
Wafaa M. Haggag
Department of Plant Pathology, National Research Centre, Giza, Dokki, Egypt
Faten M. Abd-El-Latif
Department of Plant Pathology, Faculty of Agriculture, Benha University, Egypt
Riad S.R. El-Mohamedy
Department of Plant Pathology, National Research Centre, Giza, Dokki, Egypt
Plant Pathology Journal
Year: 2015 | Volume: 14 | Issue: 4 | Page No.: 242-249







ABSTRACT
Physical and biochemical integrated management using soil solarization and chitin or chitosan singly or in combination for controlling tomato wilt disease under field conditions was studied. Chitin had no inhibitory effect on the growth of pathogenic fungus. On the other hand, chitosan at 6 g L–1 completely inhibit the linear growth and spore germination of F. oxysporum f. sp. lycopersici. Under field conditions, results indicated that maximum soil temperatures in solarized were recorded 55.2, 50.8 and 46.3°C at depths of 1-10, 11-20 and 21-30 cm of soil surface. Solarization was more effective in reducing the pathogen population. The highest reduction in total count of F. oxysporum f. sp. lycopersici was observed in mulched soil at 1-10 and 11-20 cm depths. Soil solarization and chitin at 6.0 g kg–1 soil, chitosan at 6.0 g kg–1 soil or topsin at 3.0 g kg–1 soil singly or in combination for controlling tomato, wilt disease under field conditions was evaluated. Results revealed that all treatments significantly reduced the disease incidence and severity during two growing seasons. The highest reduction in disease incidence and severity were obtained with combined treatments between soil solarization and chitin, chitosan or topsin which recorded 3.2-5.0% as disease incidence and 0.2-0.3% as disease severity during two growing seasons. As for tomato yield the highest increase in tomato yield was obtained with combined treatments between soil solarization and chitin, chitosan or topsin which increased the tomato yield more than 66.7, 68.9 and 66.7% during two growing seasons. All treatments significantly increased the chitinase activity of tomato plants. The most effective treatments were combined treatments between soil solarization and chitin or chitosan which increased the chitinase activity by 100.0 and 116.7%. It could be suggested that combined treatments between soil solarization and chitin or chitosan as safety materials might be used commercially for controlling tomato wilt diseases under field conditions.
PDF Abstract XML References Citation
Received: July 12, 2015;
Accepted: September 19, 2015;
Published: May 03, 2019
How to cite this article
Wafaa M. Haggag, Faten M. Abd-El-Latif and Riad S.R. El-Mohamedy, 2015. Physical and Biochemical Integrated Management For Controlling Tomato Wilt Disease Under Field Conditions. Plant Pathology Journal, 14: 242-249.
DOI: 10.3923/ppj.2015.242.249
URL: https://scialert.net/abstract/?doi=ppj.2015.242.249
DOI: 10.3923/ppj.2015.242.249
URL: https://scialert.net/abstract/?doi=ppj.2015.242.249
INTRODUCTION
Fusarium wilt caused by Fusarium oxysporum f. sp. lycopersici (Sacc.), is one of the major yield limiting factors of tomato (Solanum lycopersicum) which causes serious economic and yield losses. Major strategies for controlling this disease are soil fungicides (De Cal et al., 2000; Attitalla et al., 2001; Ojha and Chatterjee, 2012).
Soil solarization is an environmentally friendly method of using solar power for controlling pests such as soilborne plant pathogens including fungi, bacteria, nematodes and insect and mite pests along with weed seed and seedlings in the soil by mulching the soil and covering it with tarp, usually with a transparent polyethylene cover, to trap solar energy. It may also describe methods of decontaminating soil using sunlight or solar power. This energy causes physical, chemical and biological changes in the soil. Soil solarization was carried out as transparent polyethylene plastic placed on moist soil during the hot summer months increases soil temperatures to levels lethal to many soil-borne plant pathogens, weeds and nematodes (Abd-El-Kareem et al., 2004; Culman et al., 2006; Farrag and Fotouh, 2010; Saied-Nehal, 2011).
