
Alkadri Dima
Department of Agricultural Sciences, Alma Mater Studiorum University of Bologna, Viale Giuseppe Fanin 40, 40127, Bologna, Italy
Tonti Stefano
Department of Agricultural Sciences, Alma Mater Studiorum University of Bologna, Viale Giuseppe Fanin 40, 40127, Bologna, Italy
Amato Barbara
Department of Agricultural Sciences, Alma Mater Studiorum University of Bologna, Viale Giuseppe Fanin 40, 40127, Bologna, Italy
Nipoti Paola
Department of Agricultural Sciences, Alma Mater Studiorum University of Bologna, Viale Giuseppe Fanin 40, 40127, Bologna, Italy
Pisi Annamaria
Department of Agricultural Sciences, Alma Mater Studiorum University of Bologna, Viale Giuseppe Fanin 40, 40127, Bologna, Italy
Prodi Antonio
Department of Agricultural Sciences, Alma Mater Studiorum University of Bologna, Viale Giuseppe Fanin 40, 40127, Bologna, Italy
Plant Pathology Journal
Year: 2015 | Volume: 14 | Issue: 2 | Page No.: 86-91







ABSTRACT
Wheat is one of the most important crops in Syria, for both local consumption and export commodity and can be infected by Fusarium Head Blight (FHB) a world-widespread disease. Fusarium Head Blight reduces yield, grain quality and causes accumulation of deoxynivalenol (DON). The most promising and effective management strategy is to avail cultivars resistant to FHB. Ten Syrian durum wheat cultivars and "Simeto", one of the most susceptible Italian cultivars, were artificially infected, under growth chamber and field conditions. They were inoculated with Syrian and Italian strains of Fusarium culmorum, one of the main agents of FHB, to screen their tolerance toward FHB infection, FHB spread, kernel infection and mycotoxin levels. Jory was the most tolerant cultivar in the growth chamber and field, while Sham 9, was the most susceptible.
PDF Abstract XML References Citation
Received: April 18, 2015;
Accepted: May 19, 2015;
Published: June 11, 2015
How to cite this article
Alkadri Dima, Tonti Stefano, Amato Barbara, Nipoti Paola, Pisi Annamaria and Prodi Antonio, 2015. Assessment of Different Resistance Types of Syrian Durum Wheat Cultivars Towards FHB Agent. Plant Pathology Journal, 14: 86-91.
DOI: 10.3923/ppj.2015.86.91
URL: https://scialert.net/abstract/?doi=ppj.2015.86.91
DOI: 10.3923/ppj.2015.86.91
URL: https://scialert.net/abstract/?doi=ppj.2015.86.91
INTRODUCTION
Fusarium Head Blight (FHB) or scab is a serious global cereal disease caused by several Fusarium species, mainly Fusarium culmorum (W.G. Sm.) Sacc. and Fusarium graminearum Schwabe [teleomorph Gibberella zeae (Schwein) Petch]. Fusarium Head Blight (FHB) of wheat causes great yield losses (Brennan et al., 2005), bleached and shrunken kernels, decreasing baking quality due to destruction of starch, proteins and cell walls of the infected kernels (Shinha and Bhatnagar, 1998). Some Fusarium species, besides colonization of host tissues, held to a most serious human and animal health risk for mycotoxin accumulation in food and feed (Pestka, 2010). Mycotoxins are fungal poisonous chemical compounds and are also involved in the inhibition of host resistance reactions (Maier et al., 2006). The most important mycotoxins associated with FHB are deoxynivalenol (DON) and nivalenol (NIV), trichothecenes type (A) and zearalenone (Muller et al., 2012).
Control strategies are difficult and expensive. Breeding for resistance, besides different control practices (Xu and Nicholson, 2009), has been lately worldwide considered of high priority since it seems to be the most promising (Browne and Cooke, 2005) and the most cost-effective tool to control the disease (Buerstmayr et al., 2000). Up to now no wheat cultivars are immune to FHB agents, most of them are susceptible and only a few are moderately resistant (Parry et al., 1995; Cai et al., 2005). Triticum durum L. is more susceptible than Triticum aestivum L. (common wheat) and its grains are exposed to a higher mycotoxin concentration (Stack et al., 2002).
