
Hancheng Wang
Key Laboratory of Molecular Genetics, Guizhou Academy of Tobacco Sciences, Guiyang, 550081, People�s Republic of China
Yanfei Huang
College of Agriculture, Yangtze University, Jingzhou, 434025, People�s Republic of China
Haiqian Xia
Key Laboratory of Molecular Genetics, Guizhou Academy of Tobacco Sciences, Guiyang, 550081, People�s Republic of China
Jin Wang
College of Life Science, Yangtze University, Jingzhou, 434025, People�s Republic of China
Maosheng Wang
Key Laboratory of Molecular Genetics, Guizhou Academy of Tobacco Sciences, Guiyang, 550081, People�s Republic of China
Changqing Zhang
College of Agriculture, Yangtze University, Jingzhou, 434025, People�s Republic of China
Hongxue Lu
College of Agriculture, Yangtze University, Jingzhou, 434025, People�s Republic of China
Plant Pathology Journal
Year: 2015 | Volume: 14 | Issue: 2 | Page No.: 79-85







ABSTRACT
Tobacco brown spot caused by Alternaria alternata is a devastating disease of tobacco worldwide. Phenotypic characterization of the pathogen was investigated to provide some basic information for biology and pathology by using BIOLOG Phenotype Microarray (PM). Using PM plates 1-10, 950 different growth conditions were tested. Results exhibited that the pathogen was able to metabolize 24.74% of tested carbon sources, 85.26% of nitrogen sources, 97.14% of sulfur sources and 89.83% of phosphorus sources. Most informative utilization patterns for carbon sources of A. alternata were carbohydrates and for nitrogen were various amino acids. The pathogen presented 274 different nitrogen pathways. It had wide range adaptabilities in osmolytes with up to 10% sodium chloride, up to 6% potassium chloride, up to 5% sodium sulfate, up to 20% ethylene glycol, up to 6% sodium formate, up to 6% urea, up to 12% sodium lactate, up to 200 mM sodium phosphate, up to 100 mM ammonium sulfate, up to 100 mM sodium nitrate and up to 20 mM sodium nitrite. It also exhibited active metabolism in the range of pH values between 3.5 and 10, with optimal pH of around 6.0. The pathogen showed active decarboxylase activity, whereas no deaminase activity in the presence of various amino acids.
PDF Abstract XML References Citation
Received: March 31, 2015;
Accepted: May 19, 2015;
Published: June 11, 2015
How to cite this article
Hancheng Wang, Yanfei Huang, Haiqian Xia, Jin Wang, Maosheng Wang, Changqing Zhang and Hongxue Lu, 2015. Phenotypic Analysis of Alternaria alternata, the Causal Agent of Tobacco Brown Spot. Plant Pathology Journal, 14: 79-85.
DOI: 10.3923/ppj.2015.79.85
URL: https://scialert.net/abstract/?doi=ppj.2015.79.85
DOI: 10.3923/ppj.2015.79.85
URL: https://scialert.net/abstract/?doi=ppj.2015.79.85
INTRODUCTION
Tobacco (Nicotiana tabacum L.) is an annual, leafy and solanaceous plant grown commercially for its leaves. China is the biggest single tobacco market in the world and accounts for more than 39.6% of the total global tobacco production (Wang et al., 2013) and 40% of the global tobacco consumption (Wang et al., 2014). During tobacco production period, the major destructive foliar disease tobacco brown spot, occurs every field where tobacco growing throughout the country. Tobacco leaves infected by this disease normally become incomplete, uneven baking leaf color and leaf thickness which results in poor quality of tobacco leaves and low value of industrial use (Jenning et al., 2002; Yakimova et al., 2009). Losses can reach more than 60% if disease management practices are not utilized. In the last five years, it has been a major serious problem for tobacco production in China (Tong et al., 2012). This disease is caused by the notorious fungal pathogen Alternaria alternata (Fr.) Keissl (Main, 1969; Dobhal and Monga, 1991). It normally happens from the lower leaves of a plant and gradually spreads to the upper leaves during leaf harvest period (Stavely and Slana, 1970). The typical symptom of brown spot on leaf is a brown necrotic centre surrounded by a yellow or yellowish-green halo (Slavov et al., 2004).
