
Aradhika Tripathi
Division of Plant Pathology, ICAR-Indian Agricultural Research Institute, New Delhi, 110012, India
Sunil Chandra Dubey
Division of Plant Quarantine, ICAR-NBPGR, Pusa Campus, New Delhi, 110012, India
Plant Pathology Journal
Year: 2015 | Volume: 14 | Issue: 4 | Page No.: 234-241







ABSTRACT
Rhizoctonia solani is a destructive fungal pathogen infecting wide range of crop plants including pulses causing wet root rot or web blight disease. The present study was aimed to determine the genetic diversity of R. solani populations using Sequence-Related Amplified Polymorphism (SRAP) markers. The SRAP markers were used for genetic diversity analysis of 89 isolates of R. solani belonging to 7 Anastomosis Groups (AGs) isolated from different pulse crops representing 21 states from 16 agro-ecological regions of India. Out of 30 SRAP primer combinations evaluated, 16 combinations provided amplification with 100% polymorphism and the primer combinations Me1/Em1and Me1/Em4 provided the highest number of bands (14). The isolates of R. solani showed high level of genetic variability and grouped into 7 major clusters at 35% genetic similarity by using unweighted pair group method with an arithmetic average analysis. Bootstrap analysis grouped the isolates into five major clusters at 28% genetic similarity and about 95% isolates shared common sub-grouping patterns in both the analysis. The majority of the isolates representing various AGs were grouped together into different sub-clusters. The molecular clusters did not correspond to agro-ecological regions and crops of the origin of the isolates because of the diversity in the hosts and adopt ability of the pathogen under different environmental conditions prevalent in various parts of the country. First time an attempt was made in the present study to determine the genetic variability of the R. solani populations isolated from different pulse crops representing various AGs using SRAP markers.
PDF Abstract XML References Citation
Received: July 16, 2015;
Accepted: October 03, 2015;
Published: November 09, 2015
How to cite this article
Aradhika Tripathi and Sunil Chandra Dubey, 2015. Sequence-Related Amplified Polymorphism-PCR Analysis for Genetic Diversity in Rhizoctonia solani Populations Infecting Pulse Crops in Different Agro-Ecological Regions of India. Plant Pathology Journal, 14: 234-241.
DOI: 10.3923/ppj.2015.234.241
URL: https://scialert.net/abstract/?doi=ppj.2015.234.241
DOI: 10.3923/ppj.2015.234.241
URL: https://scialert.net/abstract/?doi=ppj.2015.234.241
INTRODUCTION
Pulses are main source of protein to vegetarian population of India and also consider a source of income to the farmers due to high market price. It is second important constituent of Indian diet after cereals and can be cultivated in all types of soil under different climatic conditions. Supplementation with cereals, pulses provide perfect mixture of diet with vegetarian protein of high biological values. Pulses are also an excellent feed and fodder for livestock. They have several unique abilities as biological nitrogen fixation, carbon sequestration, low water requirement, soil melioration and capacity to withstand under harsh climate. Therefore, they are being grown in marginal lands. India is the largest producer of pulses in the world, with 24% share in the global production. The important pulse crops are chickpea, pigeon-pea, mungbean, urdbean, lentil and fieldpea. The major pulse-producing states in India are Madhya Pradesh, Maharashtra, Rajasthan, Uttar Pradesh, Karnataka and Andhra Pradesh, which together account for about 80% of the total production. Out of the production constraints responsible for yield gaps in the pulse crops, biotic stresses are considered to be the most important worldwide. Rhizoctonia solani Kühn [teleomorph-Thanatephorus cucumeris (Fr.) Donk] is a destructive soil-borne plant pathogen (Dubey et al., 2014) infecting a wide range of agricultural and horticultural crops, including legumes, worldwide causing several diseases (Ogoshi,1987; Dubey and Dwivedi, 2000). The pathogen causes considerable yield loss in mungbean, urdbean and other pulse crops in India (Dubey, 2003; Dubey et al., 2014).
Presently, the information is available for genome sequences of fungi which made it possible to develop new marker systems. An interesting modified marker technology termed as Sequence-Related Amplified Polymorphism (SRAP) (Li and Quiros, 2001) was similar to RAPD, but it was a preferential random amplification of coding regions in genome are being applied extensively in genetic linkage map construction (Li and Quiros, 2001), genetic diversity analysis (Ferriol et al., 2003; Lin et al., 2004) and comparative genetics (Li et al., 2003) of different species. Furthermore, in the genetic diversity analysis, the information derived from SRAP markers was more concordant to the morphological variability and to the evolutionary history of the morphotypes than that of other molecular markers (Ferriol et al., 2003).
