
K.M. Abd El-Hai
Plant Pathology Research Institute, Agricultural Research Center, Giza, Egypt
M.A. El-Metwally
Plant Pathology Research Institute, Agricultural Research Center, Giza, Egypt
LiveDNA: 20.359
Naglaa T. Mohamed
Plant Pathology Research Institute, Agricultural Research Center, Giza, Egypt
Plant Pathology Journal
Year: 2016 | Volume: 15 | Issue: 2 | Page No.: 17-26







ABSTRACT
Isolation of pathogenic fungi from both cultivars of diseased lupine was carried out in five districts of Dakahlia governorate. The high frequency isolated fungi presented in Temi El-Amdeed followed by Bani-Ebeed district. Fusarium solani and F. oxysporum proved to be the most dominate isolated followed by Rhizoctonia solani . In greenhouse, Giza 1 was high susceptible cultivar for infected with root rot pathogenic fungi. Sclerotium rolfsii followed by R. solani then F. solani were the most aggressive damping-off disease. In the field experiment, Giza 2 cultivar was the best in germination% and more tolerant of damping-off than Giza 1. The application of Rhizolex-T50 followed by H2O2 at low concentrate (0.50 mM) showed a highest percentage of germination within lowest percentage of damping-off. With considerable that, no significant differences between Rhizolex-T50 and low concentrate of H2O2. The high photosynthetic pigments and phenolic content were obtained from the application of acetylsalicylic acid (ASA) at moderate concentrate (15 mM) in both cultivars. Giza 2 gave the highest values in these parameters. Soaking both cultivars of lupine seeds in both tested materials increased significantly growth parameter, yield components and seed quality. The moderate concentration of ASA (15 mM) was the most effective followed by the low concentration of H2O2 (0.50 mM). The data suggested that the application of H2O2 at 0.50 mM and ASA at 15 mM as seed soaking could be considered as fungicide alternatives for controlling lupine root rot disease as well as improve growth and productivity.
PDF Abstract XML References Citation
Received: November 26, 2015;
Accepted: January 21, 2016;
Published: March 15, 2016
How to cite this article
K.M. Abd El-Hai, M.A. El-Metwally and Naglaa T. Mohamed, 2016. Hydrogen Peroxide and Acetylsalicylic Acid Induce the Defense of
Lupine Against Root Rot Disease. Plant Pathology Journal, 15: 17-26.
DOI: 10.3923/ppj.2016.17.26
URL: https://scialert.net/abstract/?doi=ppj.2016.17.26
DOI: 10.3923/ppj.2016.17.26
URL: https://scialert.net/abstract/?doi=ppj.2016.17.26
INTRODUCTION
Lupine (Lupinus termis Forsk) is one of the most important crop which belonging to Fabaceae family. Lupine, as other fabaceaus seeds is good dietary sources of minerals (Trugo et al., 1993). Lupine seeds also contains chemical compounds i.e., protein, oil, cholesterol and alkaloids (lupulin. Luponine, lupul and sparateine). lupulin is occasionally employed as stomachic tomic. Seeds can be eaten when the bitter components have been removed. Also, the seeds roasted can make a coffee substitute. Therefore, lupine is cultivated in Egypt for food, medical and industrial purposes (Chiej, 1984; Ibrahim et al., 1990). As well as, lupine plants used in sustainable and environment-friendly agriculture because of its high potential for biological nitrogen fixation (Robinson et al., 2000).
Damping-off and root rot diseases are among the most destructive diseases attacking lupine in Egypt. Several pathogens such as Rhizoctonia solani, Sclerotium rolfsii, Fusarium solani and F. oxysporum attacking lupine seeds, root and stem base causing serious losses in seed germination and plant stand (Abd-El-Kareem et al., 2004; El Mougy, 2004; Ali et al., 2009).