During the early part of the growing season, A. alternata grows extensively over the leaf surface and only seldom penetrates tobacco leaf for invasion. Whereas at harvest period, the pathogen directly penetrates the cuticle after formation of an appressorium or infects leaves from stomata (Huang et al., 1996); later on, necrotic lesions are formed after host cells have collapsed due to fungal action (Main, 1969; Cheng et al., 2011). To the best of our knowledge, the mechanism of infection differences in two stages is still unclear. There might be some different chemicals on tobacco leaf interrelated with the invasion for the different growing stages and these chemicals could influence the metabolic activity of A. alternata. Up until now, little is known about the metabolic characters of the isolate of A. alternata. A better understanding of the phenotypes of the pathogen will be much valuable to develop management practices to decrease the impacts of the disease.
Recently, a high throughput Phenotypic Microarray (PM)/ Omnilog system, was developed by the company Biolog (Hayward, CA) to assay nearly 1000 metabolic phenotypes (Bochner et al., 2001). In the system, microbial cells in defined medium containing a reducible tetrazolium dye are inoculated into ten 96-well plates, where the plates contain various substrates to assay for: the utilization of carbon, nitrogen, sulfur, phosphorus source, biosynthetic pathways and varying effects of osmotic, ionic and pH environments. Cell growth and respiration lead to the reduction of the tetrazolium dye and result in blue color. The intensity of the color which is proportional to microbial growth, is recorded every 15 min by a CCD camera and analyzed by Omnilog software. The software plots the kinetic data of color formation in arbitrary units against time for each well and also assigns artificial color to each plot.
To address these questions, we aim to elucidate phenotype analysis of A. alternata, a species that is emerging as a model for the studies of fungal biology and pathology (Velez et al., 2007; Jia et al., 2010; Llorens et al., 2013). The outcome would provide useful information to know detail biology of A. alternata and helpful knowledge for brown spot management.
MATERIALS AND METHODS
Fungal strain and culture conditions: One isolate of A. alternata, with wild-type sensitivity and pathogenicity to tobacco, was selected as the modal species for analysis. The isolate was grown and maintained on AEA agar (yeast power 5 g L-1, NaNO3 6 g L-1, KH2PO4 1.5 g L-1, KCl 0.5 g L-1, MgSO4 0.25 g L-1, glycerol 20 mL L-1 and 16.0 g L-1 agar) (Li et al., 2005; Jin et al., 2007), in a controlled climate cabinet at 25°C in darkness. After 7 days’ incubation, conidia produced on the Petri dishes were washed with distilled water, the suspension was filtered through a double-layer of sterile cheesecloth (Grade #40: 24×20 threads per inch) to remove mycelia fragments and the resulting conidia suspension was diluted to a final concentration of 1×105 spores mL-1.
Phenotypic characterization: The metabolic abilities of A. alternata were tested by using the Phenotype Microarray (PM) system (Biolog, Hayward, CA, USA) according to the published procedure (Bochner et al., 2001; Zhou et al., 2003; Von Eiff et al., 2006). Using PM system, 950 different growth conditions were tested, including 190 different carbon sources, 95 nitrogen sources, 59 phosphorus sources, 35 sulfur sources, 94 biosynthetic pathways, 285 nitrogen pathways and 192 tolerances to different osmolytes and pH conditions. All materials, media and reagents for the PM system were purchased from Biolog. Plates 1-8 which test for catabolic pathways for carbon, nitrogen phosphorus, sulfur, as well as for biosynthetic pathways and plates 9-10 which test for osmotic/ion and pH effects, were utilized in this study. Conidial suspension of A. alternata was prepared as mentioned above and suspended in appropriate medium containing sterile FF-IF; 100 μL of a dilution of an 62% transmittance suspension of cells were added to each well of the PM plates. FF-IF was used for PM plates 1 and 2. FF-IF plus 100 mM D-glucose, 5 mM potassium phosphate (pH 6.0) and 2 mM sodium sulfate was used for plates 3, 5, 6, 7 and 8. FF-IF plus 100 mM D-glucose was used for plate 4. FF-IF plus yeast nitrogen base and 100 mM D-glucose was used for plates 9 and 10. Plates were incubated in the OmniLog at 28°C for 7 days with readings taken every 15 min. Incubation and recording of phenotypic data were performed in the OmniLog station by capturing digital images of the microarrays and storing turbidity values in a computer file displayed as a kinetic graph. Data analysis was conducted using Kinetic and Parametric software (Biolog). Phenotypes were determined based on the area under the kinetic curve of dye formation. The experiments were conducted twice.
