding buffer with the help of mortar and pestle an liquid nitrogen. It was then transferred in a 2 mL eppendrof tube and incubated at 65°C in water bath for 45 min. After 45 min, 1mL of 10 M ammonium acetate was added and again incubated for 16 min. It was then centrifuged at 10,000 rpm for 10 min at 18°C. Three layers were formed. The supernatant was collected into a clean 2 mL eppendrof tube and equal volume of chilled isopropanol was added and kept at -20°C for overnight. The DNA thus precipitated was pellet at 10,000 rpm for 10 min, washed with 70% ethanol, dried and dissolve in 1.0 mL of TE buffer. The solution was transferred to eppendorf"s tube and equal volume of phenol: chloroform: isoamyl alcohol (25:24:1) was added and gently mixed. The mixture was centrifuged at 10,000 rpm for 5 min and supernatant was transferred to fresh tube. Extraction was repeated twice with equal volume of chloroform and isoamyl (24:1). Two volume of chilled ethanol was added and kept at -20°C for 1 h. The DNA thus precipitated and washed with 70% ethanol, dried and dissolved in 1 mL T E buffer.
Purification of genomic DNA: After dissolving the DNA in TE buffer, 10-15 μL of RNAse A (10 mg mL–1) per 500 μL DNA sample was added and incubated at 37°C for 1 h to degrade RNA. Equal volume of 25:24:1, phenol: chloroform: isoamyl alcohol mixture (pH 8) was added and gently mixed for 5 min and centrifuged for 10 min at 10,000 rpm. Three layers formed, the upper layer was separated into 2 mL eppendrof tubes and double volume of 100% chilled ethanol was added for DNA precipitation (DNA was seen as clumped). Precipitated DNA was then centrifuged at 10,000 rpm for 10 min at room temperature and supernatant was discarded. For easy pelleting, 3 N sodium acetate or 5 M ammonium acetate @ 1/10 of DNA volume was added. The DNA pellet was washed thrice in 70% ethanol with gentle tapping to expel salts. Pellets were dried and dissolved in 100 μL TE buffer.
Quantification of genomic DNA: Quantification of DNA was accomplished by analyzing the purified DNA on 0.8% agarose gel alongside uncut lambda DNA as standard. The DNA was diluted in TE buffer to a concentration of approximately 12.5 ng μL–1 for use in PCR analysis.
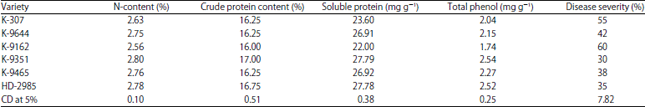
| Table 1: | Biochemical variation in mature leaves of different wheat cultivars |
 | |
| Table 2: | Soluble protein and total phenol content in different varieties of wheat seedling |
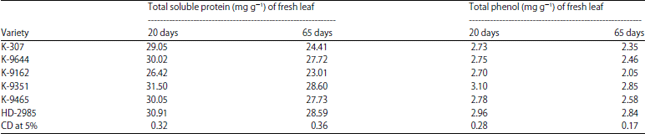 | |
Yield of purified DNA was also estimated by taking the absorbance at 260 nm, using spectrophotometer. To calculate the yield of DNA, the relationship 1 A 260 unit = 50 μg double stranded DNA was used.
PCR condition: The PCR condition were initial denaturation of DNA at 94°C for 2 min followed by 41 cycles of DNA template denaturation at 92°C for 1 min, primer annealing at 40, 42, 45, 45 and 53°C, respectively for 1 min, DNA amplification at 72°C for 3 min and final primer extension at 72°C for 10 min.
Agarose gel electrophoresis: The amplified products were separated by electrophoresis in 2.0% agarose gel containing ethidium bromide (10 mg mL–1) using 1X TBE buffer @ 50-60 V cm–1 for 3 h. The size of amplified fragments was determined by comparison with standard molecular weight markers (100 bp DNA ladder plus,). The DNA fragments were visualized in UV light and photographed using gel documentation system for permanent records.
