
C. Anuradha
ICAR National Research Centre for Banana, Trichy, Tamil Nadu, India
R. Selvarajan
ICAR National Research Centre for Banana, Trichy, Tamil Nadu, India
S. Vasantha
ICAR Sugarcane Breeding Institute, Coimbatore, Tamil Nadu, India
G. S. Suresha
ICAR Sugarcane Breeding Institute, Coimbatore, Tamil Nadu, India
Plant Pathology Journal
Year: 2015 | Volume: 14 | Issue: 4 | Page No.: 212-222







ABSTRACT
Virus infection can result in the alteration of physiological, biochemical and metabolic processes within plants leading to symptom development. Banana bunchy top virus (BBTV) is one of the most destructive viral diseases in Tropical Asia, Pacific Indian Oceania (PIO) regions and Africa leading to 100% yield loss in banana and plantains. Though molecular characterization and their diversity were studied in depth in recent years, information on physiological and biochemical changes during banana-BBTV interaction is still not convincingly explained. Therefore, the present investigation was conducted to find out the quantifiable changes in physiological and biochemical parameters such as proteins, pigment and carbohydrate content, phenolic compounds, polyphenol oxidase (PPO), peroxidase (POX), ascorbate peroxidase (APX), guaiacol peroxidase (GPX), catalase (CAT) and superoxide dismutase (SOD) activities in leaves of banana cultivars Grand Nain (AAA) and Virupakshi (AAB). The amount of carbohydrate contents, phenolic compounds, PPO, POX, APX, GPX, CAT were significantly higher in BBTV infected leaves of both the cultivars over the healthy, whereas total protein content, pigments and SOD activity showed an opposite trend. Overall the results suggest that BBTV infection induces significant changes in enzyme levels leading to irreversible symptom development. Further studies would lead to identification of biochemical markers for studying plant-virus compatible and incompatible interactions.
PDF Abstract XML References Citation
Received: August 26, 2015;
Accepted: October 19, 2015;
Published: November 09, 2015
How to cite this article
C. Anuradha, R. Selvarajan, S. Vasantha and G. S. Suresha, 2015. Biochemical Characterization of Compatible Plant Virus Interaction: A Case Study with Bunchy Top Virus-Banana Host-Pathosystem. Plant Pathology Journal, 14: 212-222.
DOI: 10.3923/ppj.2015.212.222
URL: https://scialert.net/abstract/?doi=ppj.2015.212.222
DOI: 10.3923/ppj.2015.212.222
URL: https://scialert.net/abstract/?doi=ppj.2015.212.222
INTRODUCTION
Environmental stresses of both biotic and abiotic nature produce characteristic changes in physiology and metabolic processes of higher plants (Miteva et al., 2005). Among these stresses, infection by the pathogens causes substantial alterations of biochemical changes leading to harmful effects on plant health. Banana cultivation is subjected to many natural calamities including pest and pathogen attacks which constitute the major production problem. Among plant diseases, viral diseases may cause considerable loss on production by retarding plant growth and reducing yield. Four viral diseases viz., Banana Bunchy Top Disease (BBTD) caused by Banana bunchy top virus (BBTV), Bract Mosaic Disease (BBrMD) caused by Banana bract mosaic virus (BBrMV), Banana Streak Disease (BSD) caused by different species of Banana streak virus (BSV) and banana mosaic or infectious chlorosis caused by Cucumber mosaic virus (CMV) occur in most of the banana growing regions of the world. Among banana viral diseases, Banana Bunchy Top Disease (BBTD) is one of the most destructive viral diseases in Tropical Asia, Pacific Indian Oceania (PIO) regions and Africa. Grand Nain, Virupakshi (Hill banana), Robusta, Nendran, Rasthali, Poovan, Ney Poovan, Monthan and Red Banana are severely affected by BBTD. There is no resistance gene source available in germplasm of bananas and plantains (Shekhawat et al., 2012). The BBTV affected plants show intermittent dark green dots, dash, streaks of variable length on leaf sheath, midrib, leaf veins and petioles of infected plants. Leaves produced are progressively shorter, brittle in texture, narrow and gives the appearance of bunchyness at the top leading to 100% yield loss (FAO., 2009). Plants have evolved various pre-existing physical and chemical barriers, as well as inducible defense responses that interfere with pathogen colonization (Jones and Dangl, 2006; Zhao et al., 2008; Vanitha et al., 2009). However, this requires comprehensive studies and understanding of the adaptive mechanisms and responses to BBTV infection in banana. Because viruses cannot be cultured in vitro, our knowledge about their interactions with host and changes on physiology, biochemistry and molecular biology of the host is limited. Moreover, a comprehensive report regarding biochemical alterations in banana plants infected by BBTV is still not clear. Therefore, the present investigation was conducted to find the quantitative estimations of physiological and biochemical parameters such as protein, pigment and carbohydrate contents, phenolic compounds, polyphenol oxidase (PPO), peroxidase (POX), catalase (CAT), ascorbate peroxidise (APX), guaiacol peroxidise (GPX) and superoxide dismutase (SOD) activities, indicating their role in BBTV inoculated and non-inoculated plants of Virupakshi (hill banana), a unique flavoured elite dessert banana cultivar and Grand Nain, which belongs to a Cavendish sub group occupying approximately 50% of area in the world.