However, up to now, the SRAP molecular marker had not been extensively used in the diversity analysis of phytopathogenic fungi. So, in this work, SRAP marker was used to study the genetic variability present in R. solani populations representing different Anastomosis Groups (AGs) and isolated from various pulse crops cultivated in different agro-ecological regions of India. Thus, the study showed the results of genetic diversity analysis of 89 R. solani isolates using the SRAP markers for the first time which will be of great importance to the genetics study of imperfect and basidiomycetes fungi as well as other plant pathogenic fungi at the molecular level.
MATERIALS AND METHODS
Rhizoctonia solani isolates and cultures conditions: Eighty nine isolates of Rhizoctonia solani (including 7 AG testers) isolated from diseased specimens of major pulse crops and weeds present in pulse field covering 16 agro-ecological regions from 21 states of India were used for the genetic diversity analysis. The cultures are being maintained by the authors (Table 1). Stock cultures of the different isolates were prepared on Potato Dextrose Agar (PDA) slants (potato 200, glucose 20 and agar 20 g L–1, pH 7.0).
| Table 1: | Isolates of Rhizoctonia solani used in the present study |
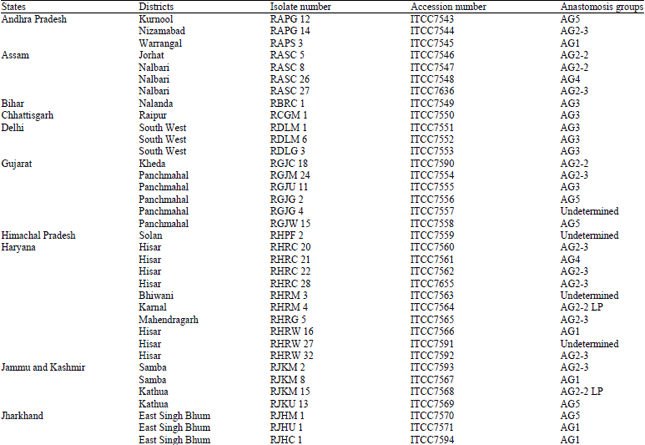 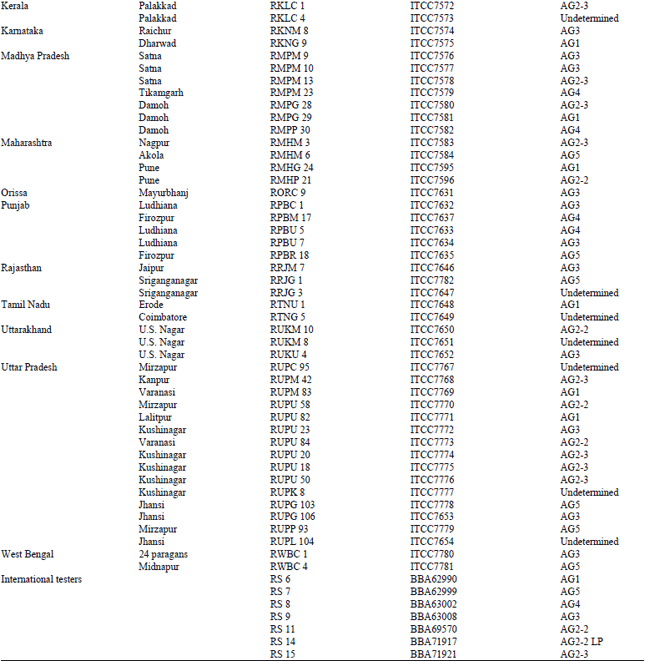 | |
| ITCC: Indian type culture collection, IARI, New Delhi, BBA: Federal Biological Research Center for Agriculture and Forestry, Germany | |
After being incubated at 25°C for 5 days, these strains were stored at 4°C for future use. The cultures were grown in liquid Potato Dextrose Broth (PDB) medium (20 g L–1; Hi-media) at 120 rpm in an orbital shaker at 25°C for 7 days. The mycelium was harvested by filtration and washed for three times with sterilized double-distilled water.