RESULTS
Phenotypic characterization: Isolate of A. alternata tested in our study presented typical phenotypic fingerprint. The pathogen was able to metabolize 24.74% of tested carbon sources (26/95 in plate PM1 and 21/95 in plate PM2), 85.26% of nitrogen sources (67/95 in plate PM3, 95/95 in plate PM6, 80/95 in plate PM7 and 82/95 in plate PM8), 97.14% of sulfur sources (34/35 in plate PM4, Wells F02-H12) and 89.83% of phosphorus sources (53/59 in plate PM4, Wells A02-E12) (Fig. 1). The pathogen presented apparent growth in the negative control without any phosphorus source (plate PM4, Well A1) or any sulfur compound (plate PM4, Well F01). The phosphorus compounds that were effectively utilized by A. alternata included D-2-phospho-glyceric acid, 2-deoxy-D-glucose 6-phosphate, D-mannose-1-phosphate, O-phospho-D-tyrosine and O-phospho-L-tyrosine (Table 1, Fig. 1). Meanwhile, nearly all S-containing compounds (35/35 tested, plate PM4, Wells F1-H12) tested could be assimilated by the pathogen, except for D, L-Ethionine (plate PM4, Well G06) (Fig. 1), while three sulfur compounds were effectively utilized by the pathogen, including dithiophosphate (plate PM4, Well F06), L-cysteine (plate PM4, Well F07) and butane sulfonic acid (plate PM4, Well H09). Simultaneously, the pathogen presented 94 different biosynthetic pathways (94/94 tested, plate PM5, Wells A3-H12).
 | |
| Fig. 1: | Data for biolog phenotype microarray PM 1-10 plates of the pathogen Alternaria alternata, (Utilization of the isolates of A. alternata from tobacco was indicated by green areas in the growth curve for each substrate) |
| Table 1: | Substrates in PM 1-10 Micro plates significantly supported the growth of Alternaria alternata |
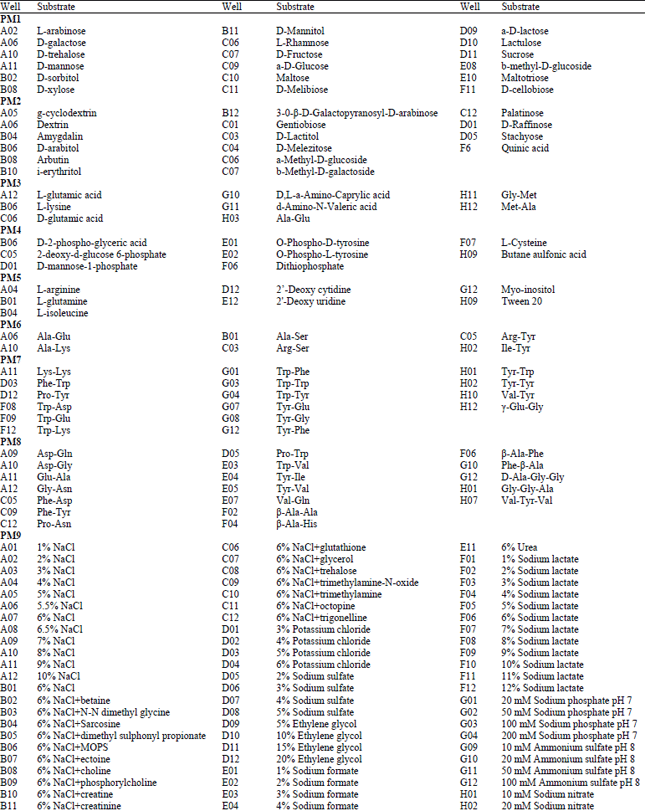 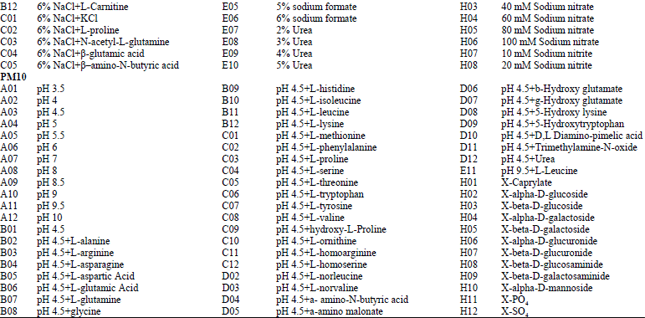 | |
Using data from PM1 and PM2 (carbon sources), isolates of A. alternata from tobacco were able to use 103 different carbon sources (Fig. 1), about 34 compounds significantly supported the growth of the pathogen (Table 1), including L-arabinose, D-galactose, D-trehalose, D-mannose, D-sorbitol, D-xylose, D-mannitol, L-rhamnose, D-fructose, a-D-glucose, maltose, D-melibiose, a-D-lactose, lactulose, sucrose, b-methyl-D-glucoside, maltotriose, D-cellobiose, g-cyclodextrin, dextrin, amygdalin, D-arabitol, arbutin, i-erythritol, 3-0-β-D-galactopyranosyl-D-arabinose, gentiobiose, D-lactitol, D-melezitose, a-methyl-D-glucoside, b-methyl-D-galactoside, palatinose, D-raffinose, stachyose and quinic acid. In comparison, around 85 compounds significantly inhibited the growth of the pathogen (Fig. 1). Using the PM3 plate, the isolate of A. alternata was tested for their ability to grow on 95 different nitrogen sources (amino acids) (Fig. 1). More than 65 compounds supported the growth of the pathogen (Table 1), typical compounds included L-glutamic acid, L-lysine, D-glutamic acid, D,L-a-amino-caprylic acid, d-amino-N-valeric acid, Ala-Glu, Gly-Met and Met-Ala. In comparison, twenty-four out of 95 nitrogen sources supported growth at the level of the negative control, indicating that A. alternata cannot metabolize these compounds (Fig. 1).