MATERIALS AND METHODS
Two cultivars of banana, Virupakshi (hill banana) and Grand Nain were used in the present investigation. Three month old suckers were planted in pots containing a mixture of sand, loam soil and compost (1:1:2). The pots were kept in insect-proof glass house. Virus vector aphids (fifteen adult or late instar aphids) were collected and transferred to healthy banana plants having four to five well developed leaves for inoculation access for 48 h. Inoculation access feeding was conducted at 25±0.5°C and 12 h light/dark photoperiod. At the completion of inoculation-access period all plants were sprayed with insecticide imidacloprid (0.1%) and kept in the greenhouse to monitor the symptom expression. All the plants showed typical BBTD symptoms after 20-30 days of inoculation. The un-inoculated plants (healthy plants) of each cultivar were maintained as control. Leaf samples were harvested from both controls and inoculated plants after symptom development to determine the enzyme activities.
Estimation of chlorophylls: Total chlorophyll, chlorophyll ‘a’ and chlorophyll ‘b’ contents of healthy and infected leaves were estimated as per the non-destructive DMSO method (Hiscox and Israelstam, 1979). The leaf discs (500 mg) were collected in test tubes, to this 10 mL of DMSO were added. The tubes were kept overnight in dark for 1-2 h. The absorbance was recorded at 663 and 645 nm in a spectrophotometer. Chlorophyll a, b and total chlorophyll were calculated by using following formulas:
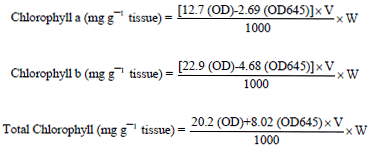 |
Where:
| OD | : | Optical density at respective nm |
| V | : | Final volume of chlorophyll extract |
| W | : | Fresh weight of the tissue extracted |
Estimation of total sugars and starch: The estimation of total sugars was done according to the method of DuBois et al. (1956) and total starch content by method of McCready et al. (1950). Briefly, 500 mg of healthy and infected leaves were taken, washed thoroughly with tap water followed by distilled water and blotted to dry in between filter paper folds. The midribs of leaf samples were removed, cut into bits and macerated with 5 mL of 80% ethanol. The macerates were transferred to centrifuge tubes and centrifuged at 5000 rpm for 15 min. The pellet was washed thrice with 80% ethanol. The supernatants were pooled and made upto known volume with 80% ethanol. The samples were heated in water bath at 85°C until the alcohol was completely lost from the samples. The supernatants were pooled and used for estimation of sugars. The pellet was subsequently used for extraction and estimation of starch.
For sugar determination, 20 mL of healthy and virus infected pooled supernatants were taken separately into the test tubes. One milliliter of distilled water and 4 mL of cold anthrone reagent were rapidly added to each tube, shaken well and incubated for 10 min on ice bath and cooled at room temperature. The blank was prepared by taking 1 mL of distilled water and 4 mL of cold anthrone reagent. The absorbance of the samples was read at 625 nm in a spectrophotometer. Amount of total sugars was estimated by using a standard curve prepared for D-glucose.
For starch determination, the pellet which was collected while preparing the extract for total sugar was solubilized in 5 mL of 52% perchloric acid (PCA) and boiled at 80°C for 10 min. The solution was filtered through glass wool. The filtrate was measured and made up to 10 mL with PCA. Twenty microliter of healthy and infected sample extracts were taken separately, added 3 mL of distilled water and 5 mL of anthrone reagent and incubated for 10 min in ice bath. The absorbance of the samples was read at 625 nm in a spectrophotometer. The amount of starch was calculated by using glucose standard curve.
Estimation of phenol: The phenol content was estimated using Folin-Ciocalteau reagent. 80% ethanol was used for extraction of phenols. One gram plant material was ground, divided into two 5 mL portions of 80% ethanol and centrifuged. The extracts were pooled and made up to 10 mL. Then, 0.1 mL of ethanol extract was evaporated on a water bath, to which 6 mL water was added and shaken well before addition of 0.5 mL Folin-Ciocalteau reagent. After 5 min, 2 mL of 20% sodium carbonate solution were added. After incubation for 30 min, absorbance at 660 nm was measured. Using pyrocatechol as standard, the phenol content in the leaf extract was calculated (Folin and Ciocalteu, 1927).
Preparation of enzyme extract: One gram of leaf sample was homogenized at 4°C in 1 mL of extraction buffer [50 mM potassium phosphate buffer (pH 7.0), 1% Triton X-100 and 7 mM 2-mercaptoethanol] with mortar and pestle. The homogenate was then centrifuged at 12000 rpm for 20 min at 4°C and the supernatant was used as the crude extract for the estimation of POX, PPO, catalase, APX, GPX and SOD activities.
Enzyme assays: Peroxidase activity was assessed following the oxidation of O-dianisidine using the method of Malic and Singh (1980). For the assay 3.5 mL of phosphate buffer (pH-6.5) were taken in a clean dry cuvette and to it 0.2 mL of enzyme extract and 0.1 mL of freshly prepared O-dianisidine solution was added. Then 0.2 mL of 0.2M H2O2 were added and immediately absorbance of the reaction mixture was read at 430 nm at every 30 sec interval upto 3 min.
Polyphenol oxidase activity was measured according to the method described by Ngadze et al. (2012).
Catalase activity was assayed by measuring the rate of disappearance of H2O2 using the method of Maehly and Chance (1959). The reaction mixture contained 2.5 mL of 50 mM phosphate buffer (pH 7.4), 0.1 mL of 1% H2O2 and 50 μL of enzyme extract diluted to keep measurements within the linear range of the analysis. The decrease in H2O2 was followed as a decline in absorbance at 240 nm.
Ascorbate peroxidase activity was determined according to the method of Chen and Asada (1989) with minor modification. The 1 mL reaction mixture was composed of 50 mM phosphate buffer (pH 7.0) containing 0.1 mM EDTA, 0.5 mM ascorbate, 1.54 mM H2O2 and 50 μL of enzyme extract. The oxidation of ascorbate was followed by the decrease in the absorbance at 240 nm.