The application of fungicides is considered one of the most famous environmental pollutions. Therefore, it is urgent to alternative safe efficient methods against plant diseases. Induced resistance of plants against pathogens can be defined as the process of active resistance depended on the host plants physical or chemical barriers activated by abiotic and biotic agents. These agents sensitizes the plant to respond rapid after infection include phytoalexin accumulation, phenols, lignifications and activation of peroxidase, polyphenoloxidase, catalase and chitinase (Meena et al., 2001; Mahmoud et al., 2006; Walters, 2007).
Some abiotic inducers i.e., acetylsalicylic acid (ASA) on lupine and hydrogen peroxide (H2O2) on lentil and peanut have been shown to induce resistance in plants against damping-off and root rot diseases (El Mougy, 2004; Morsy, 2005; Mahmoud et al., 2006).
This study aimed to study the effect of abiotic (ASA and H2O2) inducers in reducing of lupine root rot diseases and enhancing of some morphological and physiological characters as well as yield and seed quality.
MATERIALS AND METHODS
Source of lupine seeds: Seed of two lupine cultivars (Giza 1 and Giza 2) were obtained from Legume Crop Research Department, Field Crop Research Institute, Agriculture Research Center, Giza, Egypt.
Abiotic inducers: Two abiotic chemical inducers namely, hydrogen peroxide (H2O2) at 0.50, 0.75 and 1.0 mM and acetylsalicylic acid (ASA) at 10, 15 and 20 mM were used as seed soaking to study their effects in inducing resistance in lupine plant against root-rot diseases.
Isolation, purification, identification of the causal pathogens: The causal pathogens were isolated from lupine plants showing typical symptoms of root rot disease from different districts of Dakahlia governorate. The infected roots were washed thoroughly with tap water, cut into small pieces (1 cm) and surface disinfested with sodium hypochlorite 2% for 2 min. Then re-washed several times with sterilized water, dried between folds of sterilized filter paper and were placed onto Potato Dextrose Agar (PDA) medium in petri-dishes supplemented with streptomycin sulfate (100 μg mL–1). Petri-dishes were incubated at 21°C for five days. The developed fungal colonies purified and identified according to Ellis (1976), Sneh et al. (1992) and Nelson et al. (1983).
Fungal inoculums preparation: Inocula of Rhizoctonia solani, Sclerotium rolfsii, Fusarium solani and Fusarium oxysporum were prepared by growing each fungus on sorghum: coarse sand medium (1:1w/w and 40% water) for two weeks at 25±1°C according to Filonow et al. (1988).
Pathogenicity test: The previously prepared fungal in inocula were tested for their pathogenicity on lupine under greenhouse conditions. Inoculum of each isolate was mixed thoroughly with autoclaved soil in plastic pots (25 cm diam.) at the rate of 5% by weight (Abdel-Kader, 1997). Four pots were used as replicates for each fungus as well as check (uninfested soil). Healthy lupine seeds for tow cultivars (Giza 1 and Giza 2) were sown after surface sterilized at the rate of 6 seeds/pot. The percentage of root rot disease incidence was calculated as pre- and post-emergence damping off after 15 and 40 days of sowing, respectively.
Field experiments: Two field experiments were carried out at Tag El-Ezz, Agriculture Research Station, Dakahlia, Egypt during 2012/2013 and 2013/2014 seasons.
Lupine seeds were soaked for 3 h in abiotic inducers (H2O2 at 0.50, 0.75 and 1.0 mM and ASA at 10, 15 and 20 mM) while, Rhizolex-T 50 w.p. was used as seed coating at the rate of 3 g kg–1 seeds. Treated lupine seeds (cvs. Giza 1 and Giza 2) were sown in 30th and 10th of November in two seasons, respectively and left under natural infection. A split plot design with three replicates was used in these experiments. The main plots were occupied by cultivars, while sub-plots were occupied by treatments. The area of each sub-plot was 3×3.5 m, sowing with 180 seeds.
Germination and disease assessment: Germination percentage and pre-emergence damping-off were recorded at 20 days from sowing while post-emergence damping-off was determined at 80 days from sowing.