Mesterhazy (1995) found that wheat cultivars have very similar resistance reactions against F. graminearum and F. culmorum. Resistance of wheat to FHB is a complex phenomenon; It is a quantitative character and different components of physiological resistance should be considered: (1) Resistance to initial infection, (2) Resistance to spreading, (3) Resistance to kernel infection, (4) Tolerance to infection and (5) Resistance to DON accumulation (Mesterhazy, 2002).
Wheat (durum and common) is one of the most important crops in Syria, for local consumption and export commodity (NAPC., 2006). The cultivation area is divided into five sub-regions, according to agro-meteorological conditions (FAO., 2003).
There are no reports about the presence of FHB in Syria, but Fusarium species are present and frequently isolated in Syrian wheat kernel samples (Alkadri et al., 2013). The main species isolated were F. culmorum and in less quantity, F. graminearum (Alkadri et al., 2013).
Studies on the FHB resistance of Syrian cultivars are, to our knowledge, very limited; therefore, in this research for the first time, a screening over Syrian durum wheat cultivars in a multi approach experiment (field and growth chamber) has been conducted.
The aim of the study was to investigate varietal differences in ten Syrian durum wheat cultivars, after inoculation with Syrian and Italian F. culmorum strains, in comparison with an Italian susceptible variety. Fusarium Head Blight (FHB) disease severity, kernel infection and mycotoxin accumulation, are used to define Mesterhazy’s resistance types.
MATERIALS AND METHODS
Plant materials and fungal strains: Ten Syrian durum wheat cultivars, the most cultivated in different Syrian areas, were kindly offered by Arab Center for the Study of Arid zones and Dry lands (ACSAD-Syria) to set up experimental trials in the field and growth chamber. The susceptible Italian cultivar, "Simeto", was used as positive control (Table 1).
Six F. culmorum strains (F960, F961 and F966 from Syria and F11, F24 and F35 from Italy), isolated from durum wheat kernels, characterized in 3-ADON chemotype for the presence of gene Tri 12 (Alkadri et al., 2013; Prodi et al., 2011) and for the aggressiveness (Alkadri, 2012) in the laboratory of Phytopathological Mycology-Department of Agricultural Sciences-University of Bologna (Italy), were used for the artificial inocula.
Macroconidia production: Each F. culmorum strain was cultured on potato dextrose agar (PDA, Difco) plates for 7 days. Two mycelium plugs were cut from each strain, were submerged into flasks with autoclaved V8 broth (Singleton et al., 1992) and placed in a refrigerated horizontal type shaker at 140 rpm, 25°C under incident sun light for 2 weeks. The mixture of macroconidia with mycelium in the V8 medium was filtered through a sterile syringe with double layers of autoclaved cheesecloth. Macroconidia suspension was adjusted for each strain to 1×104 mL-1 conidia for floret inoculations in the growth chamber and to 2×105 mL-1 conidia for ear inoculations in the field trials (Purahong et al., 2012).
Floret inoculations in growth chamber: Seeds of the eleven durum wheat cultivars were seeded in trays. After 15 days, each seedling was transplanted into a pot and placed in a growth chamber at 25/19°C day/night temperature, 14/10 h light/dark cycle. One week after transplantation, approximately 3 g of commercial fertilizer (N/P/K) were applied to each plant. The plants were watered three times a week until harvest to avoid water stress conditions. At anthesis (BBCH 63), 20 μL of suspension, at a concentration of 1Χ104 mL-1 conidia, for each strain were injected into two florets (10 μL per floret) at the middle of each spike, between lemma and palea without wounding the ovary. Spikes were then covered with polyethylene bags for 48 h to ensure constant high humidity. For each isolate eight spikes were inoculated. Each pot was considered as a replication and disposed in a completely randomized design. The florets of the control spikes were injected with water. In total, for each cultivar, 56 spikes were evaluated. Diseased-Head Severity (DHS) evaluation was carried out at 7, 14 and 21 days after inoculation (DAI) and based on the percentage of infected area on individual head with a modified Parry’s scale (Purahong et al., 2012): 0% (no symptoms), 2, 5, 10, 25, 50, 75 and 90% (90% or more of bleached area). Diseased-Head Severity is defined as the average proportion of diseased spikelets per diseased spike (sum of the proportion of diseased spikelets per diseased spike divided by the total number of diseased spikes sampled).