Guaiacol peroxidase activity was determined according to Upadhyaya et al. (1985). The reaction mixture contained 2.5 mL of 50 mM phosphate buffer (pH 6.1), 1 mL of 1% H2O2, 1 mL of 1% guaiacol and 20 μL of enzyme extract. The increase in absorbance at 420 nm was observed for 1 min.
Superoxide dismutase activity was assayed by measuring its ability to inhibit the photochemical reduction of nitroblue tetrazolium (NBT) using the method of Dhindsa et al. (1981). The 3 mL reaction mixture contained 50 mM phosphate buffer (pH 7.8), 13 mM methionine, 75 M NBT, 2 M riboflavin, 0.1 mM EDTA and 50 μL of enzyme extract. Riboflavin was added last and the tubes were shaken and placed 30 cm below a light bank consisting of two 15 W fluorescent lamps for 10 min. The absorbance of the reaction mixture was read at 560 nm.
Estimation of total protein content: Total protein was estimated colorimetrically by using Bradford method (Bradford, 1976) recording absorbance at 595 nm. Bovine serum albumin was used as standard. Protein content in leaf samples was recorded as μg of protein per gram of leaf tissue.
Statistical analysis: All experiments were repeated twice with similar results. Data was subjected to an analysis of variance and means and standard errors calculated.
RESULTS AND DISCUSSION
The physiology of virus infected plant tissues is of prime interest to understand the processes involved in the development of symptoms. There are several reports in the literature indicating many changes in the physiology and biochemistry of host plants as a consequence of viral disease (Radwan et al., 2007). Many investigators (Matthews, 1970) in different host virus combinations have estimated and correlated the virus concentration to altered metabolic processes and symptom production and severity.
Chlorophyll: Leaf chlorophyll content can be directly related to stress physiology affecting the growth and yield. Virus infection induces changes in leaf pigmentation, hence the symptoms. The relative chlorophyll content of banana leaves was compared between BBTV infected and healthy plants. Drastic reduction of chlorophyll a, chlorophyll b and total chlorophyll was observed in infected Virupakshi and Grand Nain leaves (Fig. 1a-c). From the values of a/b ratio it can be deduced that chlorophyll b is more sensitive to viral infection than chlorophyll a. Reduced chlorophyll content has been reported in virus infected plants (Sinha and Srivatsava, 2010) and is attributed to stimulation of cell enzymes like chlorophyllase that degrades chlorophyll (Goodman et al., 1967), or it may be the effect of virus on pigment synthesis (Porter, 1959; Sadasivan, 1963; Granick and Beale, 1983; Balachandran et al., 1997) and disturbed physiological processes like photosynthesis and utilization of plastid proteins or their precursors for the synthesis of virus protein (Fraser, 1987).
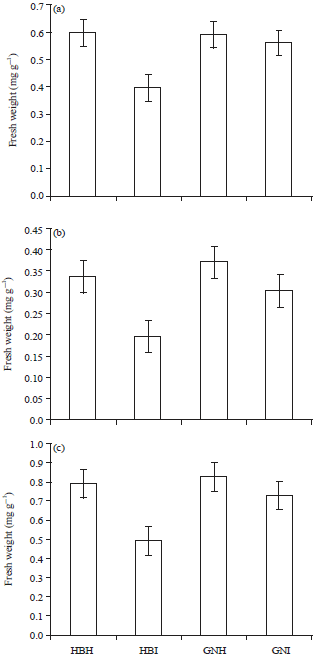 | |
| Fig. 1(a-c): | Chlorophyll content (a) Chlorophyll a (b) Chlorophyll b and (c) Total chlorophyll of healthy and infected Virupakshi (HB) and Grand Nain (GN) plants |
Other reasons for the decrease in chlorophyll content following virus infection may be accumulation of carbohydrates in the leaves (Watson, 1955). As in case of many different virus-host interactions, altered ratio of chlorophylls due to BBTV was observed probably affecting the photosynthetic efficiency (Endo et al., 2001). According to Sheffield (1933), the virus of tomato aucuba mosaic did not affect the chlorophyll in older leaves but prevented the formation of plastids in young growing leaves, on the other hand cucumber mosaic in tomato produced stripe chlorosis showing destruction of already formed chlorophyll.
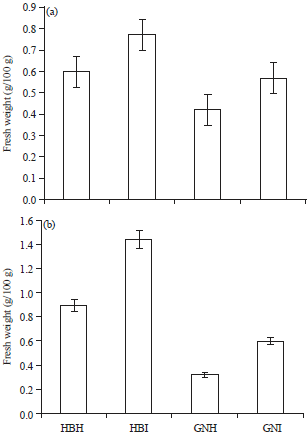 | |
| Fig. 2(a-b): | Carbohydrate content (a) Total sugars and (b) Starch of healthy and infected Virupakshi (HB) and Grand Nain (GN) plants |
Subsequent studies with improved techniques have shown that the chlorophyll 'a' associated with the reaction centre of photosystem II (i.e., CPa complex) was reduced to a greater extent than the antenna chlorophyll of light harvesting system (i.e., LHCPY, 1, 2, 3 and II d complexes). Changes in pigments are often considered to be the secondary effects on the host plant since many viruses appear to multiplying and accumulate in the cytoplasm of the cell. Although, these changes appear to be secondary as far as synthesis is considered, they are important for disease induction or resistance process, considering the plant as a whole (Crosbie and Matthews, 1974).
