
E.A. Gyamera
Department of Nuclear Agriculture and Radiation Processing, Graduate School of Nuclear and Allied Sciences, University of Ghana, P.O. Box AE1, Atomic, Accra, Ghana
H.M. Amoatey
Department of Nuclear Agriculture and Radiation Processing, Graduate School of Nuclear and Allied Sciences, University of Ghana, P.O. Box AE1, Atomic, Accra, Ghana
G.K. Owusu
Department of Nuclear Agriculture and Radiation Processing, Graduate School of Nuclear and Allied Sciences, University of Ghana, P.O. Box AE1, Atomic, Accra, Ghana
Plant Pathology Journal
Year: 2015 | Volume: 14 | Issue: 2 | Page No.: 58-64







ABSTRACT
Mathematical models play crucial role in effective plant disease management. They permit accurate forecasting of plant disease epidemics for timely interventions. However, the use of mathematical models in plant disease management has not received much attention in Ghana. This study assesses the disease intensities and the spatio-temporal spread patterns of viral diseases on a zucchini squash (Cucurbita pepo L.) field in the coastal savannah zone of Ghana. Disease intensity data were transformed by Exponential, Monomolecular, Gompertz and Logistic models. Semivariance analysis and Inverse Distance Weighting interpolations were performed using the GS+geostatistical software. The Gompertz model explained best the observed variability in disease incidence data with 90.86% agreement between field-observed and model-predicted disease incidence data. For disease severity, the Exponential model best described the progress of the disease with a co-efficient of determination of 94.38%. The semivariogram estimated a range of spatial dependence of 0.63 m and a sill of 1.91400.
PDF Abstract XML References Citation
Received: March 22, 2015;
Accepted: May 18, 2015;
Published: June 11, 2015
How to cite this article
E.A. Gyamera, H.M. Amoatey and G.K. Owusu, 2015. Spatial and Temporal Spread Patterns of Viral Diseases on a Zucchini Squash Field in the Coastal Savannah Zone of Ghana. Plant Pathology Journal, 14: 58-64.
DOI: 10.3923/ppj.2015.58.64
URL: https://scialert.net/abstract/?doi=ppj.2015.58.64
DOI: 10.3923/ppj.2015.58.64
URL: https://scialert.net/abstract/?doi=ppj.2015.58.64
INTRODUCTION
The profit oriented nature of commercial agriculture and access to limited economic resources require that plant disease control measures are undertaken only after a specific disease incidence has reached a critical threshold. This reduces production costs to enhance profit margins.
Quantitative knowledge of the various factors that contribute to disease progress is essential in developing cost effective approaches to disease management (Nutter, 2007). Various functions and techniques have been developed for describing disease spread patterns, the foremost being the use of mathematical models (Teng, 1985).
Mathematical models provide the basis and theories essential in quantifying disease progress curves that bring out relations which may otherwise not be obvious from raw disease intensity data (Madden and Campbell, 1990). The progress of an epidemic can be quantified in both time (temporal) and space (spatial) dimensions. There are numerous mathematical models available for quantifying the temporal patterns of disease spread. However, non-linear forms of asymptotic mechanistic models such as exponential, logistic, Weibull, Richards and Gompertz models are generally used in analyzing polycyclic epidemics such as virus pathosystems in cucurbits (Fekedulegn et al., 1999). Spatial Autocorrelations and Inverse Distance Weighting methods also permit the investigation of spatial dependence of disease intensities among neighbouring plant units. This study reports the spatial and temporal spread patterns of viral diseases on a zucchini squash field in the coastal savannah agro-ecological zone of Ghana.
MATERIALS AND METHODS
Experimental design and field layout: The study was conducted at the Biotechnology and Nuclear Agriculture Research Institute (BNARI) of the Ghana Atomic Energy Commission in Ghana. The experimental plot measuring 24 m in length and 20 m in width was divided into two sub-plots of 24×9 m each, with a path of 2 m separating the sub-plots. Each of the two subplots contained approximately 250 zuchini squash (Cucurbita pepo L.) plants. In all, there were approximately 500 zucchini squash plants for monitoring the spread of the cucurbit viruses. All zucchini squash rows within a sub-plot were spaced at a distance of 1.0 m. Disease incidence and severity were assessed on a 10 quadrats basis of which the entire field was partitioned. On the western border of the zucchini squash field was an old cucumber field, the Eastern and Northern borders of the zucchini squash field were cultivated with eggplants but there were weeds as well. The Southern border was largely occupied by weeds. There was no fertilizer or pesticide application throughout the period of the experiment.