Correlation coefficients (r) of disease severity with soluble protein and total phenol: The biochemical observations of wheat leaves of different wheat varieties and disease severity of the corresponding variety under field condition were statistically analyzed. Correlation coefficients (r) between soluble protein and disease severity and between total phenol and disease severity were calculated by standard statistical calculation. Simple regression equations (Y = a+bx) were also developed for both the variables (Protein and phenol) separately to understand their relation with disease severity.
RESULT AND DISCUSSION
Response of spot blotch by different varieties of wheat: The resistance could be assured to be one factor for the reduce of disease severity. Severity of disease was recorded in field trail under natural condition and the result presented in the Table 1 showed that the K-9162 variety showed maximum with 60% disease severity followed by K-307 with 55.00%. On the after hand, minimum disease severity was recorded in the variety K-9351 with the value of 30%, indicating the comparatively resistance to spot blotch among all the varieties. Statistical analysis of the data revealed that K-9162 had significant difference from rest of varieties (Alfano et al., 2007; Hossain and Azad, 1992; Kuc, 1987). Since we have done the experiment under same environmental condition and variable response indicating the effect variable response to spot blotch.
Biochemical variation of seedling stage
Soluble protein: Biochemical analysis of six varieties of wheat revealed that all these varieties content variable amount of soluble protein content (Table 2). The maximum soluble protein content with 31.50 mg g–1 of fresh leaf at 20 days, 28.60 mg g–1 of fresh leaf of 65 days was observed in wheat varieties K-9351 which was followed by HD-2985, showing 30.91 mg g–1 of fresh leaf and 28.59 mg g–1 of fresh leaf at 20 and 65 days, respectively. The statistical analysis of the data showed that the varieties K-9644 with K-9465 at 20 days are statistically at par. Similarly, at 65 days of observation, the variety K9644 with K 9465 and K 9351 with HD 2985 are statistically at par.
| Table 3: | Correlation of disease severity with soluble protein content and total phenol content at vegetative stage of wheat cultivar |
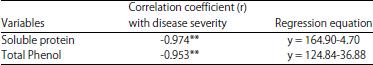 | |
| Table 4: | RAPD analysis of six varieties of wheat |
 | |
From the , it is also cleared that the soluble protein content was gradually decreased with increase the age of plant. The variety K-9162 content minimum amount of soluble protein among the varieties which is showing, 26.42 mg g–1 fresh leave at 20 days whereas at 65 days it was only 23.01 mg g–1 of fresh leave. The present findings were also supported by the worked done by Guo et al. (1986) in rice and Biswas et al. (2003) in wheat. Dwivedi (2008) was also found the different banding pattern of protein in different wheat varieties. Biochemical variation in different varieties of wheat might be one of the key factors for resistance. Mishra et al. (2011) also fund that variable resistance response to Alternaria blight by different varieties of wheat.
Total phenol: Phenol is another important parameter for governing resistant or susceptibility of host and also for determining variability among the variety. The result presented in the Table 2 showed that the total phenol content varies from variety to variety. The maximum total phenol content with 3.10 mg g–1 of fresh leaf at 20 days and 2.85 mg g–1 at 65 days was found in the variety K-9351. The minimum amount of total phenol content was found in the variety K-9162 which is 2.70 mg g–1 at 20 days and 2.05 mg g–1 at 65 days. Data from the revealed that the total phenol content maximum at 20 days then gradually decrease at 65 days showing 2.96 and 2.84 mg g–1, respectively. From the above two parameters, it is cleared that total phenol and protein content both are reduced with the increase with age of plant. Phenols are well known antifungal, antibacterial and antiviral compounds (Brown, 1964; Boller, 1985; Biehn et al., 1968). The phytoalexins involved in disease resistance are phenols in chemical constitution. Phenols are involved in disease resistance in many ways like hypersensitive cell death or lignifications of cell walls (Nicholson and Hammerschmidt, 1992; Adesh, 2008; Kumawat et al., 2008; Arzoo et al., 2012). Matern and Kneusal (1988) suggested that the first stage of defense mechanism involve a rapid accumulation of phenol at the infection site which restricts or slows the growth of the pathogen (Chaurasia et al., 1999, 2000).