Carbohydrates: Influence of plant pathogenic viruses on the carbohydrate metabolism of the infected host is very important with regard to economic damage caused to host. Some viruses appear to have little effect on carbohydrates in the leaves, while others may alter both their rate of synthesis and rate of translocation (Gaddam et al., 2012). In the present investigation, the total sugars and starch were high in infected plants compared to healthy (Fig. 2a-b). Regarding viruses, carbohydrates appear to have a role in plant defences and it has been known since the 1930s that accumulation of starch precedes the presence of virus symptoms (Holmes,1931) and for example, marrow plants infected with Zucchini yellow mosaic virus (ZYMV) (Blua et al., 1994), mosaic infected acid
lime (Gaddam et al., 2012), ZYMV of Cucurbita pepo (Radwan et al., 2007), Sunflower mosaic virus (Bhavani et al., 1998), Papaya meleira virus (Buss et al., 2011) and Yellow virus infected sugar beet (Watson, 1955) show increased sucrose content. A link of carbohydrates with defences against oxidative stress has been observed in other studies (Sulmon et al., 2004; Loreti et al., 2005) and carbohydrates are essential in the production of many anti-oxidant defences (Couee et al., 2006).
Our results suggest that sugar increases during BBTV infection may modulate photoinhibitory processes which probably induce the symptoms. Virus infection also induces singlet oxygen generation resulting from the photoinhibitory process. Singlet oxygen (O21) is a potent agent of oxidative damage that could also contribute to chlorotic symptom development (Fryer et al., 2002). The BBTV infection could in turn increases ROS generation in other cellular compartments (Diaz-Vivancos et al., 2008; Song et al., 2009) in association with respiratory and photorespiratory processes. These alternative ROS sources might contribute to the development of chlorotic symptoms.
Total protein: Involvement of protein components in plant diseases resistance has been documented in plant pathogenic interactions (Tornero et al., 2002; De Carvalho et al., 2006). Usually, infected plants show a high protein content, which could be due to both the activation of the host defense mechanism and the pathogen attack mechanism (Agrios, 1997). However, protein content was found to decrease significantly in the BBTV infected plants of both the cultivars (Fig. 3a). A possible explanation may be that the disease might have caused denaturation or break down of proteins as well as polypeptide chains and bound amino acids. Similar results have also been reported in Mesta infected with yellow vein mosaic disease (Chatterjee and Ghose, 2008) and in cotton with CLCuBuV (Siddique et al., 2014).
Total phenol: Upon infection by pathogens, the host phenolic compounds may increase and contribute to enhance the mechanical strength of host cell walls by the synthesis of lignin and suberin that are involved in the formation of physical barriers that can block the spread of pathogens (Ngadze et al., 2012; Singh et al., 2014). In the present study, the amount of total phenol was significantly higher in virus infected leaves (Fig. 3b) and the increased quantity of phenols might be attributed to a defence mechanism (Rai et al., 2010). The resistance to disease caused by pathogen was attributed to the presence of high amount of phenol (Jain and Yadav, 2003; Kushwaha and Narain, 2005; Parashar and Lodha, 2007; Meena et al., 2008; Rai et al., 2010; Siddique et al., 2014). Hence, the increased quantity of phenolics in the infected plant of the banana may be contributing to the resistance against the infection of viral pathogen. Increased levels of phenolics also suggest an acceleration of phenol synthesizing pathway following pathogen infection.
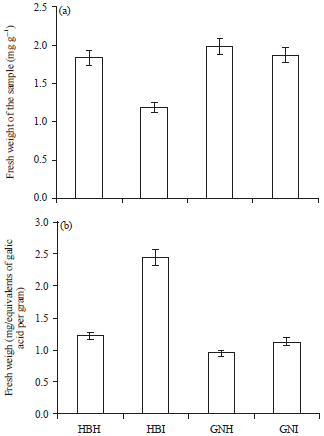 | |
| Fig. 3(a-b): | (a) Total protein and (b) Total phenol content of healthy and infected Virupakshi (HB) and Grand Nain (GN) plants |
Peroxidase: Peroxidase (POX) is one of the first enzymes responding and providing fast defense against plant pathogens (Sulman et al., 2001). The POXs are involved in the lignification, suberification, polymerization of hydroxy-proline-rich glycoproteins, regulation of cell wall elongation, wound healing and resistance against pathogens in plants (Hammond-Kosack and Jones, 1996; Yoshida et al., 2003; Maksimov et al., 2014). The POX activity was significantly higher in BBTV infected plants of both cultivars, as compared to healthy (Fig. 4a). This result is in accordance with the results of Tobacco mosaic virus infected tobacco (Lagrimini and Rothstein,1987), plum pox virus infected peaches, apricots (Diaz-Vivancos et al., 2006), Beans yellow mosaic virus infected beans (Radwan et al., 2010), Potato virus Y infected potato (Milavec et al., 2001), tomato and bell pepper infected with Tobacco mosaic virus and Tomato mosaic tobamo virus (Madhusudhan et al., 2009), Cucumber mosaic virus and Zucchini yellow mosaic virus infected Cucumis sativus and Cucurbita pepo plants (Riedle-Bauer, 2000), Tobacco mosaic virus infected tobacco plants (Kiraly et al., 2002), Tomato yellow leaf curl virus infected tomato plants (Dieng et al., 2011), Banana bunchy top virus infected cultivars of banana (Devanathan et al., 2005), geminivirus infected Capsicum annum (Meena et al., 2008) and cotton with CLCuBuV (Siddique et al., 2014).