Chitin and chitosan are naturally-occurring compounds that have potential in agriculture with regard to controlling plant diseases (Abd-El-Kareem and Haggag, 2014). These molecules were shown to display toxicity and inhibit fungal growth and development. They were reported to be active against viruses, bacteria and other pests. Chitosan applied as seed or soil treatments was shown to control Fusarium wilts in many plant species (Badawy et al., 2005). Chitosan induce host defense responses in both monocotyledons and dicotyledons. These responses include lignification, ion flux variations, cytoplasmic acidification, membrane depolarization and protein phosphorylation, chitinase and glucanase activation, phytoalexin biosynthesis, generation of reactive oxygen species, biosynthesis of jasmonic acid and the expression of unique early responsive and defense-related genes. In addition, chitosan was reported to induce callose formation, proteinase inhibitors and phytoalexin biosynthesis in many dicot species (Uppal et al., 2008; Elwagia and Algam, 2014; Mishra et al., 2014; Saied-Nehal, 2015).
Chitin was reported as resistance inducer against soilborne diseases (Kuchitsu et al., 1993; Bell et al., 1998). Addition of small quantities of chitin to soil resulted in a marked reduction in root rot diseases of some plants (Kuchitsu et al., 1993; Bell et al., 1998; Abd-El-Kareem et al., 2006). Based on these and other proprieties that help strengthen host plant defenses, interest has been growing in using them in agricultural systems to reduce the negative impact of diseases on yield and quality of crops.
The purpose of the present work was designed to evaluate the effect of soil solarization and chitin or chitosan singly or in combination for controlling tomato wilt disease under field conditions.
MATERIALS AND METHODS
Source of pathogenic fungus and tomato transplants: Pathogenic isolate of Fusarium oxysporum f. sp. lycopersici the causal agent of tomato wilt disease was kindly provided by Department of Plant Pathology (Project Integrated management for controlling tomato fungal diseases under Egyptian and Tunisian conditions), National Research Centre, Giza, Egypt. Tomato transplants cv. Kastel rock were obtained from the Department of Vegetable Crop Research, Agricultural Research Centre, Giza, Egypt.
Laboratory experiments
Effect of chitin and chitosan on the linear growth and spore germination of Fusarium oxysporum f. sp. lycopersici: The inhibitory effect of chitin and chitosan (Sigma company) against F. oxysporum f. sp. lycopersici (FOL) was tested in vitro at four concentrations, i.e. 0, 2, 4 and 6 g L–1. Chitin or chitosan were added to conical flasks containing sterilized PDA medium before its solidifying and rotated gently then disbanded into sterilized Petri-plates (9 cm diameter). Plates were individually inoculated at the centre with equal disks (6 mm diameter) taken from 10 days old culture of Fusarium oxysporum f. sp. lycopersici then incubated at 25±2°C. Linear growth of tested fungus was measured when the control plates (medium free of chitin or chitosan) reached full growth and the average growth diameter was calculated. As for spore germination test, spore suspension was prepared by culturing FOL on Petri-plates containing PDA medium for 20 days at 25°C. Colony Forming Units (CFU) containing hyphal fragments, microconidia, macroconidia and chlamydospores were released in sterilized water using a needle then adjusted to 106 CFU mL–1 using haemocytometers slide. One milliliter of suspension was transferred to each test tubes containing sterilized broth PD medium which treated with previous concentrations of culture filtrates. Test tubes were incubated for 24 h at 25°C. One milliliter of treated spore suspension (CFU) was examined microscopy and the average percent of spore germination was calculated. Each treatment was represented by 5 plates as replicates.
Field experiments
Efficacy of soil solarization and chitin or chitosan singly or in combination for controlling tomato wilt disease: The field experiment carried out at the Experimental Research Station of National Research Centre at El-Noubariya region, Behera Governorate, Egypt. Under field conditions, the efficacy of soil solarization and chitin or chitosan singly or in combination for controlling tomato wilt disease was studied.
The field trail conducted in 60 plots, each 12 m2 (3 m width×4 m length) comprised of 5 rows with 4 m length and 20 transplants/row, established in naturally heavily infested soil with tomato wilt pathogens. All plots were irrigated to field capacity and 30 plots were subjected to solarization treatment which carried out by covering with 100 μm thick transparent polyethylene sheets for 4 weeks during August then removed. The other untreated 30 plots were considered as un solarized soil. Maximum and minimum degrees of soil temperature were regularly measured during that period at the depths of 1-10, 11-20 and 21-30 cm of surface soil. The average temperature at the three soil depths was calculated at the end of mulching period.