The average value of DHS caused by all F. culmorum strains is considered, for each cultivar, a parameter to determine type I resistance. Mean severity (average of DHS values of 1st, 2nd and 3rd evaluations), terminal severity (the 3rd DHS evaluation) and disease development (the means of each evaluation; 7, 14 and 21 DAI-plotted over the estimation time), were used to determine type II resistance for each cultivar.
Koch’s postulate was fulfilled by the re-isolation of F. culmorum from the infected spikes.
Ear inoculations in the field: The eleven durum wheat cultivars were sown in the field of the experimental farm of the University of Bologna (Cadriano, 44°33'4.15"N; 11°24'39.02"E) in autumn 2010-2011. The field was subdivided into 44 micro-plots at double rows (1 m length, 15 cm between the row and 20 cm between micro-plots). Two hundred seeds were sown in each micro-plot for each cultivar (100 per row).
Two mixtures of fungal suspensions: the first was a mixture of the Syrian F. culmorum strains (F960, F961 and F966) and the second of the Italian strains (F11, F24 and F35), at concentration 2×105 mL-1 macroconidia, were prepared. At 30% anthesis (BBCH 63), 60 mL of each conidial suspension were sprayed on each micro-plot using a hand sprayer. The experimental field contained 44 micro-plots disposed in a completely randomized design; 22 inoculated with Syrian strain mixtures and 22 inoculated with Italian ones. The experiment was repeated twice on the 11 cultivars. Natural rain on the inoculation day ensured high humidity, so no addition irrigation was applied.
Five groups of 10 spikes per micro-plot were chosen randomly and marked with plastic labels for disease assessment. Diseased-Head Severity (DHS) and Disease Incidence (DI) at 14 and 21 Days After Inoculation (DAI) were evaluated. The DHS was determined as previously described for the growth chamber. The DI was calculated as the proportion of diseased spikes (number of infected spikes divided by the total number of spikes sampled) (50 spikes per replicate). Mean FHB index was calculated as the product of DI and DHS divided by 100.
The Hectoliter Weight (HW) was measured after the harvest at BBCH 99. Fusarium Damage Kernels (FDK) was determined on one hundred seeds for each replicate, estimating visually the number of scabby "tombstone" infected kernels and recorded as percentage of FDK (Mesterhazy et al., 1999). Koch’s postulate was fulfilled by the re-isolation of F. culmorum from the infected kernels.
DON analysis: Wheat grains obtained from the field experiment were ground and DON was estimated by AgraQuant DON Kit (Romer Labs, Austria), an enzyme immunoassay for the quantitative analysis of DON in cereals. The analysis were set up following the condition indicated by manufacturer; starting from 20 g of each sample shaked into flasks of 300 mL containing 100 mL double distilled water in a rotary shaker (200 rpm) for 3 min. Two replicates for each cultivar inoculated with each mixture were analyzed and DON content was calculated using a microtiter plate spectrometer (OpsysMR, Dynex technologies) and a software package distributed by the manufacturer.
Statistical analysis: Data analysis was performed using SPSS (SPSS Inc. Chicago, IL, v17, 1993-2007). The correlation coefficients among different variables were determined using the Pearson product-moment correlation at a significant level of 5%. ANOVA incorporating the post hoc "Tukey" test at the 5% level of significance, was used to differentiate the means.
RESULTS
Floret inoculations in growth chamber: Typical FHB symptoms were observed in the inoculated spikelets while, no symptoms were present in the control.
The values of DHS evaluations (%) at 7-14-21 DAI for all the cultivars are shown in Fig. 1, cv Jory had the lowest DHS values: DHS-7DAI = 6.8, DHS-14DAI = 17.2, DHS-21 DAI = 28.5.
Based on the terminal severity assessment at 21 DAI, Jory and ACSAD1333 were the most tolerant cultivars (28.6 and 35%, respectively), while all the other Syrian cultivars, except Horani (50.8%), were more susceptible than Simeto (56.8%) (Table 1).
The difference among cultivars, based on the mean diseased head severity, was not significant.
Disease development, statistically significant, ranged between 10.87 for Jory and 30.8 for Sham 5 (Table 1).
Ear inoculations in the field and DON analysis: The inoculated cultivars either with Syrian or Italian inoculum showed the same behaviour (sig (2-tailed) = 0.393>0.05 using independent t-test).
The mean values of FHB index at 14 DAI ranged, for all the different cultivars, from 1.72-7.36% and at 21 DAI from 21.98-40.81 (Table 2).