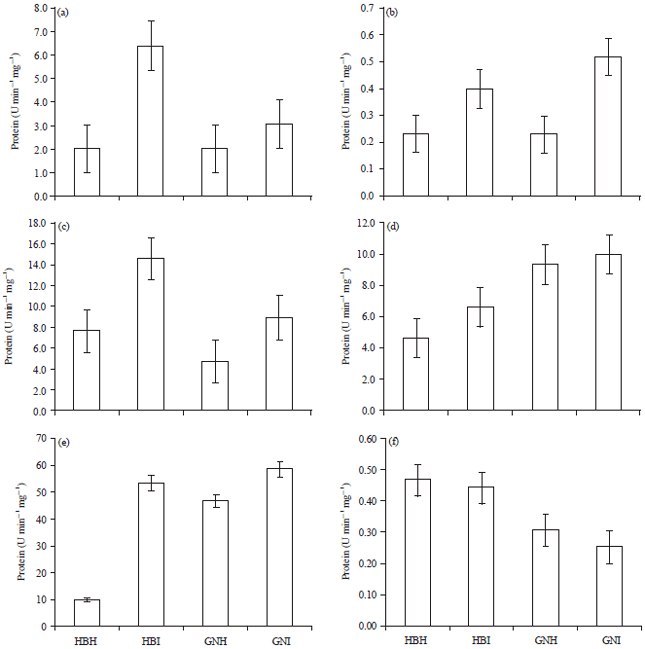 | |
| Fig. 4(a-f): | Changes in enzyme activities of (a) POX, (b) PPO, (c) CAT, (d) APX, (e), GPX (E) and (f) SOD in healthy and BBTV infected Virupakshi (HB) and Grand Nain (GN) plants |
Virus infection appears to stimulate POX activity in all hosts in which necrotic or chlorotic symptoms are induced, the degree of stimulation correlating with severity of symptoms (Wood, 1990). The POX is known to be involved in the Active Oxygen Species (AOS) mechanisms. The AOX accumulation causes oxidative damage through actions such as lipid peroxidation and membrane destruction. The AOX is proposed to be responsible for chlorophyll degradation and POX levels increase during senescence (Kuroda et al., 1990). So, there is a correlation between the decrease in chlorophyll content and increase in POX activity in the green leaves infected with virus (Milavec et al., 2001). Also, POX is known to catalyze the final polymerization step of lignin synthesis and is directly associated with the increased ability of systemically protected tissues to lignify (Chittoor et al., 1999). Therefore, higher activity of POX leads to lignification process which is considered as a resistance mechanism against pathogen attack. By way of oxidation of indole-3-acetic acid, upregulated peroxidases might also be responsible for growth reductions and malformations in virus-infected plants (Riedle-Bauer, 2000).
Polyphenol oxidase (PPO): Polyphenol oxidase (PPO) is important in the initial stage of plant defense where membrane damage causes release of phenols such as chlorogenic acid. The PPO catalyzes the oxidation of phenolics to free radicals that can react with biological molecules, thus creating an unfavorable environment for pathogen development (Jockusch, 1966; Mohamed et al., 2012). The PPO activity was found to increase in leaves of BBTV-infected plants of tested cultivars (Fig. 4b). Total soluble phenols together with PPO play a role in resistance to viral pathogens (Thipathi and Verma, 1975; Kumar et al., 1991; Li and Steffens, 2002; Ngadze et al., 2012).
Catalase: Catalase is an oxygen-scavenging enzyme that plays the role of specific peroxidative, protecting cells from the toxic effects of substrates (H2O2) during development, which are otherwise lethal (Choodamani et al., 2009; Patel et al., 2011; Hameed and Iqbal, 2014). The CAT activity was found to increase in leaves of BBTV-inoculated plants of both the tested cultivars (Fig. 4c). Similar increase in foliar CAT activity was observed in leaves of Arachis hypogaea infected with peanut mottle virus (Kobeasy et al., 2011), cotton leaves inoculated with Cotton leaf curl burewala virus (Siddique et al., 2014) and in Hibiscus cannabinus infected with Yellow vein mosaic virus (Sarkar et al., 2010).
Ascorbate peroxidase (APX): Ascorbate peroxidase (APX) is thought to play the most essential role in scavenging ROS and protecting cells in higher plants, algae and other organisms. The APX activity was significantly higher in BBTV infected plants of both cultivars (Fig. 4d) probably as an antioxidant response triggered by the increasing presence of H2O2 within cells. The increase in APX activity in banana cultivars infected with BBTV was similar to that reported for Hibiscus cannabinus infected with begomovirus (Sarkar et al., 2010), Nicotiana benthamiana infected with Pepper mild mottle virus (Hakmaoui et al., 2012) and sunflower infected with sunflower chlorotic mottle virus (Rodriguez et al., 2010). It was suggested that the over-production of APX increased the Peroxidase activity which strengthens the ROS scavenging system and leads to oxidative stress tolerance. It has been reported that in a compatible response between barley and powdery mildew the cytosolic isoenzyme of APX is up-regulated in both epidermal and mesophyll cells. In these cells, that are not able to trigger a response to stop pathogens, the APX increase limits the propagation of oxidative processes allowing cells to maintain their viability, a condition required for the penetration of biotrophic powdery mildew in plant tissues (Burhenne and Gregersen, 2000). This up-regulation of APX confirms previous results reporting an increase in APX activity during successful infection of barley leaves by biotrophic compatible pathogens (El-Zahaby et al., 1995; Vanacker et al., 1998; Kuzniak and Sklodowska, 1999) and has also been reported to occur in leaves of susceptible apricot infected by Plum pox virus (Hernandez et al., 2001).
Guaiacol peroxidase: Guaiacol peroxidase (GPX) is an important group from peroxidases, which oxidize guaiacol (omethoxyphenol) as a commonly used reducing substrate. Guaiacol peroxidase (GPX) activity was higher in BBTV infected cultivars (Fig. 4e). The GPX decomposes Indole-3-Acetic Acid (IAA) and has a role in the biosynthesis of lignin and defence against biotic stresses by consuming H2O2.