Correlation between disease severity and soluble protein content in wheat cultivar: The correlation between disease severity and soluble protein content at vegetative stage of wheat varieties revealed that there was negative correlation (r) = -0.974. The regression equation of soluble protein and disease severity showed the correlation coefficient y = 164.90-4.70. Phenol is another important parameter for resistance response in plant. The high content of phenol indicates lower disease severity. There was also negative correlation between disease severity and total phenol content that is (r) = -0.953. The regression equation of total phenol content and disease severity showed that correlation coefficient y = 124.84-36.88 (Table 3). The negative correlation co-efficient between soluble protein and total phenol with disease incidence were also found in rice against brown leaf spot (Kumawat et al., 2010), in tomato against Fusarium wilt (Kumar and Biswas, 2010; Uddin et al., 2006; Attitalla et al., 2001; Larkin and Fravel, 1999; Fuchs et al., 1997; Baysal et al., 2002; Ramamoorthy et al., 2002), in wheat against spot blotch (Mishra et al., 2011; Maraite et al., 1998; Prabhu and Prasada, 1966).
RAPD analysis: The present finding revealed that morphological pathological and biochemical variability are exist among the six varieties of wheat. But for further confirmation, variability at molecular level has been done through RAPD (Mullis, 1990; Gepts, 1993; Astarini et al., 2004). The experimental finding revealed that there are exist polymorphism among the six varieties (Table 4). In recent year, molecular markers have received considerable attention and have been extensively used for analysis of the genetic diversity, phylogenetic evolution, mapping and tagging genes of agronomically importance plant species. The present findings have also been made the genetic variability of six hexapliod Indian wheat varieties through RAPD analysis (Fig. 1).
The introduction of molecular markers in plant breeding has presented a valuable tool for the characterization of genetic materials. Among them, the RAPD markers have been successfully used in wheat germplasm evaluation because of their many advantages. The suitability of the RAPD technique for genetic diversity studies and germplasm evaluations has been shown in many crops (Erlich et al., 1991).
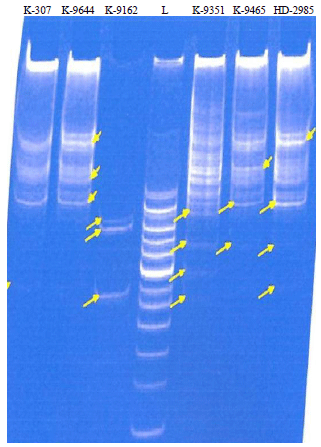 | |
| Fig. 1: | DNA fragment resolved by RAPD analysis for different varieties of wheat with 100 bp DNA Ladder gene ruler (L) |
The RAPD technique is quick (Colombo et al., 1998; Mukhtar et al., 2002), cost effective (Fahima et al., 1999) and to perform analysis without need for prior sequencing of the genome (Hongtrakul et al., 1997; Migdadi et al., 2006). Although major bands from RAPD reactions are highly reproducible, minor bands can difficult to repeat due to random priming nature of this PCR reaction and potential confounding effects associated with co-migration with other markers (Tessier et al., 1999). Replication slippage is thought to occur more frequently than single nucleotide mutations and insertion/deletion events, which generated the polymorphisms detected by RAPD analysis (Powell et al., 1996). Various numbers of primers have been used in the study of different species of the genus Triticum that revealed various degrees of polymorphism.
CONCLUSION
The present study showed that highest numbers of fragments are found in variety K-9351 which is 13. On the other hand minimum numbers of fragments were found in K-9162 (3). The variety K-9465 and HD-2985 were showing same number of fragment self different in their size. The presence or absence of fragment and their variable size might be the responsible for morphological and biochemical variability along with resistance response to spot blotch. Electrophoresis of PCR produced on 2% agarose gels containing ethidium bromide revealed that the number of fragment present in each variety ranged from 3-13. The primer ITS 3 (5' - GCATCGATGAAGAACGCAGC -3') produced fragment ranges from 25-500 bp in size.