Treatments: The following treatments were applied:
| • | Single treatment: |
| • | Soil solarization for 4 weeks |
| • | Chitosan at 6.0 g kg–1 soil |
| • | Chitin at 6.0 g kg–1 soil |
| • | Topsin at 3 g kg–1 soil |
| • | Combined treatment: |
| • | Soil solarization+chitosan at 6.0 g kg–1 soil |
| • | Soil solarization+chitin at 6.0 g kg–1 soil |
| • | Soil solarization+topsin at 3 g kg–1 soil |
| • | Control (Un-treated) |
Application
Seed bed treatment: Tomato transplants cv. Kastle rock which grown in foam trays (209 cells) containing peat-moss soil mixed individually with chitin or chitosan at 6.0 g kg–1 soil as well as topsin at 3 g kg–1 soil. In addition to tomato transplants grown in free soil treatments in foam trays served as control.
Effect of soil mulching on population density of Fusarium oxysporum f. sp. lycopersici
Preparation of Fusarium oxysporum f. sp. lycopersici inoculum: Inoculum of F. oxysporum f. sp. lycopersici was prepared by culturing the fungus on 50.0 mL Potato Dextrose Broth (PDB) medium in 250 mL Erlenmeyer flasks for 15 days at 25°C. Inoculum of F. oxysporum f. sp. lycopersici was prepared from the growing upper solid layers which washed and blended in sterilized water. Colonies forming units (CFU) were adjusted to 106 CFU mL–1 using haemocytometers slide. Soil infestation was carried out at rate of 50 mL (106 CFU mL–1)/kg soil (Elad and Baker, 1985).
Soil infestation: Certain weights of field soil was sterilized with autoclave at 120°C for 1 h and artificially infested with F. oxysporum f. sp. lycopersici. Soils were mixed individually with the prepared inoculum.. Soil infestation was carried out at rate of 50 mL (106 CFU mL–1)/kg soil (Elad and Baker, 1985). Artificially infested soils were filled into cloths bags at the rate of 1 kg soil/bag and 40 bags was used. Before soil mulching cloths bags were buried into the field soil at three different levels down below the surface at depths of 1-10, 11-20 and 21-30 cm at three spots of each plot and three plots were used.
After removal the polyethylene sheets, the buried bags of each certain level in either solarized or un-solarized plots were collected and mixed into one mother composite sample for each tested fungus. Total count of pathogenic fungus in solarized and un-solarized soil compared with their count before soil mulching was estimated following the plate count technique (Porras et al., 2007).
Determination of total count of pathogenic fungus: Total count of pathogenic fungus was carried out according to the method described by Porras et al. (2007) as mentioned before. The resulting colonies were calculated as colonies per gram of dry soil and the reduction was calculated as follow:
Disease assessment: The disease incidence was recorded as percentage of wilted plants after 80 days of transplanting. Meanwhile, The disease severity was recorded by 0-4 scale as described by Biswas et al. (2012), where zero represents no infection and four denotes complete infection.
The scale of 0-4 of the disease severity was modified as follows:
| • | 0-No infection |
| • | 1-Slight infection, which is about 25% of leaves turned yellow and wilted |
| • | 2-Moderate infection, which is about 50% of leaves turned yellow and wilted |
| • | 3-Extensive infection, which is about 75% of leaves turned yellow and wilted |
| • | 4-Complete infection, which is about 100% of leaves wilted and the plants died |
Determination of tomato yield: Accumulation of tomato yield (kg m–2) for each treatment was determined.
Effect of soil solarization and chitin or chitosan singly or in combination on chitinase activity of tomato plants: The efficacy of soil solarization and chitin or chitosan singly or in combination on chitinase activity of tomato plants was studied.
Extraction of chitinase enzyme: Chitinase activity was determined after 60 days of transplanting. Extraction of enzyme from tomato leaves was done according to method of Tuzun et al. (1989).
Chitinase assay: Chitinase activity was determined by colourimetric method of Boller and Mauch (1988). Colloidal chitin was used as a substrate and dinitrosalicylic acid as reagent to measure reducing sugars.
Chitinase activity was expressed as millimolar N-acetylglucose amine equivalent released/gram fresh weight tissue/60 min.
Statistical analysis: Tukey test for multiple comparisons among means was utilized (Neler et al., 1985).