Superoxide dismutase: Superoxide dismutase is one of the most important scavenging enzymes and catalyzes the dismutation of superoxide radicals to active oxygen species hydrogen peroxide (Ehsani-Moghaddam et al., 2006; Hameed and Iqbal, 2014). The SOD activity was significantly higher in leaves of healthy plants of both the cultivars (Fig. 4f). This is in agreement with results of Hernandez et al. (2004), who has observed a significant decrease in SOD from Plum pox virus inoculated peaches, similarly Clarke et al. (2002) in WClMV-infected Phaseolus vulgaris L., plants, Buonaurio and Montalbini (1993) in case of PVY infected tobacco plants, Ashfaq et al. (2010) in Urdbean leaf crikle virus (ULCV) infected black gram and Zhuang et al. (1993) in a resistant soybean infected with Soybean mosaic virus. Higher activity of SOD could be a strategy of the plant to restrict virus colonization, because the excess ROS can be removed (Govrin and Levine, 2000; Ehsani-Moghaddam et al., 2006) and vice versa when SOD was lower resulting in oxidative stress.
CONCLUSION
All the physiological and biochemical parameters analyzed in banana leaves showed significant changes in response to BBTV infection. Reduced chlorophyll content with increased carbohydrate content may lead to chlorotic/morse code symptom development in BBTV infected banana plants. Early and elevated expression of defence enzymes is an important feature of plant resistance to pathogens. It has been suggested that the higher activity of antioxidant enzymes can interrupt defence signals generated by ROS in several pathosystems and thus leads to compatible virus-plant interactions. Our findings indicate that there is a significant increase in total protein, phenolic compounds, POX, PPO, CAT, APX, GPX and a decrease in SOD activity which may play an active role in disease resistance against BBTV infection. Induced levels of phenolics with these enzymes may have correlation with resistance and these enzymes could be considered as biochemical markers for studying plant virus interaction in this case, BBTV in particular.
REFERENCES
- Agrios, G.N., 1997. Plant Pathology. 4th Edn., Academic Press, Cambridge, Massachusetts, ISBN-13: 9780120115648, Pages: 635.
Direct Link - Ashfaq, M., M.A. Khan, N. Javed, S.M. Mughal, M. Shahid and S.T. Sahi, 2010. Effect of urdbean leaf crinkle virus infection on total soluble protein and antioxidant enzymes in blackgram plants. Pak. J. Bot., 42: 447-454.
Direct Link - Balachandran, S., V.M. Hurry, S.E. Kelley, C.B. Osmond and S.A. Robinson et al., 1997. Concepts of plant biotic stress. Some insights into the stress physiology of virus-infected plants, from the perspective of photosynthesis. Physiologia Plantarum, 100: 203-213.
CrossRefDirect Link - Bhavani, U., K.V. Subbaiah, A.S. Rao and D.V.R.S. Gopal, 1998. Studies on mosaic disease of sunflower: Biochemical changes and growth parameters. Indian Phytopathol., 51: 357-358.
Direct Link - Blua, M.J., T.M. Perring and M.A. Madore, 1994. Plant virus-induced changes in aphid population development and temporal fluctuations in plant nutrients. J. Chem. Ecol., 20: 691-707.
CrossRefDirect Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Buonaurio, R. and P. Montalbini, 1993. Peroxidase, superoxide dismutase and catalase activities in tobacco plants protected against Erysiphe cichoracearum by a necrotic strain of potato virus Y. Rivista Patologia Vegetale Serie V, 3: 23-31.
Direct Link - Burhenne, K. and P.L. Gregersen, 2000. Up-regulation of the ascorbate-dependent antioxidative system in barley leaves during powdery mildew infection. Mol. Plant Pathol., 1: 303-314.
CrossRefDirect Link - Buss, D.S., G.B. Dias, M.P. Santos, J.A. Ventura and P.M.B. Fernandes, 2011. Oxidative stress defence response of Carica papaya challenged by nitric oxide, papaya meleira virus and Saccharomyces cerevisiae. Open Nitric Oxide J., 3: 55-64.
CrossRefDirect Link - De Carvalho, D., R.A. Ferreira, L.M. de Oliveira, A.F. de Oliveira and R.C.R. Gemaque, 2006. Proteins and isozymes electroforesis in seeds of Copaifera langsdorffii Desf. (leguminosae caesalpinioideae) artificially aged. Revista Arvore, 30: 19-24.
CrossRefDirect Link - Chatterjee, A. and S.K. Ghosh, 2008. Alterations in biochemical components in mesta plants infected with yellow vein mosaic disease. Braz. J. Plant Physiol., 20: 267-275.
CrossRefDirect Link - Chen, G.X. and K. Asada, 1989. Ascorbate peroxidase in tea leaves: Occurrence of two isozymes and the differences in their enzymatic and molecular properties. Plant Cell Physiol., 30: 987-998.
Direct Link - Choodamani, M.S., P. Hariprasad, M.K. Sateesh and S. Umesha, 2009. Involvement of catalase in bacterial blight disease development of rice caused by Xanthomonas oryzae pv. oryzae. Int. J. Pest Manage., 55: 121-127.
CrossRefDirect Link - Clarke, S.F., P.L. Guy, D.J. Buritt and P.E. Jameson, 2002. Changes in the activities of antioxidant enzymes in response to virus infection and hormone treatment. Physiologia Plantarum, 114: 157-164.