REFERENCES
- Alfano, G., M.L. Ivey, C. Cakir, J.I.B. Bos and S.A. Miller et al., 2007. Systemic modulation of gene expression in tomato by Trichoderma hamatum 382. Phytopathology, 97: 429-437.
CrossRefDirect Link - Astarini, I.A., J.A. Plummer, R.A. Lancaster and G. Yan, 2004. Fingerprinting of cauliflower cultivars using RAPD markers. Aust. J. Agric. Res., 55: 117-124.
CrossRefDirect Link - Attitalla, I.H., P. Johnson, S. Brishammar and P. Quintanilla, 2001. Systemic resistance to Fusarium wilt in tomato induced by Phytopthora cryptogea. J. Phytopathol., 149: 373-380.
CrossRefDirect Link - Baysal, O., P. Laux and W. Zeller, 2002. Further studies on the Induced Resistance (IR) effect of plant extract from Hedera helix against fire blight (Erwinia amylovora). Acta Hort., 590: 273-277.
CrossRefDirect Link - Biswas, S.K., K.D. Srivastava, R. Aggarwal, S. Praveen and D.V. Singh, 2003. Biochemical changes in wheat induced by Chaetomium globosum against spot blotch pathogen. Indian Phytopathol., 54: 374-379.
Direct Link - Chaurasia, S., R. Chand and A.K. Joshi, 2000. Relative dominance of Alternaria triticina Pras. et Prab. and Bipolaris sorokiniana (Sacc.) shoemaker in different growth stages of wheat (T. aestivum L.). J. Plant Dis. Prot., 107: 176-181.
Direct Link - Chaurasia, S., A.K. Joshi, R. Dhari and R. Chand, 1999. Resistance to foliar blight of wheat: A search. Genet. Resour. Crop Evol., 46: 469-475.
CrossRefDirect Link - Colombo, C., G. Second, T.L. Valle and A. Charrier, 1998. Genetic diversity characterization of cassava cultivars (Manihot esculenta Crantz). I) RAPD markers. Genet. Mol. Biol., 21: 69-84.
CrossRefDirect Link - Doyle, J.J. and J.L. Doyle, 1987. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull., 19: 11-15.
Direct Link - Erlich, H.A., A.D. Gelfand and J.J. Sninsky, 1991. Recent advances in the polymerase chain reaction. Science, 252: 1643-1651.
CrossRefDirect Link - Fahima, T., G.L. Sun, A. Beharav, T. Krugman, A. Beiles and E. Nevo, 1999. RAPD polymorphism of wild emmer wheat populations, Triticum dicoccoides, in Israel. Theor. Appl. Genet., 98: 434-447.
CrossRefDirect Link - Fuchs, J.G., Y. Moenne-Loccoz and G. Defago, 1997. Nonpathogenic Fusarium oxysporum strain Fo47 induces resistance to Fusarium wilt in tomato. Plant Dis., 81: 492-496.
CrossRefDirect Link - Joshi, A.K., G. Ortiz-Ferrara, J. Crossa, G. Singh, R.C. Sharma, R. Chand and R. Prasad, 2007. Combining superior agronomic performance and terminal heat tolerance with resistance to spot blotch (Bipolaris sorokiniana) of wheat in the warm humid gangetic plains of South Asia. Field Crops Res., 103: 53-61.
CrossRefDirect Link - Arzoo, K., S.K. Biswas and M. Rajik, 2012. Biochemical evidences of defence response in tomato against Fusarium wilt induced by plant extracts. Plant Pathol. J., 11: 42-50.
CrossRefDirect Link - Kumawat, G.L., S.K. Biswas and S.S.L. Srivastava, 2008. Biochemical evidence of defense response in paddy induced by bio-agents against brown leaf spot pathogen. Indian Phytopathol., 61: 197-203.
Direct Link - Kumawat, G.L., S.K. Biswas and M. Rajik, 2010. Antagonistic evaluation of Trichoderma spp. And there effect of on seed germination and growth of paddy seedling. J. Plant Dis. Sci., 5: 203-207.