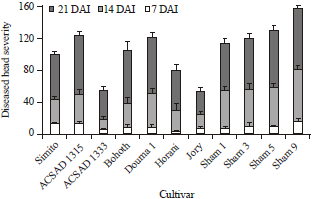 | |
| Fig. 1: | Diseased-head severity evaluations (7, 14 and 21 DAI), in growth chamber for the eleven durum wheat cultivars tested, Error bars represent the standard error of mean |
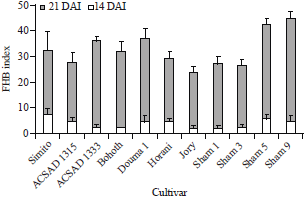 | |
| Fig. 2: | FHB index evaluation (14 and 21 DAI) in %, in the field for the eleven durum wheat cultivars tested, Error bars represent the standard error of mean |
| Table 1: | Values of Diseased-Head Severity (DHS) and disease development in growth chamber for the eleven durum wheat cultivars tested |
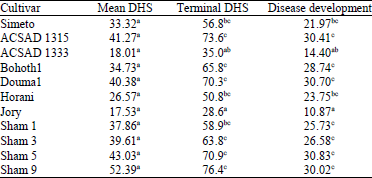 | |
| *Means with the same letter within a column are not significantly different at p = 0.05 according to Tukey test | |
Jory (21.98%) and ACSAD1333 (23.50%) were the most tolerant cultivars, showing significant difference with the most susceptible cultivar Sham 9 (40.81%). The disease development (type II) among all the cultivars was different using ANOVA test; Simeto, Jory, Horani, ACSAD1333 showed a low variability (17.91, 19.91, 20.31 and 20.97, respectively), while Sham 9 had the highest value (36.16) (Table 2 and Fig. 2).
| Table 2: | Field trial: values of FHB index (%) in the 1st and 2nd evaluations after inoculation with Syrian and Italian F. culmorum strains (mix) in the eleven durum wheat cultivars tested, Disease development (%), Fusarium-Damaged Kernels (FDK) in %, Hectoliter Weight (HW) in kg hL-1 and DON level in ppm |
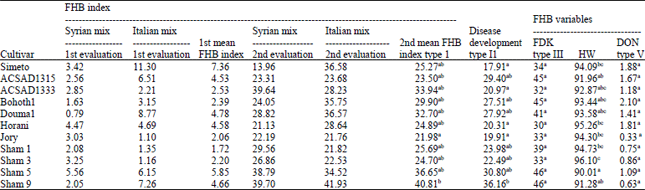 | |
| *Means with the same letter within a column are not significantly different at p = 0.05 according to Tukey test | |
| Table 3: | Correlations among different variables; in the eleven durum wheat cultivars evaluated for resistance to FHB in the field and growth chamber |
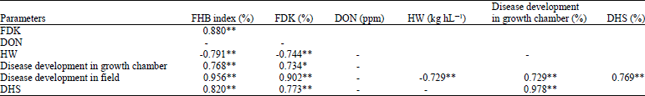 | |
| *,**: Significant at p< 0.05 and p< 0.01, respectively. HW: Hectoliter weight, DHS: Disease head severity, FHB: Fusarium head blight, FDK: Fusarium-damaged kernels and DON: Deoxynivalenol | |
There were no significant differences among all the different cultivars for FDK (type III) and DON levels (type V), however FDK ranged from 30-46% whilst DON from 0.33-2.1 ppm (Table 2).
HW showed a significant diversity among the cultivars; the highest value was for Sham 3 (96.1 kg m-3) while Sham 5 had the lowest (90.01 kg m-3) (Table 2).
Comparison between the results of growth chamber and field trials: The correlations among the different variables in the growth chamber and field are reported in Table 3.
Mean FHB index showed high positive correlation with FDK and negative correlation with HW (r = 0.88 and -0.791, p<0.01, respectively). Furthermore, high correlation was found between DHS and FHB index (r = 0.820, p<0.01). Disease development in the field was correlated with disease development in the growth chamber (r = 0.729, p<0.01). Deoxynivalenol (DON) levels did not show a correlation with any variable.
DISCUSSION
The present study provides preliminary data on FHB infection in most of Syrian cultivated durum wheat cultivars tested in growth chamber and in the Italian experimental field using Syrian and Italian F. culmorum strains.