Physiological characters: At 75 days from sowing, photosynthetic pigments (chlorophyll a, b and carotenoids) were extracted in methanol 90% from the blade of the third leaf from plant tip (terminal leaflet) according to Robinson and Britz (2000) then determined spectrophotometrically according to Mackinney (1941).
Total phenolic compounds were determined in fresh shoot using the Folin-ciocalteau reagent according to Malik and Singh (1980).
Morphological and yield characters: At harvest, 175 days from sowing for Giza 1 cultivar and 160 days for Giza 2 cultivar, samples were taken to estimate plant height, number of branches and leaves per plant, number of pods, plant yield and weight of 100-seed.
Seed quality: In the second season, lupine seeds were dried at 70°C for 48 h, grounded and analyzed for alkaloid lupinine (Dabbas, 1973) and total nitrogen by semi-micro-Kjldahle (Pregl, 1945). Protein percentage was calculated by multiplying the N% by 6.25.
Statistical analysis: All data were statistically analyzed by the Software; CoStat (2005) in consultation with the analysis of variance (Gomez and Gomez, 1984).
RESULTS
Isolation of pathogenic fungi: Infected lupine cvs. Giza 1 and Giza 2 with typical symptoms of root rot diseases collected from different districts of Dakahlia governorate, Egypt are shown in Table 1. It was observed that Giza 1 cultivar was high susceptible for infected with root rot pathogenic fungi except, Rhizoctonia solani as compared with Giza 2 cultivar. The high frequency isolated fungi were found in Temi El-Amdeed district followed by Bani-Ebeed then Senblaween, while Dekerns came late. Fusarium solani was isolated at high percentage followed by F. oxysporum then Rhizoctonia solani.
Pathogenicity testes: Data presented in Table 2 show that Giza 1 lupine cultivar was more sensitive to the infection of pre- and post-emergence damping-off than Giza 2 cultivar.
| Table 1: | Frequency of the isolated fungi from lupine roots at different districts in Dakahlia governorate |
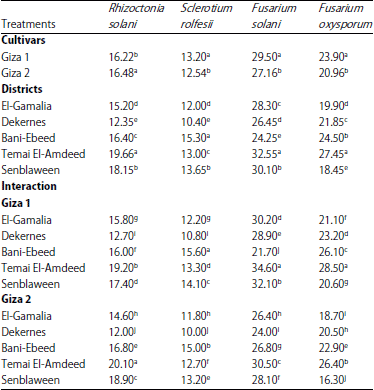 | |
| Mean within a column followed by the same letter(s) is not significantly different according to Duncan’s multiple range tests | |
| Table 2: | Pathogenicity test of isolated fungi from lupine plants under greenhouse conditions |
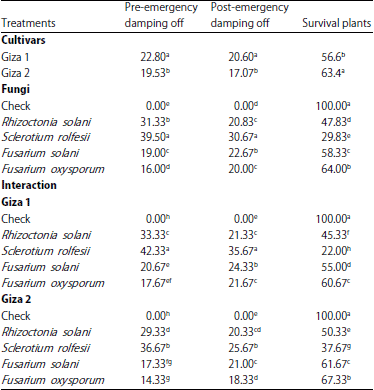 | |
| Mean within a column followed by the same letter(s) is not significantly different according to Duncan’s multiple range tests | |
| Table 3: | Effect of inducers on germination percentage and damping off disease of lupine plants under field conditions |
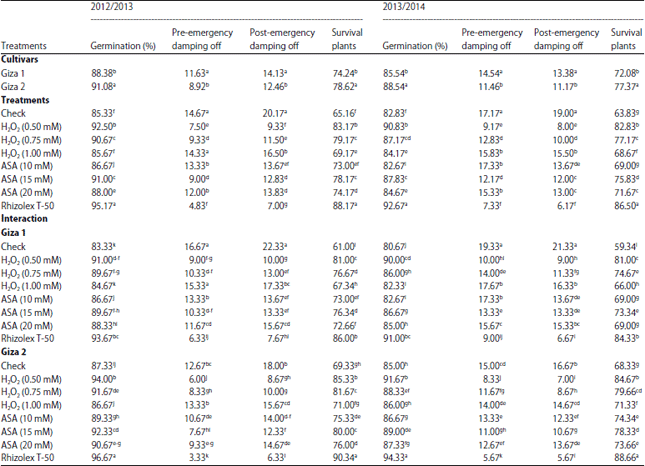 | |
| Mean within a column followed by the same letter(s) is not significantly different according to Duncan’s multiple range tests | |
Generally, Sclerotium rolfsii showed highest percentage of pre- and post-emergence damping-off in both lupine cultivars than other pathogenic fungi. Rhizoctonia solani came second followed by F. solani then F. oxysporum. With considerable that, all tested fungi were pathogenic and causes typical symptoms of pre- and post-emergence damping-off of lupine seedlings.