CrossRefPubMedDirect Link - Couee, I., C. Sulmon, G. Gouesbet and A. El Amrani, 2006. Involvement of soluble sugars in reactive oxygen species balance and responses to oxidative stress in plants. J. Exp. Bot., 57: 449-459.
CrossRefDirect Link - Crosbie, E.S. and R.E.F. Matthews, 1974. Effects of TYMV infection on leaf pigments in Brassica pekinensis Rupr. Physiol. Plant Pathol., 4: 379-387.
CrossRefDirect Link - Devanathan, M., M. Ramaiah, A.R. Sundar and M. Murugan, 2005. Changes of peroxidase and polyphenol oxidase in bunchy top nana virus infected and healthy cultivars of banana. Ann. Plant Physiol., 19: 114-116.
Direct Link - Dhindsa, R.S., P. Plumb-Dhindsa and T.A. Thorpe, 1981. Leaf senescence: Correlated with increased levels of membrane permeability and lipid peroxidation and decreased levels of superoxide dismutase and catalase. J. Exp. Bot., 32: 93-101.
CrossRefDirect Link - Diaz-Vivancos, P., M. Rubio, V. Mesonero, P.M. Periago, A.R. Barcelo, P. Martinez-Gomez and J.A. Hernandez, 2006. The apoplastic antioxidant system in Prunus: Response to long-term plum pox virus infection. J. Exp. Bot., 57: 3813-3824.
CrossRefDirect Link - Diaz-Vivancos, P., M.J. Clemente-Moreno, M. Rubio, E. Olmos, J.A. Garcia, P. Martinez-Gomez and J.A. Hernandez, 2008. Alteration in the chloroplastic metabolism leads to ROS accumulation in pea plants in response to plum pox virus. J. Exp. Bot., 59: 2147-2160.
CrossRefDirect Link - Dieng, H., T. Satho, A. Abu Hassan, A.T. Aziz and R.E. Morales et al., 2011. Peroxidase activity after viral infection and whitefly infestation in juvenile and mature leaves of Solanum lycopersicum. J. Phytopathol., 159: 707-712.
CrossRefDirect Link - DuBois, M., K.A. Gilles, J.K. Hamilton, P.A. Rebers and F. Smith, 1956. Colorimetric method for determination of sugars and related substances. Anal. Chem., 28: 350-356.
CrossRefDirect Link - Ehsani-Moghaddam, B., M.T. Charles, O. Carisse and S. Khanizadeh, 2006. Superoxide dismutase responses of strawberry cultivars to infection by Mycosphaerella fragariae. J. Plant Physiol., 163: 147-153.
CrossRefDirect Link - El-Zahaby, H.M., G. Gullner and Z. Kiraly, 1995. Effects of powdery mildew infection of barley on the ascorbate-glutathione cycle and other antioxidants in different host-pathogen interactions. Phytopathology, 85: 1225-1230.
CrossRefDirect Link - Folin, O. and V. Ciocalteu, 1927. On tyrosine and tryptophane determinations in proteins. J. Biol. Chem., 27: 627-650.
Direct Link - Fryer, M.J., K. Oxborough, P.M. Mullineaux and N.R. Baker, 2002. Imaging of photo‐oxidative stress responses in leaves. J. Exp. Bot., 53: 1249-1254.
CrossRefDirect Link - Gaddam, S.A., V.S. Kotakadi, M.N. Reddy and D.V.R. Saigopal, 2012. Antigenic relationships of citrus yellow mosaic virus by immunological methods. Asian J. Plant Sci. Res., 2: 566-569.
Direct Link - Govrin, E.M. and A. Levine, 2000. The hypersensitive response facilitates plant infection by the necrotrophic pathogen Botrytis cinerea. Curr. Biol., 10: 751-757.
CrossRefDirect Link - Granick, S. and S.I. Beale, 1983. Hemes, chlorophylls and related compounds: Biosynthesis and metabolic regulation. Adv. Enzymol. Relat. Areas Mol. Biol., 46: 33-203.
PubMed - Hakmaoui, A., M.L. Perez-Bueno, B. Garcia-Fontana, D. Camejo, A. Jimenez, F. Sevilla and M. Baron, 2012. Analysis of the antioxidant response of Nicotiana benthamiana to infection with two strains of Pepper mild mottle virus. J. Exp. Bot., 63: 5487-5496.
CrossRefDirect Link - Hameed, A. and N. Iqbal, 2014. Chemo-priming with mannose, mannitol and H2O2 mitigate drought stress in wheat. Cereal Res. Commun., 42: 450-462.
CrossRefDirect Link - Hammond-Kosack, K.E. and J.D. Jones, 1996. Resistance gene-dependent plant defense responses. Plant Cell, 8: 1773-1791.
CrossRefPubMedDirect Link - Hernandez, J.A., J.M. Talavera, P. Martinez-Gomez, F. Dicenta and F. Sevilla, 2001. Response of antioxidative enzymes to plum pox virus in two apricot cultivars. Physiologia Plantarum, 111: 313-321.
CrossRefDirect Link - Hernandez, J.A., M. Rubio, E. Olmos, A. Ros-Barcelo and P. Martinez-Gomez, 2004. Oxidative stress induced by long-term plum pox virus infection in peach (Prunus persica). Physiologia Plantarum, 122: 486-495.
CrossRefDirect Link - Hiscox, J.D. and G.F. Israelstam, 1979. A method for the extraction of chlorophyll from leaf tissue without maceration. Can. J. Bot., 57: 1332-1334.
CrossRefDirect Link - Jain, A. and H.S. Yadav, 2003. Biochemical constituents of finger millet genotypes associated with resistance to blast caused by Pyricularia grisea Sacc. Ann. Plant Protect. Sci., 11: 70-74.