The data obtained show that all the cultivars differed in their behaviour within the same variable but they were similar, when compared under growth chamber and field conditions, in fact the most tolerant and the most susceptible cultivars kept FHB scores. Furthermore, the terminal DHS in the growth chamber was higher than FHB index in the field. This might be attributed to the fact that humidity and temperature in the growth chamber were adjusted to be ideal for disease development, while the conditions in the field were not controlled. Resistance to initial infection (type I) was strongly influenced by environmental conditions, while the spread of the pathogen within the spike (type II) was related to cultivar resistance (Wisniewska et al., 2004). It is necessary to combine type I and type II resistance to get FHB resistant wheat plants and furthermore, HW could be used to determine the tolerance of cultivars, since it is considered a quality parameter influenced by FHB.
The high correlation between FDK and FHB index is in accordance with Mesterhazy (2002) and Wegulo et al. (2011). According to our results, there was a significant difference within HW among the cultivars and a correlation between HW and FHB index. Ramirez-Marchand et al. (2003) also reported a correlation between FHB and HW.
The DON levels were not significantly different and were not correlated to other variables which imply that cultivars with resistance to FHB do not necessarily show low DON levels. Moreover, toxin and disease resistance are two different phenomena. This is in accord with Wisniewska et al. (2004) and Chrpova et al. (2007), who reported that some cultivars with FHB symptoms clearly expressed high resistance to DON accumulation. Bai et al. (2001) reported that severe visual symptoms may not always be associated with high DON levels. On the contrary, Perkowski and Chelkowski (1993) and Lemmens et al. (1997) observed a significant correlation between resistance to FHB and DON accumulation in seeds after natural infection. Karlovsky (2011) reported that an increase in resistance against FHB was moderate in wheat expressing DON acetylation activity.
These contradictions could be interpreted as the mechanisms of DON accumulation are rather complicated and depend on ecological conditions other than to host and fungal genotypes (Mesterhazy et al., 1999).
Preliminary results indicate that the majority of Syrian cultivars assayed, grown in growth chamber and under Italian field conditions, are susceptible to FHB. Nevertheless, "Jory" showed better tolerance than "Simeto", the most cultivated cultivar in southern Italy, where the climatic conditions are quite similar to some Syrian wheat growing areas. Environmental changes might induce FHB spread in Syrian disease-free areas, being F. culmorum already present in Syrian wheat kernels (Alkadri et al., 2013). Evaluation of a wider range of durum wheat cultivars grown in Syrian regions towards the tolerance/resistance to FHB and DON accumulation would provide more choices and increased benefits to producers and to the food processing industries.
ACKNOWLEDGMENTS
We are grateful to Prof. Ayman Al-Oda from ACSAD for providing the Syrian cultivars and Mr. Stefano Borsari for his collaboration in the field and growth chamber work.
REFERENCES
- Alkadri, D., P. Nipoti, K. Doll, P. Karlovsky, A. Prodi and A. Pisi, 2013. Study of fungal colonization of wheat kernels in Syria with a Focus on Fusarium species. Int. J. Mol. Sci., 14: 5938-5951.
CrossRefDirect Link - Bai, G.H., R. Plattner, A. Desjardins, F. Kolb and R.A. McIntosh, 2001. Resistance to Fusarium head blight and deoxynivalenol accumulation in wheat. Plant Breed., 120: 1-6.
CrossRefDirect Link - Brennan, J.M., D. Egan, B.M. Cooke and F.M. Doohan, 2005. Effect of temperature on head blight of wheat caused by Fusarium culmorum and F. graminearum. Plant Pathol., 54: 156-160.
CrossRefDirect Link - Browne, R.A. and B.M. Cooke, 2005. Resistance of wheat to Fusarium spp. in an in vitro seed germination assay and preliminary investigations into the relationship with Fusarium head blight resistance. Euphytica, 141: 23-32.
CrossRefDirect Link - Buerstmayr, H., B. Steiner, M. Lemmens and P. Ruckenbauer, 2000. Resistance to Fusarium head blight in winter wheat: Heritability and trait associations. Crop Sci., 40: 1012-1018.
CrossRef - Cai, X., P.D. Chen, S.S. Xu, R.E. Oliver and X. Chen, 2005. Utilization of alien genes to enhance Fusarium head blight resistance in wheat: A review. Euphytica, 142: 309-318.