Field experiments
Germination and disease assessment: Data of germination percentage and damping-off of lupine plants as affected by inducers under field conditions were recorded in Table 3. Giza 2 cultivar was the best in generation% and was more tolerant of damping-off than Giza 1. Soaking of lupine seeds in each one of both inducers significantly increased germination percentage within decreased pre and post-emergence damping-off in both seasons compared with check.
Concerning the effects of treatments and its interacted with cultivars, data show that Rhizolex-T50 was the most effective followed by H2O2 then ASA in both cultivars. The low concentration of H2O2 (0.50 mM) was more effective in this respect. It is worthy to mention that there is no significant difference between H2O2 at 0.50 mM and Rhizolex-T50 treatments.
Physiological characters: Photosynthetic pigments and total phenols are not only a good parameters reflecting the health conditions of plant but also, carotenoids and phenols are known that a highly effective antioxidants. As shown in Table 4, Giza 2 cultivar gave the highest values of photosynthetic pigments (Chl. a, b and carotenoids) and total phenol content as compared with Giza 1 cultivar. There is a positive relationship among chlorophyll a, b and total phenols content. Both tested inducers increased significantly photosynthetic pigment and phenols. The maximum increase in chlorophyll a and b as well as phenolic content occurred under the application of ASA followed by H2O2.
| Table 4: | Effect of inducers on germination percentage and damping off disease of lupine plants under field conditions |
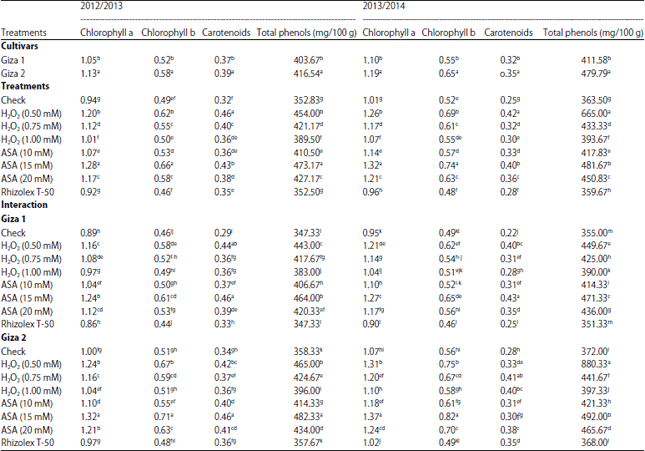 | |
| Mean within a column followed by the same letter(s) is not significantly different according to Duncan’s multiple range tests | |
The moderate concentrate of ASA (15 mM) was more effective. Whilst, Rhizolex-T50 had no significant effect on photosynthetic pigments and total phenols in lupine plants. On the other side, the highest increase in carotenoids content was observed with low level of H2O2 followed by moderate level of ASA.
Lupine growth and yield components: As shown in Table 5 and 6, there were significant differences between treatments of both lupine cultivars regarding lupine growth (plant height, branches and leaves number per plant) and yield components (number of pods/plant, plant yield and weight of 100-seeds).
Data in Table 6 show that Giza 2 cultivar recorded the highest values of plant height, branches and leaves number per plant. Soaking lupine seeds in both tested inducers increased significantly plant height, number of branches and leaves/plant in both cultivars during the two growing seasons. Acetylsalicylic acid at 15 mM appeared excellent superiority in all treatments on plant height, number of branches and leaves/plant followed by H2O2 at 0.50 mM.