Assessment of disease incidence and severity: Each zucchini squash plant was examined for symptom development on weekly basis for a period of seven weeks. Virus-like symptoms were recorded, together with the time of symptom occurrence and the geographical location of the symptomatic plant. Disease Incidence (DI) and Disease Severity Factor (DSF) were calculated, first, for each quadrat and then for all the quadrats by the following relations:
| (1) |
| (2) |
where, a is the severity score for plant 1, 2…n (range: 0-5) and b is the upper limit for disease severity score.
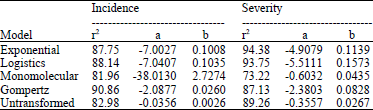
Assessment of the temporal patterns of disease spread: Disease incidence data for all quadrats were analyzed by Microsoft Excel 2007 and Minitab version 15. A scatter plot was generated for each quadrat and then for the average of all quadrats. Disease intensity data were fitted to the linearized forms of the non-linear exponential, logistic, gompertz and monomolecular mechanistic models as shown in Table 1. The model that gave the highest coefficient of determination (r2), was selected as the most appropriate model for describing the temporal pattern of the epidemic in a particular quadrat or the entire field (Madden, 2012). Primary infection and apparent infection (secondary transmission) rates of the disease spread were estimated from the selected models for each quadrat and for the entire field. Tables of estimated parameters from the selected empirical and mechanistic models were generated for all the quadrats and for the entire field.
Assessment of the spatial patterns of disease spread: Semivariogram and inverse distance weighting interpolation (IDW) methods were used in analyzing the spatial patterns of the disease spread across the zucchini squash field.
| Table 1: | Linearized forms of the non-linear mechanistic growth models used in transforming the disease intensity (y) |
 | |
| Source: Madden (1980, 2012) | |
Data for all symptomatic squash plants were weighted by assigning a temporal value to each squash plant which corresponded with the time of symptom appearance on the particular plant in relation to the last day of disease assessment. Plants that became symptomatic earlier in the epidemic received higher weightings compared with plants that became infected around the period of final field disease assessment. Each of the plants in the field was also assigned X and Y coordinate values in a Cartesian X-Y plane based on the specific location of the plant in the field. The 2-dimensional spatial and temporal weighting data were then analyzed with GS+ version 9 Gama Design Software. Parameters such as nugget, sill and range of spatial dependence were estimated based on the selected semivariogram models. Interpolations were performed using the inverse distance weighting method (Roumagnac et al., 2004). A map of the disease field was then generated that showed the estimated disease status of all zucchini squash plants on the field, including all asymptomatic plants.
RESULTS
Disease incidence and severity: Figure 1 shows the average disease intensities observed in all ten quadrats of the zucchini squash field. Disease intensities increased significantly from the third week of disease assessment onwards.
Temporal patterns of disease spread: In terms of disease incidence, all the fitted mechanistic models gave a better fit compared to the untransformed data except data fitted to the monomolecular model. The gompertz model explained best, the observed variability in the incidence data with 90.86% agreement between field-observed and model-predicted disease incidence data. This model estimated the apparent infection rate to be 0.026 day‾1 and predicted the doubling time (tD) with reference to the last day of disease assessment to be 15 days. For disease severity, the exponential model best described the temporal pattern of the disease spread with a high coefficient of determination (r2) of 94.38%. Only the logistic and exponential model transformations described the pattern of disease severity better than the untransformed data. Table 2 shows the parameters estimated from the selected mechanistic models used in fitting disease intensity data.
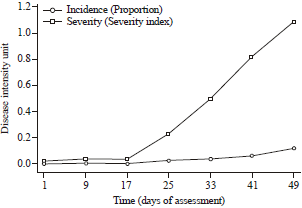 | |
| Fig. 1: | Average disease intensities observed on the entire zucchini squash field |
Figure 2 shows the spread of the model transformed disease incidence data points around the fitted regression functions whilst Fig. 3 shows the scatterplots generated after data from disease severity factors from the zucchini squash field were transformed by exponential, logistic monomolecular and gompertz models.
Spatial patterns of disease spread
Semivariance analyses: An autocorrelation was performed with an angle of 0° North relative to the principal axis at a tolerance limit of 180°, a Gaussian model was fitted to the data with a Residual Sum of Square (RSS) value of 1.12.
| Table 2: | Estimated disease parameters from the zucchini squash virus epidemics from fitted models |
 | |
The nugget which is a representation of the localized discontinuity, was estimated to be 0.07300. The Range of Spatial Dependence (RSD) and the sill were also estimated to be 0.63 m and 1.91400, respectively. When the angle relative to the principal axis was tilted from the initial 0-90°, the nugget variance increased to 1.58300 but the sill and the RSD remained unchanged.