CrossRefDirect Link - Chrpova, J, V. Sip, E. Matejova and S. Sykorova, 2007. Resistance of winter wheat varieties registered in the Czech Republic to mycotoxin accumulation in grain following inoculation with Fusarium culmorum. Czech J. Genet. Plant Breed., 43: 44-52.
Direct Link - Karlovsky, P., 2011. Biological detoxification of the mycotoxin deoxynivalenol and its use in genetically engineered crops and feed additives. Applied Microbiol. Biotechnol., 91: 491-504.
CrossRefPubMedDirect Link - Lemmens, M., R. Josephs, R. Schuhmacher, H. Grausgruber and H. Buerstmayr et al., 1997. Head blight (Fusarium spp.) on wheat: Investigations on the relationship between disease symptoms and mycotoxin content. Cereal Res. Commun., 25: 459-465.
Direct Link - Maier, F.J., T. Miedaner, B. Hadeler, A. Felk and S. Salomon et al., 2006. Involvement of trichothecenes in fusarioses of wheat, barley and maize evaluated by gene disruption of the trichodiene synthase (TRI5) gene in three field isolates of different chemotype and virulence. Mol. Plant Pathol., 7: 449-461.
CrossRefPubMed - Mesterhazy, A., 2002. Role of deoxynivalenol in aggressiveness of Fusarium graminearum and F. culmorum and in resistance to Fusarium head blight. Eur. J. Plant Pathol., 108: 675-684.
CrossRefDirect Link - Mesterhazy, A., 1995. Types and components of resistance to Fusarium head blight of wheat. Plant Breed., 114: 377-386.
CrossRefDirect Link - Mesterhazy, A., T. Bartok, C.G. Mirocha and R. Komoroczy, 1999. Nature of wheat resistance to Fusarium head blight and the role of deoxynivalenol for breeding. Plant Breed., 118: 97-110.
CrossRefDirect Link - Parry, D.W., P. Jenkinson and L. McLeod, 1995. Fusarium ear blight (Scab) in small grain cereals-A review. Plant Pathol., 44: 207-238.
CrossRefDirect Link - Pestka, J.J., 2010. Toxicological mechanisms and potential health effects of deoxynivalenol and nivalenol. World Mycotoxin J., 3: 323-347.
Direct Link - Prodi, A., S. Tonti, D. Salomoni, I. Alberti, P. Nipoti, A. Pisi and D. Pancaldi, 2011. Fusarium graminearumm and F. Culmorum chemotypes as potential producers of mycotoxins in durum wheat, grown in some Italian regions. Proceedings of the 7th International Congress of Food Technologists, Biotechnologists and Nutritionists-Opatjia, September 21-23, 2011, Croatia, Europe, pp: 328-333.
- Purahong, W., D. Alkadri, P. Nipoti, A. Pisi, M. Lemmens and A. Prodi, 2012. Validation of a modified Petri-dish test to quantify aggressiveness of Fusarium graminearum in durum wheat. Eur. J. Plant Pathol., 132: 381-391.
CrossRefDirect Link - Ramirez-Marchand, C., C. Mendoza-Zamora, S. Gilchrist, F. Capettini and N. Marban-Mendoza, 2003. Toxins and damage induced by seven species of Fusarium head blight on barley in the central highlands of Mexico. Proceedings of the National Fusarium Head Blight Forum Proceedings, December 13-15, 2003, Bloomington, Indiana, pp: 165-169.
Direct Link - Wegulo, S.N., W.W. Bockus, J.H. Nopsa, E.D. de Wolf, K.M. Eskridge, K.H.S. Peiris and F.E. Dowell, 2011. Effects of integrating cultivar resistance and fungicide application on Fusarium head blight and deoxynivalenol in winter wheat. Plant Dis., 95: 554-560.
CrossRefDirect Link - Wisniewska, H., J. Perkowski and Z. Kaczmarek, 2004. Scab response and deoxynivalenol accumulation in spring wheat kernels of different geographical origins following inoculation with Fusarium culmorum. J. Phytopathol., 152: 613-621.
CrossRefDirect Link - Xu, X. and P. Nicholson, 2009. Community ecology of fungal pathogens causing wheat head blight. Annu. Rev. Phytopathol., 47: 83-103.
CrossRefDirect Link