Data concerning yield components in relation to the effect of tested inducers are present in Table 6. It can easily notice that Giza 2 cultivar gave the highest average of pods number per plant, plant yield and weight of 100-seed.
Moreover, all treatments increased significantly the same parameters in both cultivars. Generally, the low concentration of H2O2 and the moderate concentration of ASA lead to the highest values. In this respect ASA at 15 mM was the most effective followed by H2O2 at 0.50 mM. Meanwhile, Rhizolex-T50 had no significantly effect on the pervious parameters when compared with check.
Seed quality: Lupine seed quality was determined as protein and lupinine (alkaloids) percentages in the second season only. Data in Table 7 show that Giza 2 cultivar seeds were contains protein percentage more than Giza 1 while, Giza 1 contains lupinine percentage more than Giza 2. The maximum values of protein and lupinine in both lupine cultivars occurred under the application of ASA at moderate concentration followed by H2O2 at 0.50 mM.
| Table 5: | Effect of inducers on some morphological characters of lupine plants under field conditions |
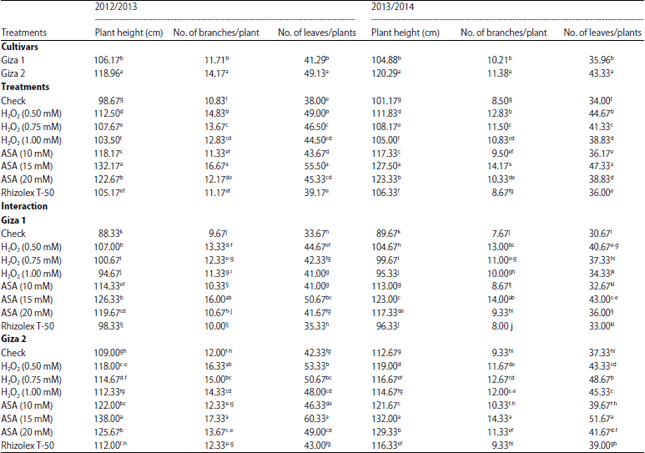 | |
| Mean within a column followed by the same letter(s) is not significantly different according to Duncan’s multiple range tests | |
DISCUSSION
Abiotic inducers are considered one of the alternative methods to decrease the use of fungicides in plant disease control. Soaking lupine seeds in both inducers, especially at low concentration of H2O2 and moderate ASA concentration gave a significant effects in reducing percentage of disease parameters, in turn increasing% of healthy survival plants. The role of H2O2 in induced disease resistance may be due to activation of peroxidase, polyphenol oxidase, catalase and B-1, 3-glucanase enzymes, which protect plants against pathogen infection (Morsy, 2005; Khalifa et al., 2007). Moreover, Martinez et al. (2000) stated that H2O2 positively influences one the local and systemic accumulation of salicylic acid which correlated with enhancement of peroxidase activity. Hydrogen peroxide also increased lignin and suberin content as well as activated of peroxidase and chitinase enzymes (Wu et al., 1997), which activities the defense mechanisms. In addition to, H2O2 inhibites pathogens directly and/or it may generate other reactive free radicals that are antimicrobial (Peng and Kuc, 1992). Hydrogen peroxide at lowest concentration (0.25%) enhancement the activity of oxidative enzymes and increases the content of phenols compounds (Mahmoud et al., 2006). On contrast, increasing of hydrogen peroxide concentration led to decrease its positively effect due to the role of H2O2 in rapid generation of Active Oxygen Species (AOS) called the oxidative burst (Lamb and Dixon, 1997). Active Oxygen Species (AOS) gives opposite effect on physiological processes in plants in increased its concentration, especially the role of hydrogen peroxide in accumulation of SA (Martinez et al., 2000). While, Lu and Higgins (1999) stated that H2O2 may remarkably inhibit the growth of pathogenic fungi and that H2O2 concentration effective in killing the fungus is considerably lower than the concentration causing plant cell death. Some studies have shown that acting at a relatively low concentration of H2O2 could be a factor inducing the expression of defence-related genes, including genes coding for catalase (Polidoros and Scandalios, 1999; Guan and Scandalios, 2000).