DNA extraction: Total DNA was extracted from modified CTAB method (Murray and Thompson, 1980; Dubey and Singh, 2008). The DNA pellets were re-suspended in 200 μL TE buffer (10 mmol L–1 tris hydrochloric acid and 1 mmol L–1 sodium EDTA, pH 8) and analyzed by electrophoresis at 5 V cm–1, on a 1.0% agarose gel in 0.5×TBE buffer (45 mM Tris-borate and 1 mM EDTA at pH 8.0) stained with 0.5 μg mL–1 ethidium bromide. Quality and quantity of DNA were estimated by spectrophotometer and stored at -20°C in small aliquots.
SRAP markers and polymerase chain reaction: The SRAP technique is a PCR-based marker system employing a combination of two primers, a forward primer of 17 bases and a reverse primer of 18 bases, which consisted of preferential amplification of Open Reading Frames (ORFs). The primers were composed of a core sequence of 13-14 bases, where the first 10 or 11 bases starting at the 5' end were ‘Filler’ sequences of no specific constitution followed by the sequence CCGG in the forward primer and AATT in the reverse primer. Three selective bases were added to the 3' ends of the primers. A variation in these three selective nucleotides generated a set of primers sharing the same core sequence. The forward primer preferentially amplified exonic regions. The reverse primer preferentially amplified intronic regions. The observed polymorphism fundamentally originated in the variation of the length of these introns, promoters and spacers, both among individuals and among species (Li and Quiros, 2001). In the present study, 30 different combinations of SRAP primers (Sun et al., 2006) were employed using five forward primers and six reverse primers (Table 2).
The polymerase chain reaction mixture (25 μL) consisted of 50 ng template DNA, 1.0 U Taq polymerase, 2.0 mM of MgCl2, 0.4 mM of each of the dNTPs and 7.5 pmol of each primer in 1×reaction buffer (Bangalore Genei, India). For amplification, first five cycles were run at 94°C for 1 min, 35°C for 1 min and 72°C for 1 min, for denaturing, annealing and extension, respectively. Then the annealing temperature was raised to 50°C for another 35 cycles. Amplification products were analyzed by electrophoresis in 2% agarose gel in 1×TAE buffer at 110 V for 1 h. Gels were stained with ethidium bromide (1 μg mL–1) and observed under UV light (Bio RadTM gel doc system). The primers that gave reproducible and scorable amplifications were used for the analysis.
Scoring and data analysis: Relatedness among 89 isolates of R. solani was estimated by means of scorable DNA bands amplified from different SRAP markers. Each band was considered as character and was scored as either present (coded as 1) or absent (coded as 0). Bands were reproducible and scored in identical positions for each time.
| Table 2: | Sequences of the forward and reverse SRAP primer combinations used in this study |
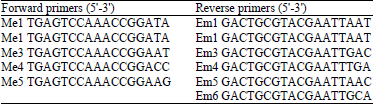 | |
Cluster analysis employing Unweighted Pair Group Method with an Arithmetic Average (UPGMA) algorithm was performed using NTSYS-PC (v. 2.01) to produce the different dendrogram (Rohlf, 1998). Bootstrap analysis was also performed using Winboot software for all the SRAP primers which amplified the R. solani isolates.
RESULTS
A total of 30 different combinations of SRAP primers were employed using five forward and 6 reverse primers (Table 2). Among all the combinations only 16 combinations of the primers revealed the polymorphisms between the isolates of R. solani and no monomorphic band was observed (Table 3). The primers amplified all the isolates of R. solani and the isolates were highly variable in respect of their banding profiles. The number of fragments amplified using each primer combination was ranging from 9-14 with an average of 11 polymorphic bands per combination of primers and band size of 0.1-1 kb. All 172 bands generated with 16 primers combinations were polymorphic showing high range of variability with 100% polymorphism (Table 3). The primer combination Me1/Em1 and Me1/Em4 showed maximum polymorphism and a representative gel picture of primer Me2/Em4 is given (Fig. 1).
Eighty nine isolates of the pathogen representing 7 Anastomosis Groups (AGs) were grouped into 7 major clusters at 35% similarity coefficient using UPGMA analysis (Fig. 2).