Inverse distance weighting interpolation and IDW map: The Inverse Distance Weighting (IDW) interpolation model estimated large clusters of possible infections around the primary infection foci which largely occurred in quadrats five and nine. The majority of the interpolated clusters of higher incidences were toward the eastward and South-Eastern-ward direction of the disease field, particularly around the borders of these directions. The interpolated clusters of least incidences were also estimated to be in the mid-portion of the field, with few patches on the middle section of the Western border.
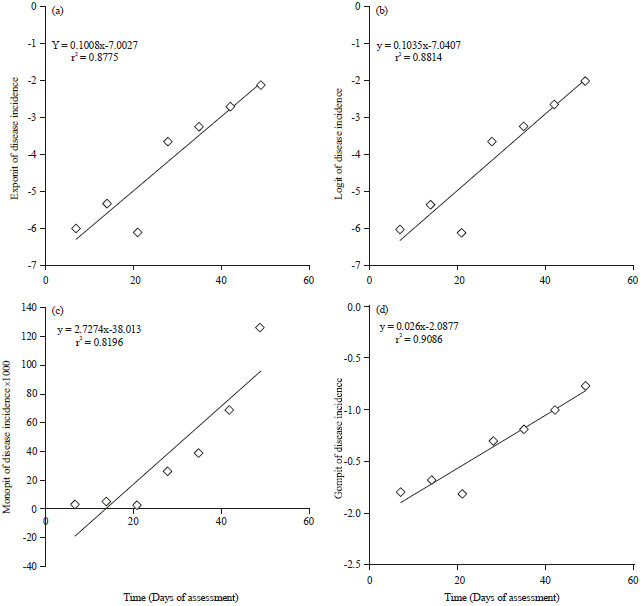 | |
| Fig. 2(a-d): | Scatterplots of mechanistic model transformed disease incidence data from zucchini squash epidemics, (a) Exponit, (b) Logit, (c) Monopit and (d) Gompit of disease severity index |
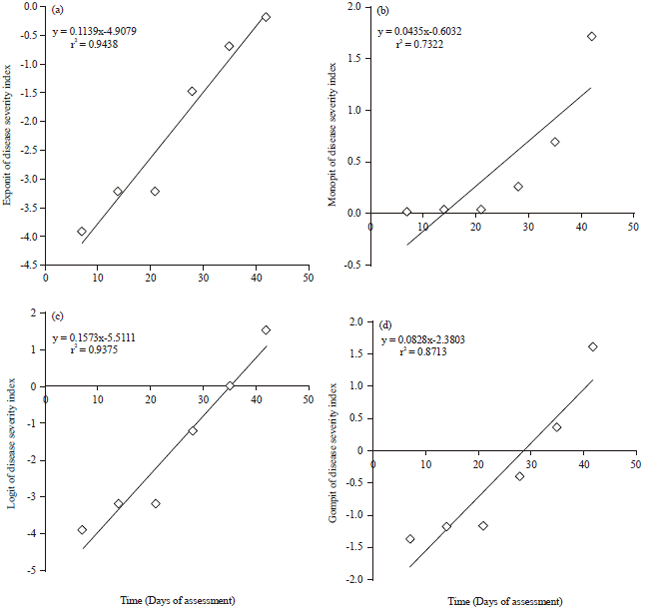 | |
| Fig. 3(a-d): | Scatterplots of mechanistic model transformed disease severity data from the zucchini squash field, (a) Exponit, (b) Logit, (c) Monopit and (d) Gompit of disease severity index |
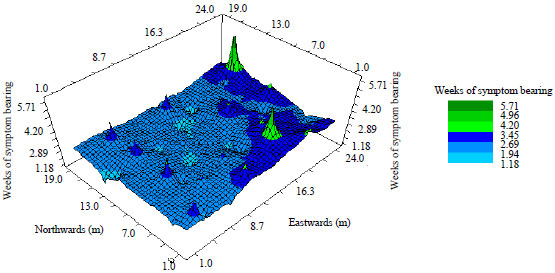 | |
| Fig. 4: | 3-D Inverse Distance Weighting (IDW) Map with peaks and colour-coded regions corresponding to the level of disease intensity across the zucchini squash field |
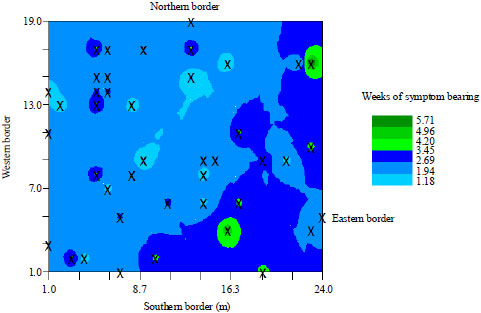 | |
| Fig. 5: | 2-D Inverse Distance Weighting (IDW) map showing the locations of symptomatic zucchini squash plants across the zucchini squash field |
There were also isolated clusters of smaller sized incidence peaks toward the western section of the squash field. Figure 4 and 5 show 2-dimensional and 3-dimensional maps of the virus epidemic, respectively, on the zucchini squash field generated through IDW interpolation.