| Table 6: | Effect of inducers on some yield components of lupine plants under field conditions |
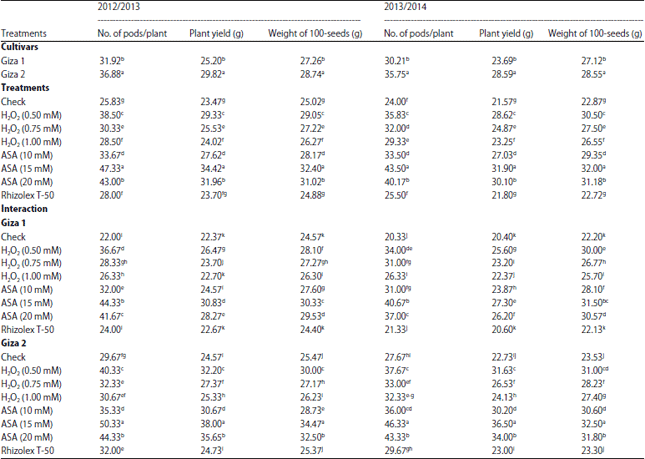 | |
| Mean within a column followed by the same letter(s) is not significantly different according to Duncan’s multiple range tests | |
Moreover, Levine et al. (1994) suggested that H2O2 directly or indirectly, plays as a signal for inducing systemic acquired resistance. Hydrogen peroxide and other activated oxygen species in the plant cell wall and in plasma membrane is often considered to be a defensive oxidative barrier to phytopathogenic fungi (Merzlyak et al., 1990; Galal and Abdou, 1996).
The present investigation revealed that ASA increased lupine germination percentage and decreased per- and post-emergence damping-off. These results are in harmony with Zhang et al. (1999), who stated that the addition of SA and ASA on wheat seeds not only increase germination rate but also increase germination and activities of alpha-amylase and proteinase in endosperm and their contents of soluble sugars, protein and free amino acids. Rizolex-T decrease root rot incidence due to the expected degradation of fungicide when introduced into the soil and exposed to the environmental conditions (Abdel-kader, 1997). Treated lupine seeds with ASA or Rizolex-T provide such protection to seed bed region against soil-borne pathogens reflected on the observed lower disease incidence at pre- emergence stage before exposure to degradation factors (El Mougy, 2004). Acetylsalicylic acid reduced lupine root rot incidence might be attributed to the act of ASA as plant defense inducers or to their direct effect on soil borne plant pathogens (El Mougy, 2002). Also, ASA induced resistance in various plants is associated with enhancing the activities of chitinase and B-1, 3-glucanase which hydrolysis hyphal cell wall of fungi (Matta et al., 1988). The effect of ASA on damping-off decreased with increasing concentration from 15-20 mM may be due to the damage effects of SA at high concentration on physiological processes, includes inhibited phosphorus uptake and potassium absorption (Harper and Balke, 1981). In addition to, it caused the collapse of the transmembrane electrochemical potential of mitochondria which had effect on ATP-production (Macri et al., 1986).