| Table 3: | Sequence related amplified polymorphism primer combinations and their polymorphism obtained in this study |
 | |
| NA: Not amplified | |
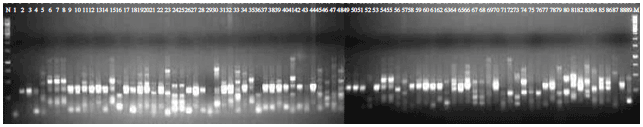 | |
| Fig. 1: | DNA profile generated by SRAP primer combination of Me2 and Em4. M marker-1 kb. Lanes 1-12: AG 1, 13-20: AG 2-2, 21-23: AG 2-2LP, 24-41: AG 2-3, 42-59: AG 3, 60-66: AG 4, 67-78: AG 5 and 79-89: Unknown AG isolates of Rhizoctonia solani |
The first cluster had three isolates of AG 1 each from Andhra Pradesh (RAPS3; AG1), Haryana (RHRW16; AG1) and Jammu and Kashmir (RJKM18; AG1). The second cluster had 28 isolates representing 5 AGs, namely, AG1, AG2-2, AG2-2LP, AG2-3 and AG3 originated from Uttar Pradesh, Haryana, Delhi, Jharkhand, Uttarakhand, Gujarat, Jammu and Kashmir, Assam, Madhya Pradesh, Bihar and Maharashtra. The third clusters had 6 isolates from 3 AGs, namely, AG1, AG2-2 and AG2-3 originated from 4 states, namely Madhya Pradesh, Haryana, Gujarat, Maharashtra and Tamil Nadu. Thirteen isolates originated from Andhra Pradesh, Assam, Maharashtra, Punjab, Haryana, Rajasthan, Uttarakhand, Delhi, Kerala and Gujarat representing 3 AGs, namely, AG2-3, AG3 and AG5 along with 2 isolates of undetermined AG were grouped into the fourth cluster. The fifth cluster had 29 isolates from 3 AGs, namely, AG3, AG4 and AG5 along with 9 isolates of undetermined AG originated from 15 states representing almost all parts of the country. Nine isolates from 6 states belongs to 4 AGs, namely, AG3, AG2-3, AG3 and AG5 were grouped in the sixth cluster. The seventh clusters had only one isolate from Madhya Pradesh (RMPM23; AG4).
The dendrogram derived from the data on amplified products obtained with 16 primer sets by using bootstrap analysis showed different levels of genetic similarity among the populations of R. solani. At 28% genetic similarity, the isolates were grouped into five major clusters. The first major cluster consisting of 81 isolates was further sub-divided into 13 sub-clusters. Majority of the sub-clusters had AG specific isolates originated from different parts of the country along with their respective AG tester isolates. The second and third clusters had only 2 isolates in each representing AG2-3 and AG3, respectively. The fourth cluster had 3 isolates belonging to AG3 and AG5 whereas, the fifth cluster had only one isolate of AG4 (Fig. 3). About 95% isolates showing similar sub-clustering by using both types of analysis.
DISCUSSION
Genetic diversity analysis of R. solani populations with Sequence Related Amplified Polymorphism (SRAP) markers using UPGMA and bootstrap indicated that the isolates were highly variable in their genetic makeup. More or less similar clustering patterns were obtained in both the analysis. The AG specific sub-clustering was common in both the analysis with a few exceptions might be due to genetic variation in the isolates of the same AG. The groups generated by UPGMA and bootstrap analyses were not corresponding to the place of origin, agro-ecological region and host of the isolates. Thus, the polymorphism of R. solani was very rich that may relate with their diverse growing environments, wide host range and genetic background. In the present study, first time attempt was made to explore the possibility of SRAP markers for diversity analysis of fungal plant pathogens. Earlier to this, the SRAP was used for diversity analysis to resolve the taxonomy of plants and other fungi. Sun et al. (2006) successfully demonstrated the utility of SRAP markers for taxonomic analysis of 31 different Ganoderma strains. The results revealed the genetic diversity of Ganoderma strains and their correlation with geographic environments. It has also been suggested that the SRAP markers could be used in the taxonomic analysis of fungi.
Ma et al. (2010) studied the genetic diversity of Tricholoma matsutake using SRAP technique. A total of 129 strains from 13 geographical locations in Northeastern China were amplified by using 12 primer pairs. Abundant genetic variation was detected within the individual populations. The analyses showed significant positive correlation between genetic distance and geographical distance and no correlation between genetic distance and altitudinal differences among the populations could be established. In the present study also, high level genetic variability among the populations of R. solani was observed. The present study also supported by Chang et al. (2012), who observed the genetic variations among and within native Saccharum spontaneum populations collected from Sichuan, China. Ren et al. (2012) also revealed a large genetic variation in 20 strains of endophytic fungi using SRAP markers. They concluded that the SRAP technology is more efficient than traditional morphological identification. It was found that SRAP markers could more really reflect the genetic diversity of endophytic fungal strains and also could be used as a method for identification of endophytic fungi.