RESULTS
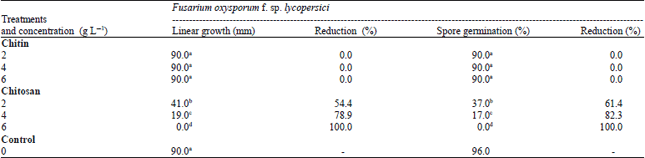
Effect of chitin and chitosan on the linear growth and spore germination of Fusarium oxysporum f. sp. lycopersici: The inhibitory effect of chitin and chitosan against F. oxysporum f. sp. lycopersici was tested in vitro at four concentrations, i.e. 0, 2, 4 and 6 g L–1. Results in Table 1 indicate that chitin has no inhibitory effect on the growth of pathogenic fungus.
| Table 1: | Linear growth and spore germination of Fusarium oxysporum f. sp. lycopersici as affected by different concentrations of chitin and chitosan |
 | |
| Values with the same letter are not significantly different (p = 0.05) | |
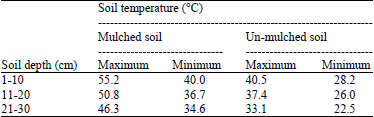
| Table 2: | Average of maximum and minimum soil temperatures at different soil depths in mulched and un-mulched soil |
 | |
| Table 3: | Percentage reduction in pathogenic fungus at three depths as affected with mulched and un-mulched soil under field conditions |
 | |
On the other hand, all concentrations of chitosan had significant inhibitory effect against F. oxysporum f. sp. lycopersici. chitosan at 6 g L–1 completely inhibit the linear growth and spore germination of F. oxysporum f. sp. lycopersici. The highest reduction was obtained with chitosan at 4 and 6 g L–1 which reduced the linear growth and spore germination by 78.9, 82.3 and 100% in both linear growth and spore germination, respectively. Meanwhile, chitosan at 2 g L–1 was less effective.
Effect of soil solarization on soil temperatures: Average of maximum and minimum soil temperatures in solarized and non-solarized soil was recorded during solarization period. Results in Table 2 indicate that maximum soil temperatures in solarized were recorded 55.2, 50.8 and 46.3°C at depths of 1-10, 11-20 and 21-30 cm of soil surface.
Meanwhile in un-solarized soil recorded 40.5, 37.4 and 33.1°C as maximum soil temperatures at three depths, respectively.
Effect on total count of pathogenic fungus: The population density of F. oxysporum f. sp. lycopersici at three soil depths either in mulched or un-mulched soil was determined. Results in Table 3 indicate that fungal population decreased in both mulched and un-mulched soils at the end of experimental period. However, solarization was more effective in reducing the pathogen population. The highest reduction in total count of F. oxysporum f. sp. lycopersici was observed in mulched soil at 1-10 and 11-20 cm depths. As for the lower soil depth, 21-30 cm, the pathogen population was reduced by 60.1%. The same trend was also noticed in fallow un-mulched soil at the three similar depths, i.e., 23.4, 17.9 and 7.8%, respectively.
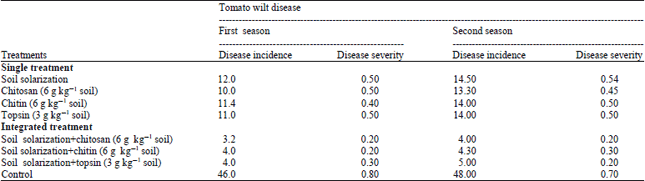
Efficacy of soil solarization and chitosan, chitin or topsin singly or in combination for controlling tomato wilts disease: Soil solarization and chitosan at 6.0 g kg–1 soil, chitin at 6.0 g kg–1 soil or topsin at 3.0 g kg–1 soil singly or in combination for controlling tomato wilt disease under field conditions was evaluated. Results in Table 4 reveal that all treatments significantly reduced the disease incidence and severity during two growing seasons. The highest reduction in disease incidence and severity were obtained with combined treatments between soil solarization and chitin, chitosan or topsin which recorded 3.2-5.0% as disease incidence and 0.2-0.3 as disease severity during the two growing seasons. Meanwhile, single treatments show moderate effect.