Direct Link - Hongtrakul, V., G.M. Huestis and S.J. Knapp, 1997. Amplified fragment length polymorphisms as a tool for DNA fingerprinting sunflower germplasm: Genetic diversity among oilseed inbred lines. Theor. Applied Genet., 95: 400-407.
CrossRefDirect Link - Hossain, I. and A.K. Azad, 1992. Reaction of wheat to Helminthosporium sativum in Bangladesh. Hereditas, 116: 203-205.
CrossRefDirect Link - Larkin, R.P. and D.R. Fravel, 1999. Mechanisms of action and dose-response relationships governing biological control of Fusarium wilt of tomato by nonpathogenic Fusarium spp. Phytopathology, 89: 1152-1161.
CrossRefDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Matern, U. and R.E. Kneusel, 1988. Phenolic compounds in plant disease resistance. Phytoparasitica, 16: 153-170.
CrossRefDirect Link - Mishra, V.K., S.K. Biswas and M. Rajik, 2011. Biochemical mechanism of resistance to alternaria blight by different varieties of wheat. Int. J. Plant Pathol., 2: 72-80.
CrossRefDirect Link - Mukhtar, M.S., M.U. Rahmanw and Y. Zafar, 2002. Assessment of genetic diversity among wheat (Triticum aestivum L.) cultivars from a range of localities across Pakistan using Random Amplified Polymorphic DNA (RAPD) analysis. Euphytica, 128: 417-425.
CrossRefDirect Link - Nasser, W., M. de Tapia and G. Burkard, 1990. Maize pathogenesis-related proteins: Characterization and cellular distribution of 1,3-β-glucanases and chitinases induced by brome mosaic virus infection or mercuric chloride treatment. Physiol. Mol. Plant Pathol., 36: 1-14.
CrossRefDirect Link - Porter, W.M. and D.H. Smith, 1982. Detection of identification errors in germplasm collections. Crop Sci., 22: 701-703.
CrossRefDirect Link - Powell, W., M. Morgante, C. Andre, M. Hanafey, J. Vogel, S. Tingey and A. Rafalski, 1996. The comparison of RFLP, RAPD, AFLP and SSR (microsatellite) markers for germplasm analysis. Mol. Breed., 2: 225-238.
CrossRefDirect Link - Ramamoorthy, V., T. Raguchander and R. Samiyappan, 2002. Induction of defense-related proteins in tomato roots treated with Pseudomonas fluorescens Pf1 and Fusarium oxysporum f. sp. lycopersici. Plant Soil, 239: 55-68.
CrossRefDirect Link - Shetty, H.S. and R. Ahmad, 1980. Changes in phenolic content of sorghum and maize cultivars resistant and susceptible to sorghum downy mildew. Curr. Sci., 49: 439-441.
Direct Link - Sinha, B., R.M. Singh and U.P. Singh, 1991. Genetics of leaf blight resistance in wheat. Theor. Applied Genet., 82: 399-404.
CrossRefDirect Link - Snyder, W.C. and H.W. Hansen, 1940. The species concept in Fusarium. Am. J. Bot., 27: 64-67.
Direct Link - Tessier, C., J. David, P. This, J.M. Boursiquot and A. Charrier, 1999. Optimization of the choice of molecular markers for varietal identification in Vitis vinifera L. Theor. Applied Genet., 98: 171-177.
Direct Link - Uddin, S.A., K.M. Khalequzzaman and A.B. Rashid, 2006. Effect of relative humidity on the development of head blight by Bipolaris sorokiniana in wheat. J. Agric. Rural Dev., 4: 61-65.
CrossRefDirect Link - Van Loon, L.C., P.A.H.M. Bakker and C.M.J. Pieterse, 1998. Systemic resistance induced by rhizosphere bacteria. Annu. Rev. Phytopathol., 36: 453-483.
CrossRefPubMedDirect Link - Nicholson, R.L. and R. Hammerschmidt, 1992. Phenolic compounds and their role in disease resistance. Annu. Rev. Phytopathol., 30: 369-389.
CrossRefDirect Link