| Table 7: | Effect of inducers on protein (%) and lupinine (%) of lupine seeds under field conditions |
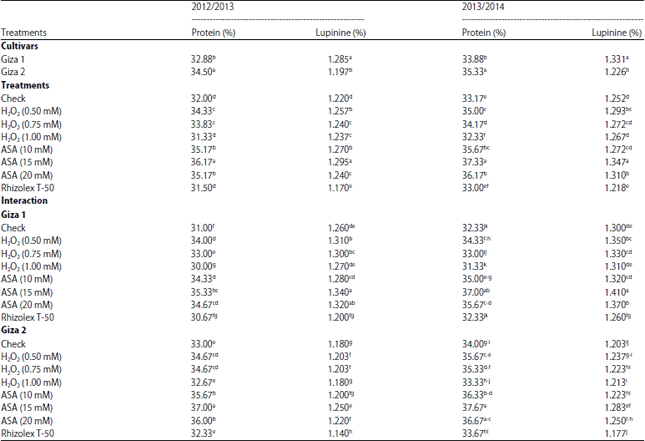 | |
| Mean within a column followed by the same letter(s) is not significantly different according to Duncan’s multiple range tests | |
Generally, the antimicrobial effect of inducers may be due to one or more the following reasons: a) inhibit the functions of several enzymes by the oxidized compounds, b) dissolve in membrane lipids and interfere with membrane functions, c) interfere with the synthesis of protein, RNA and DNA and, d) act on the sites and number of hydroxyl groups on the phenol compounds which increase toxicity to microorganisms (Nesci et al., 2003).
In the present study, it could be stated that both inducers had a stimulatory effects on photosynthetic pigments, phenol content, lupine plant growth and yield components as well as seed quality. These effect may be due to the increase in photosynthesis process, consequently increase carbohydrate content. Carbohydrates includes cellulose, hemicelluloses and pectin which consider as a barrier against pathogen invasion (Hahlbrock and Scheel, 1989). They added that, phenolic compounds are associated with structural carbohydrates, which play major role in plant defense. Moreover, Morkunas et al. (2005) indicated that soluble carbohydrates my be involved in the mechanism of resistance, because it can be used as carbon skeletons for synthesis of isoflavonoids, which are important elements of the defense system of legumes.
CONCLUSION
It could be concluded that application of hydrogen peroxide at 0.50 mM and acetylsalicylic acid at 15 mM as seed soaking is recommended for reducing root rot in lupine plants as well as improving growth, yield and seed quality.
REFERENCES
- Ali, A.A., K.M. Ghoneem, M.A. El-Metwally and K.M. Abd El-Hai, 2009. Induce systemic resistance in lupine against root rot diseases. Pak. J. Biol. Sci., 12: 213-221.
CrossRefPubMedDirect Link - El Mougy, N.S., 2004. Preliminary evaluation of salicylic acid and acetylsalicylic acid efficacy for controlling root rot disease of lupin under greenhouse conditions. Egypt. J. Phytopathol., 32: 11-21.
Direct Link - Filonow, A.B., H.A. Melouk, M. Martin and J. Sherwood, 1988. Effect of calcium sulfate on pod rot of peanut. Plant Dis., 72: 589-593.
Direct Link - Guan, L.M. and J.G. Scandalios, 2000. Hydrogen peroxide-mediated catalase gene expression in response to wounding. Free Radical Biol. Med., 28: 1182-1190.
CrossRefDirect Link - Wu, G., B.J. Shortt, E.B. Lawrence, J. Leon and K.C. Fitzsimmons et al., 1997. Activation of host defense mechanisms by elevated production of H2O2 in transgenic plants. Plant Physiol., 115: 427-435.
PubMedDirect Link - Hahlbrock, K. and D. Scheel, 1989. Physiology and molecular biology of phenylpropanoid metabolism. Annu. Rev. Plant Physiol. Plant Mol. Biol., 40: 347-369.
CrossRefDirect Link - Harper, J.R. and N.E. Balke, 1981. Characterization of the inhibition of K+ absorption in oat roots by salicylic acid. Plant Physiol., 68: 1349-1353.
Direct Link - Levine, A., R. Tenhaken, R. Dixon and C. Lamb, 1994. H2O2 from the oxidative burst orchestrates the plant hypersensitive disease resistance response. Cell, 79: 583-593.
CrossRefPubMedDirect Link - Lu, H. and V.J. Higgins, 1999. The effect of hydrogen peroxide on the viability of tomato cells and of the fungal pathogen Cladosporium fulvum. Physiol. Mol. Plant Pathol., 54: 131-143.
CrossRefDirect Link - Mackinney, G., 1941. Absorption of light by chlorophyll solutions. J. Biol. Chem., 104: 315-322.