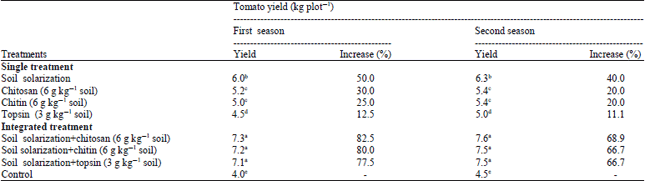
Efficacy of integrated treatments between soil solarization and chitosan, chitin or topsin on tomato yield under field conditions: Results in Table 5 soil solarization and chitosan at 6.0 g kg–1 soil, chitin at 6.0 g kg–1 soil or topsin at 3.0 g kg–1 soil singly or in combination on tomato yield under field conditions was evaluated. Results in Table 5 reveal that all treatments significantly increased the tomato yield during the two growing seasons. The highest increase in tomato yield was obtained with combined treatments between soil solarization and chitin, chitosan or topsin which increased the tomato yield more than 66.7, 68.9 and 66.7% during the two growing seasons. Meanwhile, single treatments show moderate increase.
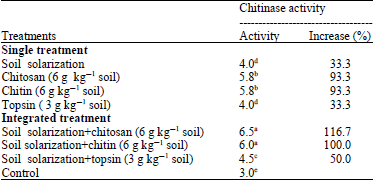
Effect of soil solarization and chitosan, chitin or topsin singly or in combination on chitinase activity of tomato plants: The efficacy of soil solarization and chitosan, chitin or topsin singly or in combination on chitinase activity of tomato plants was studied.
Results in Table 6 indicate that all treatments significantly increased the chitinase activity of tomato plants. The most effective treatments were combined treatments between soil solarization and chitosan or chitin which increased the chitinase activity by 116.7 and 100.0%.
| Table 4: | Efficacy of integrated treatments between soil solarization and chitosan, chitin or topsin on tomato wilt disease under field conditions |
 | |
| Values with the same letter are not significantly different (p = 0.05) | |
| Table 5: | Efficacy of integrated treatments between soil solarization and chitosan, chitin or topsin on tomato yield under field conditions |
 | |
| Values with the same letter are not significantly different (p = 0.05) | |
| Table 6: | Efficacy of integrated treatments between soil solarization and chitosan, chitin or topsin on chitinase activity of tomato under field conditions |
 | |
| Values with the same letter are not significantly different (p = 0.05) | |
Meanwhile, single treatments of chitin or chitosan resulted in increasing chitinase activity by 93.3%. Other treatments show moderate increase. While, solarization alone or topsin alone increase chitinase activity by 33.3% only.
DISCUSSION
Fusarium wilt caused by Fusarium oxysporum f. sp. lycopersici (Sacc.), is one of the major yield limiting factors of tomato (Solanum lycopersicum) which causes serious economic and yield losses (De Cal et al., 2000; Attitalla et al., 2001; Ojha and Chatterjee, 2012).
Chitosan applied as seed or soil treatments was shown to control Fusarium wilts in many plant species (Badawy et al., 2005; Saied-Nehal, 2015). Chitosan induce host defense responses in both monocotyledons and dicotyledons (Uppal et al., 2008; Elwagia and Algam, 2014; Mishra et al., 2014). In the present study results revealed that chitin had no inhibitory effect on the growth of pathogenic fungus. On the other hand, chitosan at 6 g L–1 completely inhibit the linear growth and spore germination of F. oxysporum f. sp. lycopersici. In this respect, Kulikov et al. (2006) reported that the antimicrobial activity increased with the increase in chitosan molecular weight and seems to be faster on fungi and algae than on bacteria. Fungicidal activity of chitosan has been documented against various species of fungi and oomycetes (Vasyukova et al., 2000; El-Mohamedy et al., 2013). The minimal growth-inhibiting concentrations varied between 10 and 5,000 ppm (Rabea et al., 2005). Some of the derivatives also repressed spore formation at rather high concentrations (Badawy et al., 2005). Recently, Palma-Guerrero et al. (2009) demonstrated that chitosan is able to permeabilize the plasma membrane of Neurospora crassa and kills the cells. In general, chitosan is able to reduce the in vitro growth of a number of fungi and oomycetes (Palma-Guerrero et al., 2008). For instance, chitosan was reported to exert an inhibitory action on the hyphal growth of numerous pathogenic fungi, including root and necrotrophic pathogens, such as Fusarium oxysporum, Botrytis cinerea, Monilina laxa, Alternaria alternata and Pythium aphanidermatum (El Hassni et al., 2004). The mechanism by which chitosan affects the growth of several pathogenic fungi has not been fully elucidated, but several hypotheses have been postulated, first: its polycationic nature, it is believed that chitosan interferes with negatively charged residues of macromolecules exposed on the fungal cell surface. This interaction leads to the leakage of intracellular electrolytes and proteinaceous constituents (El Hassni et al., 2004). Second the interaction of diffused hydrolysis products with microbial DNA, which leads to the inhibition of mRNA and protein synthesis (Vasyukova et al., 2000), third the chelating of metals, spore elements and essential nutrients (Rabea et al., 2005). Forth the interaction of chitosan with fungal DNA and RNA (Palma-Guerrero et al., 2008) and five the malformation of fungal mycelial. Chitosan is not only effective in inhibition the growth of the pathogen fungi, but also induces marked morphological changes, structural alterations and molecular disorganization of fungal cells (Barka et al., 2004). Moreover, El Hassni et al. (2004) reported that, chitosan caused morphological changes such as large vesicles or empty cells devoid of cytoplasm in the mycelium of B. cinerea. Furthermore, Bautista-Banos et al. (2006) revealed that by microscopic observation of fungi treated with chitosan, it can affect the morphology of the hyphae.