Using data from PM6 to PM8 (nitrogen pathways), A. alternata presented 274 different nitrogen pathways, indicating that various combinations of different amino acids supported the growth of the pathogen (Fig. 1). Plates PM9 and PM10 were used to test growth under various stress conditions. A. alternata showed active metabolism with up to 10% sodium chloride, up to 6% potassium chloride, up to 5% sodium sulfate, up to 20% ethylene glycol, up to 6% sodium formate, up to 6% urea, up to 12% sodium lactate, up to 200 mM sodium phosphate (pH 7.0), up to 100 mM ammonium sulfate (pH 8.0), up to 100 mM sodium nitrate and up to 20 mM sodium nitrite, however, it could not metabolize any sodium benzoate (pH 5.2) which ranged from 20-200 mM (plate PM 9, Well G05, G06, G07 and G08) in our analysis (Fig. 1). When combined with various osmolytes at the treatment of 6% sodium chloride, A. alternata presented active growth in all tests (plate PM9, Well B01 to B12 and C01 to C12). The pH range where A. alternata exhibited active growth was between 3.5 and 10, with an optimal pH of around 6.0. When combined with various amino acids at the pH of 4.5, A. alternata showed active growth in all tests except for combining with the amino acids of anthranilic acid (plate PM 10, well D01) and p-aminobenzoate (plate PM 10, well D05) (Fig. 1, PM10). In comparison, when combined with various amino acids at the pH of 9.5, the pathogen presented no growth in all tests. PM 10, wells B1-D12 and E1-G12, tested the decarboxylase and deaminase activities of the pathogen in the presence of amino acids at pH 4.5 and pH 9.5, respectively. A. alternata showed active decarboxylase activities in the presence of most of the amino acids, except for anthranilic acid (plate PM 10, Well D1) and p-aminobenzoate (plate PM 10, Well D05); in comparison, it exhibited no deaminase activities (Fig. 1, plate PM 10), except for L-leucine (plate PM 10, Well E11).
DISCUSSION
While a large number of genetic and molecular biological studies have been conducted with A. alternata, phenotypic diversity is still poorly explored. Direct high-throughput assessment of phenotypes using the Phenotype MicroArray (PM) system (Bochner et al., 2001) has stirred much attention for molecular biological, genomic and population studies of microorganisms (Velez et al., 2007). Here, metabolic ability of the pathogen from host of flue-cured tobacco was systematically analyzed using PMs and significant metabolic diversity was found.