DISCUSSION
Temporal spread patterns of viral diseases on zucchini squash field: Among four models employed in the study, the gompertz model provided the best fit to the disease incidence data, with a coefficient of determination of 90.86%. Earlier workers (Vuori et al., 2006; FAO., 2013; Berger, 1981) have indicated the satisfactory use of the gompertz model in describing disease progress curves. The model is noted for its consistent and stable parameter estimates in pathosystems of oats, potato, celery, bean, rice and several others (Mohapatra et al., 2008). The gompertz curve exhibits a decline in apparent infection rate as disease intensity increases and this is actually observed in practice (Fleming, 1983). As more and more plants become infected, the probability of new infections decline due to decline in the proportion of healthy uninfected plants in the field. This accounts for the decline in the apparent infection rate as the disease intensity increases. The gompertz model is also known to be ideal for modeling time series events such as disease progress where the initial growth and growth toward the end of the time series are slowest with an exponential growth in the mid-season of the growth. This nature of disease progress was observed in the zucchini squash field. Disease incidence started at a very low value of 0.24% and by 21 DAP, due to loss of few symptomatic plants, incidence had reduced to 0.22%, however, incidence increased exponentially from 28 DAP at a value of 2.5-11.82% by 49 DAP. As at the last week of disease assessment (49 DAP), incidence was still increasing at an increasing rate (Fig. 1). This shows that though, at this stage, the zucchini squash plants had reached their maturity, the disease progress was still in its mid-season.
The Gomp-Exp law was postulated to explain the exponential growth observed in growth curves that are best described by gompertz functions and hence supposed to follow a gompertz law and not an exponential law. As at the last week of assessment, the critical size threshold had not been reached since incidence was only 11.82%. This explains why the rate of apparent infection was still increasing at an increasing rate from the 42 DAP to the 49 DAP (Fig. 1).
Though the gompertz model was more superior in fitting the incidence data, in terms of estimating time saved due to the sanitation measure, all three mechanistic models were consistent in their estimation. According to the gompertz function, the incidence observed at 49 DAP would have doubled by 64 DAP. The estimated apparent infection rate (r) of 0.026 from the gompertz model is the sum of expression of the magnitude of the susceptibility of the zucchini squash plants to virus infection, the effects of environmental factors such as temperature, precipitation, the efficiency of the insect vectors and other factors on the progress of the epidemic in a physical quantifiable term. The effects of seed infection rate, quarantine measures, crop rotation schemes and vertical resistance genes in the host plants are not quantified by the secondary infection rate but rather by the initial disease parameter (yo) (Madden, 2012).
Spatial spread pattern of virus disease in zucchini squash field: Estimated parameters from the Gaussian model and a subsequent inverse distance weighting form of mapping showed that there was significant variability in disease incidence and the level of aggregation of diseased plants across the quadrats.
There was almost 22 fold increase in the nugget variance due to the tilting of the principal axis through an angle of 90° confirming the variability of aggregation of diseased plants across the field. The nugget variance, also termed as localized discontinuity, measures the extent of aggregation of diseased plants across the field in a given perspective. The initial 0.0730 nugget variance showed that from the 0° perspective of the field, more of the diseased plants were closer to each other compared to the level of aggregation from the 90° perspective of the field. This phenomenon is termed as anisotropy; thus, on the same field, the level of closeness of diseased zucchini squash plants was not uniform but rather there existed a directional difference in the level of spatial associations among the diseased plants. If the level of aggregation of diseased plants were uniform across the field, then the nugget variance would have remained constant irrespective of the angle of rotation of the principal axis.