On the other hands, chitosan induce host defense responses against several plant diseases (Uppal et al., 2008; Elwagia and Algam, 2014; Mishra et al., 2014; Saied-Nehal, 2015). In the present study, under field conditions results revealed that the highest reduction in disease incidence and severity were obtained with combined treatments between soil solarization and chitin, chitosan or topsin which recorded 3.2-5.0% as disease incidence and 0.2-0.3% as disease severity during two growing seasons. As for tomato yield the highest increase in tomato yield was obtained with combined treatments between soil solarization and chitin, chitosan or topsin which increased the tomato yield more than 66.7, 68.9 and 66.7% during two growing seasons.
All treatments significantly increased the chitinase activity of tomato plants. The most effective treatments were combined treatments between soil solarization and chitin or chitosan which increased the chitinase activity by 100.0 and 116.7%. In this regard, chitosan has been extensively utilized as a foliar treatment to control the growth, spread and development of many diseases involving viruses, bacteria, fungi and pests (Badawy et al., 2005; Faoro et al., 2008; Elwagia and Algam, 2014). It has also been used to increase yield and tuber quality of potatoes (Kowalski et al., 2006). Chitosan had different properties i.e. had inhibitory effect against pathogenic fungus and had ability to be potent elicitors of plant defense resistance.
Chitin was reported as resistance inducer against soil borne diseases (Kuchitsu et al., 1993, Bell et al., 1998; Abd-El-Kareem and Haggag, 2014). Addition of small quantities of chitin to soil resulted in a marked reduction in root rot diseases of some plants (Kuchitsu et al., 1993; Bell et al., 1998). In the present study, results indicated that under field conditions results revealed that the highest reduction in disease incidence and severity were obtained with combined treatments between soil solarization and chitin, chitosan or topsin and increased tomato yield. Also, combined treatments between soil solarization and chitin or chitosan increased the chitinase activity by 100.0 and 116.7%.
Soil solarization was carried out as transparent polyethylene plastic placed on moist soil during the hot summer months increases soil temperatures to levels lethal to many soil-borne plant pathogens, weeds and nematodes (Abd-El-Kareem et al., 2004; Culman et al., 2006; Farrag and Fotouh, 2010; Saied-Nehal, 2011). In the present study, under field conditions results revealed that the highest reduction in disease incidence and severity were obtained with combined treatments between soil solarization and chitin, chitosan or topsin and increased tomato yield. All treatments significantly increased the chitinase activity of tomato plants. The most effective treatments were combined treatments between soil solarization and chitin or chitosan.
In the current study, the population density of FOL was determined in artificially infested soil at three soil depths either in mulched or un-mulched soil.
Reduction in disease incidence and increase in obtained yield due to soil solarization were reported by many investigators (Osman et al., 1986). Pullman et al. (1981) presented a detailed study on thermal death of four soil-borne plant pathogens as affected by time and temperature of the treatment. They reported that R. solani was found to be killed at 50°C in only 10 min as exposure time. In the present study, the recorded maximum soil temperature measured at 1-10, 11-20 and 21-30 cm of soil depths in mulched soil reached 55.2, 50.8 and 46.3°C in average during 6 weeks of mulching period. Several investigators reported the effect of soil solarization in reducing the incidence and severity of plant diseases caused by soil-borne pathogens (Abd-El-Kareem et al., 2004; Culman et al., 2006; Saied-Nehal, 2011).