In this study, a pathogenic isolate of A. alternata from the host of flue-cure tobacco was analyzed by the high-throughput PM technique. A narrow range of carbon compounds could be utilized by the pathogen and most of the nitrogen, sulfur and phosphorus sources were metabolized. Since, carbon substrate is the most important nutrient in the life of most microorganisms, the narrow range of carbon utilization of A. alternata may indicate a poor adaptability of the pathogen in tobacco developing period. Early studies have proved that tobacco leaves are seldom infected by A. alternata during the early part of the growing season, whereas at harvest time all plant parts above ground become heavily infected (Ramm and Lucas, 1963; Stavely and Slana, 1970; Tong et al., 2012). There might be some carbon substrates produced on tobacco leaves only at harvest time to induce the infection of the pathogen. The narrow carbon substrates metabolized in our study may in some case explain the infection mechanism of A. alternata on tobacco. However, whether those carbon substrates are presence in the mature or near mature tobacco leaves or not is still unclear, more work is need to be conducted to confirm this hypothesis.
The most informative plates for A. alternata were PM1/PM2 (carbon sources), PM3 (nitrogen sources), PM 9 (osmolytes conditions) and PM 10 (pH conditions). Most informative utilization patterns for carbon sources were just some carbohydrates and for nitrogen sources some amino acids. There were some carbohydrates reported in tobacco leaf that regulated leaf development (Miller et al., 2000), stimulated ethylene production (Philosoph-Hadas et al., 1985), metabolized just during leaf ageing progress (Crafts-Brandner, 1991) and that existed just on the leaf surface of tobacco (Menetrez et al., 1990). Such compounds might play a role in supporting the infection of tobacco leaves by A. alternata. Additionally, the pathogen had wide range adaptabilities in osmolytes and pH conditions which was found by using plates PM 9 and PM 10. The phenotypic diversity in osmolytes of the pathogen can be rationalized by considering the seasonal variation in osmolytes due to varying conditions of dryness and watering. Decarboxylase of the pathogen generates alkaline amines by the catabolism of amino acids which help to counteract an acidic pH (Maurer et al., 2005). A high pH on the other hand can be countered by deaminases which generate acids. A. alternata in our study showed active decarboxylase activities, whereas no deaminase activities. Hence, strong alkalic chemicals might have some potential ability to inhibit the metabolism of the pathogen and could be used for tobacco brown spot management. Consequently, phenotypic characters for utilization of those sources and wide range adaptabilities of A. alternata could have a high potential value in pathogen-tobacco interaction studies and survival of the pathogen in environment.
Since, the PM technique examines strains for many characters relating to how environmental stressors affected pathogen’s activity, it becomes possible to determine whether it has a reasonable chance of being useful for disease management in agriculture. Enhancing the amount of carbon and nitrogen sources that could not be metabolized by A. alternata, or decreasing the amount of these sources that could be utilized by the pathogen in the field may reduce the damage caused by brown spot. Meanwhile, changing the osmolytes and pH environment in tobacco leaves to make it unadaptable for the pathogen A. alternata may also lighten the happening of brown spot. These imagines could be proved in the next study in the near future.
CONCLUSION
In conclusion, phenomics study of A. alternata in our study increases our understanding of the pathogen.
ACKNOWLEDGMENTS
This study was supported by National Natural Science Foundation of China (31360448), Guizhou Tobacco Company Program (201305, 2013036) and key laboratory of Tobacco molecular Genetics and Qianxi Tobacco Company program (2013-06). The authors appreciate the anonymous reviewers for critical reviews of the manuscript.
REFERENCES
- Bochner, B.R., P. Gadzinski and E. Panomitros, 2001. Phenotype microarrays for high-throughput phenotypic testing and assay of gene function. Genome Res., 11: 1246-1255.
Direct Link - Cheng, D.D., Y.J. Jia, H.Y. Gao, L.T. Zhang, Z.S. Zhang, Z.C. Xue and Q.W. Meng, 2011. Characterization of the programmed cell death induced by metabolic products of Alternaria alternata in tobacco BY-2 cells. Phsiol. Plant., 141: 117-129.
CrossRefDirect Link - Huang, S.L., K. Kohmoto, H. Otani and M. Kodama, 1996. Nuclear behavior during the formation of appressoria by Alternaria alternata. Mycoscience, 37: 41-47.
CrossRefDirect Link - Jenning, D.B., M.E. Daub, D.M. Pharr and J.D. Williamson, 2002. Constitutive expression of a celery mannitol dehydrogenase in tobacco enhances resistance to the mannitol-secreting fungal pathogen Alternaria alternata. Plant J., 32: 41-49.