The low Range of Spatial Dependence (RSD) of 0.63 m indicates the magnitude of closeness of diseased zucchini squash plants in the field. The variability of incidence and level of aggregation of diseased squash plants were also confirmed from the Inverse Distance Weighting (IDW) map. The IDW analysis and its subsequent map estimated two major primary disease foci which were all toward the Eastern section of the field and in accordance with the theories underlying IDW mapping, the model predicted clusters of possible high levels of incidence around the two major disease foci (Fig. 4 and 5).
However, a significant number of the symptomatic zucchini squash plants did not fall within the model predicted areas of high incidences. The early loss of one of the initially diseased plants in the Eastern border region of the field, the close proximity of an already established cucumber field to the Western border of the zucchini squash field and the behavioral pattern of insect vectors in the field are proposed to account for the observed variability in incidence and the level of disease aggregation as well as the significant departure of symptomatic plants from the model predicted zones of high incidence. Viruses depend on vectors for their transmission from plant to plant due to their inability to break the physical barriers, such as cuticle and the cellulose cell walls of their hosts (Esker et al., 2007). Therefore, the behaviour of insect vectors, particularly in relation to virus transmission, significantly influences the pattern of observed disease in a field (Jeger et al., 2004). The estimated closeness of symptomatic plants with an RSD of just 0.63 m may suggest that insect vectors fed from clusters of plants that were within a smaller circumference, moving from one plant to another nearby plant, rather than long distances of flight within an infinitesimally small amount of time in search of hosts. If such proposed pattern of insect feeding is true, then the loss of one of the initially diseased plants in the Eastern border region of the field and the not more than 2 m proximity of a cucumber field to the Western border of the zucchini squash field may explain why a significant number of plants did not fall within the model predicted zones of high incidence in the Eastern part of the field and the high level of aggregation of symptomatic plants in the Western part of the field. Particularly in the upper section of the Eastern part of the field, since the initially infected plant no longer existed from the early part of the epidemic, it could not have served as a source of inoculum to infect zucchini squash plants in its vicinity of which the model had predicted higher incidence. In the Western section of the field, the close proximity of the cucumber plants may have served as ready source of inoculum to infect nearby squash plants in the western section of the field.
Quantification of disease progress curves in zucchini squash through mathematical modeling revealed more insight into the virus epidemic which could otherwise not have been possible. The spatial model and Inverse Distance Weighting maps estimated areas, particularly toward the Eastern border of the field, where disease intensities were expected to be high. A significant number of symptomatic plants did not fall within these critical zones; this could have been as a result of the proximity of the already established cucumber field to the western border of the zucchini squash field.
ACKNOWLEDGMENT
We appreciate the support from the staff of the Biotechnology and Nuclear Agriculture Research Institute of the Ghana Atomic Energy Commission, Ghana.
REFERENCES
- Berger, R.D., 1981. Comparison of the Gompertz and logistic equations to describe plant disease progress. Phytopathology, 71: 716-719.
CrossRefDirect Link - Esker, P.D., A.H. Sparks, G. Antony, M. Bates and W. Dall' Acqua et al., 2007. Ecology and Epidemiology in R: Modeling dispersal gradients. Plant Health Instructor.
CrossRef - Fekedulegn, D., M.P.M. Siurtain and J.J. Colbert, 1999. Parameter estimation of nonlinear growth models in forestry. Silva Fennica, 33: 327-336.
Direct Link - Fleming, R.A., 1983. Development of a simple mechanistic model of cereal rust progress. Phytopathology, 73: 308-312.
Direct Link - Jeger, M.J., J. Holt, F. Van Den Bosch and L.V. Madden, 2004. Epidemiology of insect-transmitted plant viruses: Modelling disease dynamics and control interventions. Physiol. Entomol., 29: 291-304.
CrossRefDirect Link - Mohapatra, N.K., A.K. Mukherjee, A.V.S. Rao and P. Nayak, 2008. Disease progress curves in the rice blast pathosystem compared with the logistic and gompertz models. J. Agric. Biol. Sci., 3: 28-37.
Direct Link - Roumagnac, P., O. Pruvost, F. Chiroleu and G. Hughes, 2004. Spatial and temporal analyses of bacterial blight of onion caused by Xanthomonas axonopodis pv. allii. Phytopathology, 94: 138-146.
CrossRefDirect Link - Teng, P.S., 1985. A comparison of simulation approaches to epidemic modeling. Ann. Rev. Phytopathol., 23: 351-379.
CrossRefDirect Link - Vuori, K., I. Stranden, M. Sevon-Aimonen and E.A. Mantysaari, 2006. Estimation of non-linear growth models by linearization: a simulation study using a Gompertz function. Genet. Sel. Evol., 38: 343-358.
Direct Link