It could be suggested that combined treatments between soil solarization and chitin or chitosan as safety materials might be used commercially for controlling tomato wilt diseases under field conditions.
REFERENCES
- Abd-El-Kareem, F., M.A. Abd-Allah, N.G. El-Gamal and N.S. El-Mougy, 2004. Integrated control of Lupin root rot disease in solarized soil under greenhouse and field condition. Egypt. J. Phytopathol., 32: 49-63.
Direct Link - Abd-El-Kareem, F., N.S. El-Mougy, N.G. El-Gamal and Y.O. Fotouh, 2006. Use of chitin and chitosan against tomato root rot disease under greenhouse conditions. Res. J. Agric. Biol. Sci., 2: 147-152.
Direct Link - Abd-El-Kareem, F. and W.M. Haggag, 2014. Chitosan and citral alone or in combination for controlling early blight disease of potato plants under field conditions. Res. J. Pharmaceut. Biol. Chem. Sci., 5: 941-949.
Direct Link - Barka, E.A., P. Eullaffroy, C. Clement and G. Vernet, 2004. Chitosan improves development and protects Vitis vinifera L. against Botrytis cinerea. Plant Cell. Rep., 22: 608-614.
CrossRefDirect Link - Attitalla, I.H., P. Johnson, S. Brishammar and P. Quintanilla, 2001. Systemic resistance to Fusarium wilt in tomato induced by Phytopthora cryptogea. J. Phytopathol., 149: 373-380.
CrossRefDirect Link - Badawy, M.E.I., E.I. Rabea, T.M. Rogge, C.V. Stevens, W. Steurbaut, M. Hofte and G. Smagghe, 2005. Fungicidal and insecticidal activity of O-acyl chitosan derivatives. Polym. Bull., 54: 279-289.
CrossRefDirect Link - Rabea, E.I., M.E.I. Badawy, T.M. Rogge, C.V. Stevens, M. Hofte, W. Steurbaut and G. Smagghe, 2005. Insecticidal and fungicidal activity of new synthesized chitosan derivatives. Pest Manage. Sci., 61: 951-960.
CrossRefPubMedDirect Link - Bautista-Banos, S., A.N. Hernandez-Lauzardo, M.G. Velazquez-del Valle, M. Hernandez-Lopez, E.A. Barka, E. Bosquez-Molina and C.L. Wilson, 2006. Chitosan as a potential natural compound to control pre and postharvest diseases of horticultural commodities. Crop Prot., 25: 108-118.
CrossRefDirect Link - Bell, A.A., J.C. Hubbard, L. Lui, R.M. Davis and K.V. Subbarao, 1998. Effects of chitin and chitosan on the incidence and severity of Fusarium yellows of celery. Plant Dis., 82: 322-328.
CrossRefDirect Link - Biswas, S.K., N.K. Pandey and M. Rajik, 2012. Inductions of defense response in tomato against Fusarium wilt through inorganic chemicals as inducers. J. Plant Pathol. Microbiol., Vol. 3, No. 4.
CrossRefDirect Link - Boller, T. and F. Mauch, 1988. Colorimetric assay for chitinase. Meth. Enzymol., 161: 430-435.
CrossRefDirect Link - Culman, S.W., J.M. Duxbury, J.G. Lauren and J.E. Thies, 2006. Microbial community response to soil solarization in Nepal's rice-wheat cropping system. Soil Biol. Biochem., 38: 3359-3371.
CrossRefDirect Link - De Cal, A., R. Garcia-Lepe and P. Melgarejo, 2000. Induced resistance by Penicillium oxalicum against Fusarium oxysporum f. sp. lycopersici: Histological studies of infected and induced tomato stems. Phytopathology, 90: 260-268.
CrossRefDirect Link - Elad, Y. and R. Baker, 1985. Influence of trace amounts of cations and siderophore-producing pseudomonads on chlamydospore germination of Fusarium oxysporum. Phytopathology, 75: 1047-1052.
CrossRefDirect Link - El Hassni, M., A. El Hadrami, F. Daayf, E.A. Barka and I. El Hadrami, 2004. Chitosan, antifungal product against Fusarium oxysporum f. sp. albedinis and elicitor of defence reactions in date palm roots. Phytopathologia Mediterranea, 43: 195-204.