CrossRefDirect Link - Jia, Y.J., D.D. Cheng, W.B. Wang, H.Y. Gao, A.X. Liu, X.M. Li and Q.W. Meng, 2010. Different enhancement of senescence induced by metabolic products of Alternaria alternata in tobacco leaves of different ages. Physiol. Plant., 138: 164-175.
CrossRefDirect Link - Jin, L.H., C.J. Chen, J.X. Wang, Y. Chen and M.G. Zhou, 2007. Activity of azoxystrobin and SHAM of four plant pathogens. Sci. Agric. Sin., 40: 2206-2213.
Direct Link - Llorens, E., E. Fernandez-Crespo, B. Vicedo, L. Lapena and P. Garcia-Agustin, 2013. Enhancement of the citrus immune system provides effective resistance against Alternaria brown spot disease. J. Plant Physiol., 170: 146-154.
CrossRef - Maurer, L.M., E. Yohannes, S.S. Bondurant, M. Radmacher and J.L. Slonczewski, 2005. pH regulates genes for flagellar motility, catabolism and oxidative stress in Escherichia coli K-12. J. Bacteriol., 187: 304-319.
CrossRefDirect Link - Menetrez, M.L., H.W. Spurr Jr., D.A. Danehower and D.R. Lawson, 1990. Influence of tobacco leaf surface chemicals on germination of Peronospora tabacina adam sporangia. J. Chem. Ecol., 16: 1565-1576.
CrossRefDirect Link - Miller, A., C. Schlagnhaufer, M. Spalding and S. Rodermel, 2000. Carbohydrate regulation of leaf development: Prolongation of leaf senescence in Rubisco antisense mutants of tobacco. Photosynth. Res., 63: 1-8.
CrossRefDirect Link - Philosoph-Hadas, P.S., S. Meir and N. Aharoni, 1985. Carbohydrates stimulate ethylene production in tobacco leaf discs II. Sites of stimulation in the ethylene biosynthesis pathway. Plant Physiol., 78: 139-143.
Direct Link - Slavov, S., S. Mayama and A. Atanassov, 2004. Toxin production of Alternaria alternata tobacco pathotype. Biotechnol. Biotechnol. Equip., 18: 90-95.
CrossRefDirect Link - Stavely, J.R. and L.J. Slana, 1970. Relation of leaf age to the reaction of tobacco to Alternaria alternata. Phytopathology, 61: 73-78.
Direct Link - Crafts-Brandner, S.J., 1991. Nonstructural carbohydrate metabolism during leaf ageing in tobacco (Nicotiana tabacum). Physiol. Plant., 82: 299-305.
CrossRefDirect Link - Tong, Z., T. Jiao, F. Wang, M. Li and X. Leng et al., 2012. Mapping of quantitative trait loci conferring resistance to brown spot in flue-cured tobacco (Nicotiana tabacum L.). Plant Breed., 131: 335-339.
CrossRefDirect Link - Velez, H., N.J. Glassbrook and M.E. Daub, 2007. Mannitol metabolism in the phytopathogenic fungus Alternaria alternata. Fungal Genet. Biol., 44: 258-268.
CrossRefDirect Link - Von Eiff, C., P. McNamara, K. Becker, D. Bates and X.H. Lei et al., 2006. Phenotype microarray profiling of Staphylococcus aureus menD and hemB mutants with the small-colony-variant phenotype. J. Bacteriol., 188: 687-693.
CrossRefDirect Link - Wang, H.C., J. Wang, W.H. Li, Y.F. Huang and H.Q. Xia et al., 2014. Cladosporium cladosporioides identified in China on tobacco seeds. Plant Dis., 98: 1002-1002.
CrossRef - Wang, H.C., M.S. Wang, H.Q. Xia, S.J. Yang and Y.S. Guo et al., 2013. First report of Fusarium wilt of tobacco caused by Fusarium kyushuense in China. Plant Dis., 97: 424-424.
CrossRefDirect Link - Yakimova, E.T., Z.P. Yordanova, S. Slavov, V.M. Kapchina-Toteva and E.J. Woltering, 2009. Alternaria alternata AT toxin induces programmed cell death in tobacco. J. Phytopathol., 157: 592-601.
CrossRefDirect Link - Zhou, L., X.H. Lei, B.R. Bochner and B.L. Wanner, 2003. Phenotype microarray analysis of Escherichia coli K-12 mutants with deletions of all two-component systems. J. Bacteriol., 185: 4956-4972.
CrossRefDirect Link