 | |
| Fig. 2: | Dendrogram derived from combined analysis of 16 SRAP primers combinations for 89 Rhizoctonia solani isolates using UPGMA. The bottom scale represents the percentage of Jaccard’s similarity coefficient |
 | |
| Fig. 3: | Dendrogram derived from combined analysis of 16 SRAP primers combinations for 89 Rhizoctonia solani isolates using boot step analysis. Numbers at the forks show the percentage of time the group consisting of the isolates which are to the right of that fork occurred |
Zhang et al. (2014) analyzed the genetic diversity of endangered Polyporus umbellatus using SRAP and clearly indicated the suitability of markers for diversity analysis. Dubey et al. (2012) analyzed the genetic diversity of similar set of R. solani isolates with different molecular markers namely, Universal Rice Primers (URPs), Random Amplified Polymorphic DNA (RAPD) and Inter Simple Sequence Repeats (ISSR) markers. The majority of the isolates representing various AGs were grouped together into different sub-clusters. At genetic level, the isolates did not correspond to agro-ecological regions or states and crops of the origin because of the wide host range and adopt ability of the pathogen under different environmental conditions. The present finding is accordance with these results, indicating the suitability of SRAP for diversity analysis.
SRAP markers were also used to analyzed the genetic diversity of different fungal pathogens (Pasquali et al., 2010; Que et al., 2012; Xu et al., 2014; Dinolfo et al., 2015). Pasquali et al. (2010) first time used SRAP technique to analyse the molecular variability of fungal pathogen Puccinia striiformis f. sp. tritici and identified the polymorphisms among the isolates. Que et al. (2012) assessed molecular variation among Sporisorium scitamineum isolates causing sugarcane smut by SRAP markers. The molecular variation of S. scitamineum was associated with geographic origin and there was no evidence of co-evolution between sugarcane and the pathogen. Xu et al. (2014) observed considerable genetic variation among the isolates of S. scitamineum and the environmental heterogeneity has played an important role for high degree of variation. They reported that the genetic difference in the population of the pathogen depends to a large extent on the heterogeneity of their habitats and long-term adaptations of pathogens to their ecological environments. In the present study also several AG specific isolates from the same place clustered together clearly indicates the variability in the population of R. solani due to adoption and variation in the host in particular area. The present results are also in accordance with the observations made by Dinolfo et al. (2015) used SRAP markers for Fusarium poae genetic variability. The molecular analysis showed high intraspecific variability within F. poae isolates and a partial relationship was revealed between variability and the host/geographic origin.
The present study clearly indicated that the SRAP markers are suitable for genetic diversity analysis of R. solani. The bootstrap analysis clearly showed that the R. solani population in India broadly originated from similar ancestors as out of 89 isolates (including 7 tester isolates), 81 grouped in a single cluster with 28% genetic similarity. Although, AG wise sub-clustering was obtained. The variation in the population might be due to diversification of crops and variation in the environmental conditions in different agro-ecological regions of the country. More or less similar, AG specific sub-grouping pattern was obtained with UPGMA analysis.
CONCLUSION
The populations of Rhizoctonia solani belonging to 7 Anastomosis Groups (AGs) isolated from different pulse crops representing 21 states from 16 agro-ecological regions of India showed high level of variability during genetic diversity analysis using SRAP markers. The markers provided 100% polymorphism showed suitability for genetic diversity analysis of phytopathogenic fungi. The genetic groups did not correspond to origin of the crops and agro-ecological regions might be due to wide range of the hosts and adaptability of the pathogen under divers environmental conditions present in various parts of the country but partially corresponding to the Anastomosis Groups (AGs) of the pathogen. The present study deals first attempts on the use of SRAP markers for genetic diversity study of plant pathogenic fungus R. solani.
ACKNOWLEDGMENT
Authors are thankful to the funding agency ICAR-NFBSFARA for financial support.
REFERENCES
- Chang, D., F.Y. Yang, J.J. Yan, Y.Q. Wu and S.Q. Bai et al., 2012. SRAP analysis of genetic diversity of nine native populations of wild sugarcane, Saccharum spontaneum, from Sichuan, China. Genet. Mol. Res., 11: 1245-1253.
Direct Link - Dinolfo, M.I., E. Castanares and S.A. Stenglein, 2015. SRAP as an informative molecular marker to study the Fusarium poae genetic variability. J. Phytopathol., 163: 657-663.
CrossRefDirect Link - Dubey, S.C., 2003. Integrated management of web blight of urd/mung bean by bio-seed treatment. Indian Phytopathol., 56: 34-38.