Direct Link - El-Mohamedy, R.S., M.M. Abdel-Kader, F. Abd-El-Kareem and N.S. El-Mougy, 2013. Inhibitory effect of antagonistic bio-agents and chitosan on the growth of tomato root rot pathogens in vitro. J. Agric. Technol., 9: 1521-1533.
Direct Link - Faoro, F., D. Maffi, D. Cantu and M. Iriti, 2008. Chemical-induced resistance against powdery mildew in barley: The effects of chitosan and benzothiadiazole. BioControl, 53: 387-401.
CrossRefDirect Link - Farrag, E.S.H. and Y.O. Fotouh, 2010. Solarization as a method for producing fungal-free container soil and controlling wilt and root-rot diseases on cucumber plants under greenhouse conditions. Arch. Phytopathol. Plant Protect., 43: 519-526.
CrossRefDirect Link - Kowalski, B., F.J. Terry, L. Herrera and D.A. Penalver, 2006. Application of soluble chitosan in vitro and in the greenhouse to increase yield and seed quality of potato minitubers. Potato Res., 49: 167-176.
CrossRefDirect Link - Kuchitsu, K., M. Kikuyama and N. Shibuya, 1993. N-acetylchitooligosaccharides, biotic elicitor for phytoalexin production, induce transient membrane depolarization in suspension-cultured rice cells. Protoplasma, 174: 79-81.
CrossRefDirect Link - Kulikov, S.N., S.N. Chirkov, A.V. Il'ina, S.A. Lopatin and V.P. Varlamov, 2006. Effect of the molecular weight of chitosan on its antiviral activity in plants. Applied Biochem. Microbiol., 42: 200-203.
CrossRefDirect Link - Mishra, S., K.S. Jagadeesh, P.U. Krishnaraj and S. Prem, 2014. Biocontrol of tomato leaf curl virus (ToLCV) in tomato with chitosan supplemented formulations of Pseudomonas sp. under field conditions. Aust. J. Crop Sci., 8: 347-355.
Direct Link - Ojha, S. and N. Chatterjee, 2012. Induction of resistance in tomato plants against Fusarium oxysporum f. sp. Lycopersici mediated through salicylic acid and Trichoderma harzianum. J. Plant Protect. Res., 52: 220-225.
CrossRefDirect Link - Palma-Guerrero, J., H.B. Jansson, J. Salinas and L.V. Lopez-Llorca, 2008. Effect of chitosan on hyphal growth and spore germination of plant pathogenic and biocontrol fungi. J. Applied Microbiol., 104: 541-553.
CrossRefDirect Link - Palma-Guerrero, J., C.I. Huang, H-B. Jansson, J. Salina, L.V. Lopez-Llorca and N.D. Read, 2009. Chitosan permeabilizes the plasma membrane and kills cells of Neurospora crassa in an energy dependent manner. Fungal Genet. Biol., 45: 585-594.
CrossRefDirect Link - Porras, M., C. Barrau, F.T. Arroyo, B. Santos, C. Blanco and F. Romero, 2007. Reduction of Phytophthora cactorum in strawberry fields by Trichoderma spp. and soil solarization. Plant Dis., 91: 142-146.
CrossRefDirect Link - Pullman, G.S., J.E. DeVay and R.H. Garber, 1981. Soil solarization and thermal death: A logarithmic relationship between time and temperature for four soilborne plant pathogens. Phytopathology, 71: 959-964.
Direct Link - Tuzun, S., M.N. Rao, U. Vogeli, C.L. Schardl and J. Kuc, 1989. Induced systemic resistance to blue mold: Early induction and accumulation of β-1,3-glucanases, chitinases and other pathogenesis-related proteins (b-proteins) in immunized tobacco. Phytopathology, 79: 979-983.
CrossRefDirect Link - Uppal, A.K., A. El Hadrami, L.R. Adam, M. Tenuta and F. Daayf, 2008. Biological control of potato Verticillium wilt under controlled and field conditions using selected bacterial antagonists and plant extracts. Biol. Control, 44: 90-100.
CrossRefDirect Link - Vasyukova, N.I., G.I. Chalenko, N.G. Gerasimova, E.A. Perekhod and O.L. Ozeretskovskaya et al., 2000. Chitin and chitosan derivatives as elicitors of potato resistance to late blight. Applied Biochem. Microbiol., 36: 372-376.
CrossRefDirect Link