Direct Link - Macri, F., A. Vianello and S. Pennazio, 1986. Salicylate-collapsed membrane potential in pea stem mitochondria. Physiol. Planta., 67: 136-142.
CrossRefDirect Link - Mahmoud, E.Y., S.Y.M. Shokry and Z.N. Hussin, 2006. Induction of resistance in peanut plants against root rot diseases under greenhouse conditions by some chemical inducers. J. Agric. Sci. Mansoura Univ., 31: 3511-3524.
Direct Link - Martinez, C., J.C. Baccou, E. Bresson, Y. Baissac and J.F. Daniel et aal., 2000. Salicylic acid mediated by the oxidative burst is a key molecule in local and systemic responses of cotton challenged by an avirulent race of Xanthomonas campestris pv malvacearum. Plant Physiol., 122: 757-766.
CrossRefDirect Link - Matta, A., G.I. Abattista and L. Ferraris, 1988. Stimulation of 1,3-β-glucanase and chitinase by stresses that induce resistance to Fusarium wilt in tomato. Phytopathol. Medit., 27: 45-50.
Direct Link - Merzlyak, M.N., I.V. Reshetnikova, O.B. Chivkunova, D.G. Ivanova and N.I. Maximova, 1990. Hydrogen peroxide- and superoxide-dependent fatty acid breakdown in Phytophthora infestans zoospores. Plant Sci., 72: 207-212.
CrossRefDirect Link - Morkunas, I., L. Marczak, J. Stachowiak and M. Stobiecki, 2005. Sucrose-induced lupine defense against Fusarium oxysporum: Sucrose-stimulated accumulation of isoflavonoids as a defense response of lupine to Fusarium oxysporum. Plant Physiol. Biochem., 43: 363-373.
CrossRefPubMedDirect Link - Morsy, K.M.M., 2005. Induced resistance against damping-off, root rot and wilt diseases of lentil. Egypt. J. Phytopathol., 33: 53-63.
Direct Link - Nesci, A., M. Rodriguez and M. Etcheverry, 2003. Control of Aspergillus growth and aflatoxin production using antioxidants at different conditions of water activity and pH. J. Applied Microbiol., 95: 279-287.
CrossRefDirect Link - Peng, M. and J. Kuc, 1992. Peroxidase generated hydrogen peroxide as a source of antifungal activity in vitro and on Tobacco leaf disks. Phytopathology, 82: 696-699.
Direct Link - Polidoros, A.N. and J.G. Scandalios, 1999. Role of hydrogen peroxide and different classes of antioxidants in the regulation of catalase and glutathione S-transferase gene expression in maize (Zea mays L.). Physiol. Plant., 106: 112-120.
CrossRefDirect Link - Robinson, J.M. and S.J. Britz, 2000. Tolerance of a field grown soybean cultivar to elevated ozone level is concurrent with higher leaflet ascorbic acid level, higher ascorbate-dehydroascorbate redox status and long term photosynthetic productivity. Photosynth. Res., 64: 77-87.
CrossRefDirect Link - Robinson, K.O., D.A. Beyene, P. van Berkum, R. Knight-Mason and H.L. Bhardwaj, 2000. Variability in plant-microbe interaction between Lupinus lines and Bradyrhizobium strains. Plant Sci., 159: 257-264.
CrossRefDirect Link - Trugo, L.C., C.M. Donangelo, Y.A. Duarte and C.L. Tavares, 1993. Phytic acid and selected mineral composition of seed from wild species and cultivated varieties of lupin. Food Chem., 47: 391-394.
CrossRefDirect Link - Abd-El-Kareem, F., M.A. Abd-Allah, N.G. El-Gamal and N.S. El-Mougy, 2004. Integrated control of Lupin root rot disease in solarized soil under greenhouse and field condition. Egypt. J. Phytopathol., 32: 49-63.
Direct Link - Lamb, L. and R.A. Dixon, 1997. The oxidative burst in plant disease resistance. Annu. Rev. Plant Physiol. Plant Mol. Biol., 48: 251-275.
CrossRefPubMedDirect Link