Direct Link - Dubey, S.C. and S.R. Singh, 2008. Virulence analysis and oligonucleotide fingerprinting to detect diversity among Indian isolates of Fusarium oxysporum f. sp. ciceris causing chickpea wilt. Mycopathologia, 165: 389-406.
CrossRefDirect Link - Dubey, S.C., A. Tripathi and B.K. Upadhyay, 2012. Molecular diversity analysis of Rhizoctonia solani isolates infecting various pulse crops in different agro-ecological regions of India. Folia Microbiologica, 57: 513-524.
CrossRefDirect Link - Dubey, S.C., A. Tripathi, B.K. Upadhyay and U.K. Deka, 2014. Diversity of Rhizoctonia solani associated with pulse crops in different agro-ecological regions of India. World J. Microbiol. Biotechnol., 30: 1699-1715.
CrossRefDirect Link - Ferriol, M., B. Pico and F. Nuez, 2003. Genetic diversity of a germplasm collection of Cucurbita pepo using SRAP and AFLP markers. Theoret. Applied Genet., 107: 271-282.
CrossRefDirect Link - Li, G., M. Gao, B. Yang and C.F. Quiros, 2003. Gene for gene alignment between the Brassica and Arabidopsis genomes by direct transcriptome mapping. Theoret. Applied Genet., 107: 168-180.
CrossRefDirect Link - Li, G. and C.F. Quiros, 2001. Sequence-Related Amplified Polymorphism (SRAP), a new marker system based on a simple PCR reaction: its application to mapping and gene tagging in Brassica. Theoret. Applied Genet., 103: 455-461.
CrossRefDirect Link - Ma, D., G. Yang, L. Mu and Y. Song, 2010. Application of SRAP in the genetic diversity of Tricholoma matsutake in Northeastern China. Afr. J. Biotechnol., 9: 6244-6250.
Direct Link - Murray, M.G. and W.F. Thompson, 1980. Rapid isolation of high molecular weight plant DNA. Nucl. Acids Res., 8: 4321-4326.
CrossRefPubMedDirect Link - Ogoshi, A., 1987. Ecology and pathogenicity of anastomosis and intraspecific groups of Rhizoctonia solani Kuhn. Annu. Rev. Phytopathol., 25: 125-143.
CrossRefDirect Link - Pasquali, M., H. Komjati, D. Lee and R. Bayles, 2010. SRAP technique efficiently generates polymorphisms in Puccinia striiformis isolates. J. Phytopathol., 158: 708-711.
CrossRefDirect Link - Que, Y., L. Xu, J. Lin, R. Chen and M.P. Grisham, 2012. Molecular variation of Sporisorium scitamineum in Mainland China revealed by RAPD and SRAP markers. Plant Dis., 96: 1519-1525.
CrossRefDirect Link - Ren, N., J. Liu, D. Yang, J. Chen, M. Luan and J. Hong, 2012. Sequence-Related Amplified Polymorphism (SRAP) marker as a new method for identification of endophytic fungi from Taxus. World J. Microbiol. Biotechnol., 28: 215-221.
CrossRefDirect Link - Rohlf, F., 1998. NTSYS-pc: Numerical Taxonomy and Multivariate Analysis System. Version 2.2, Exeter Publications Setauket, New York.
Direct Link - Sun, S.J., W. Gao, S.Q. Lin, J. Zhu, B.G. Xie and Z.B. Lin, 2006. Analysis of genetic diversity in Ganoderma population with a novel molecular marker SRAP. Applied Microbiol. Biotechnol., 72: 537-543.
CrossRefDirect Link - Xu, L., Y. Lu, Q. You, X. Liu, M.P. Grisham, Y. Pan and Y. Que, 2014. Biogeographical variation and population genetic structure of Sporisorium scitamineum in mainland China: Insights from ISSR and SP-SRAP markers. Sci. World J., Vol. 2014.
CrossRef - Zhang, Y., Y. Qin, Z. Wang, L. Guo, X. Chen, Z. Liang and H. Guo, 2014. DNA isolation and optimization of SRAP-PCR condition for endangered Polyporus umbellatus. J. Med. Plants Res., 8: 133-137.
Direct Link - Lin, Z.X., X.L. Zhang and Y.C. Nie, 2004. [Evaluation of application of a new molecular marker SRAP on analysis of F2 segregation population and genetic diversity in cotton]. J. Genet. Genomics, 31: 622-626, (In Chinese).
PubMedDirect Link