Direct Link - Jones, J.D.G. and J.L. Dangl, 2006. The plant immune system. Nature, 444: 323-329.
CrossRefDirect Link - Kiraly, Z., B. Barna, A. Kecskes and J. Fodor, 2002. Down-regulation of antioxidative capacity in a transgenic tobacco which fails to develop acquired resistance to necrotization caused by TMV. Free Radic. Res., 36: 981-991.
CrossRefDirect Link - Kumar, A., V.S. Pundhir and K.C. Gupta, 1991. The role of phenols in potato tuber resistance against soft rot by Erwinia carotovora ssp. carotovora. Potato Res., 34: 9-16.
CrossRefDirect Link - Kuroda, M., T. Oaiawa and H. Imagawa, 1990. Changes in chloroplast peroxidase activities in relation to chlorophyll loss in barley leaf segments. Physiologia Plantarum, 80: 555-560.
CrossRefDirect Link - Kushwaha, K. and U. Narain, 2005. Biochemical changes in pigeon-pea leaves infested with Alternaria tenuissima. Ann. Plant Protect. Sci., 13: 415-417.
Direct Link - Kuzniak, E. and M. Sklodowska, 1999. The effect of Botrytis cinerea infection on ascorbate-glutathione cycle in tomato leaves. Plant Sci., 148: 69-76.
CrossRefDirect Link - Lagrimini, L.M. and S. Rothstein, 1987. Tissue specificity of tobacco peroxidase isozymes and their induction by wounding and tobacco mosaic virus infection. Plant Physiol., 84: 438-442.
PubMedDirect Link - Li, L. and J.C. Steffens, 2002. Overexpression of polyphenol oxidase in transgenic tomato plants results in enhanced bacterial disease resistance. Planta, 215: 239-247.
CrossRefDirect Link - Loreti, E., A. Poggi, G. Novi, A. Alpi and P. Perata, 2005. A genome-wide analysis of the effects of sucrose on gene expression in Arabidopsis seedlings under anoxia. Plant Physiol., 137: 1130-1138.
CrossRefPubMedDirect Link - Madhusudhan, K.N., B.M. Srikanta, M.D. Shylaja, H.S. Prakash and H.S. Shetty, 2009. Changes in Antioxidant Enzymes, Hydrogen Peroxide, Salicylic Acid and Oxidative Stress in Compatible and Incompatible Host-Tobamovirus Interaction. J. Plant Interact., 4: 157-166.
CrossRefDirect Link - Maksimov, I., N. Troshina, O. Surina and E. Cherepanova, 2014. Salicylic acid increases the defense reaction against bunt and smut pathogens in wheat calli. J. Plant Interact., 9: 306-314.
CrossRefDirect Link - McCready, R.M., J. Guggolz, V. Silviera and H.S. Owens, 1950. Determination of starch and amylose in vegetables. Anal. Chem., 22: 1156-1158.
CrossRefDirect Link - Milavec, M., M. Ravnikar and M. Kovac, 2001. Peroxidases and photosynthetic pigments in susceptible potato infected with potato virus YNTN. Plant Physiol. Biochem., 39: 891-898.
CrossRefDirect Link - Miteva, E., D. Hristova, V. Nenova and S. Maneva, 2005. Arsenic as a factor affecting virus infection in tomato plants: Changes in plant growth, peroxidase activity and chloroplast pigments. Scientia Hortic., 105: 343-358.
CrossRefDirect Link - Kobeasy, M.I., H.S. El-Beltagi, M.A. El-Shazly and E.A. Khattab, 2011. Induction of resistance in Arachis hypogaea L. against Peanut mottle virus by nitric oxide and salicylic acid. Physiol. Mol. Plant Pathol., 76: 112-118.
CrossRefDirect Link - Ngadze, E., D. Icishahayo, T.A. Coutinho and J.E. van der Waals, 2012. Role of polyphenol oxidase, peroxidase, phenylalanine ammonia lyase, chlorogenic acid and total soluble phenols in resistance of potatoes to soft rot. Plant Dis., 96: 186-192.
CrossRefDirect Link - Parashar, A. and P. Lodha, 2007. Phenolic estimation in Foeniculum vulgare infected with ramularia blight. Ann. Plant Protect. Sci., 15: 396-398.
Direct Link - Patel, S.J., R.B. Subramanian and Y.S. Jha, 2011. Biochemical and molecular studies of early blight disease in tomato. Phytoparasitica, 39: 269-283.
CrossRefDirect Link - Porter, C.A., 1959. Biochemistry of plant virus infection. Adv. Virus Res., 6: 75-91.
CrossRefDirect Link - Radwan, D.E.M., K.A. Fayez, S. Younis Mahmoud, A. Hamad and G. Lu, 2007. Physiological and metabolic changes of Cucurbita pepo leaves in response to zucchini yellow mosaic virus (ZYMV) infection and salicylic acid treatments. Plant Physiol. Biochem., 45: 480-489.
CrossRefDirect Link - Rai, V.P., N. Jaiswal, S.P. Singh, R. Kumar and A.B. Rai, 2010. Response of total phenol and peroxidase activity in chilli exposed to pepper leaf curl virus disease. Vegetable Sci., 37: 78-80.
Direct Link - Riedle-Bauer, M., 2000. Role of reactive oxygen species and antioxidant enzymes in systemic virus infections of plants. J. Phytopathol., 148: 297-302.
CrossRefDirect Link - Sarkar, T.S., U. Majumdar, A. Roy, D. Maiti and A.M. Goswamy et al., 2010. Production of nitric oxide in host-virus interaction: A case study with a compatible begomovirus-kenaf host-pathosystem. Plant Signall. Behav., 5: 668-676.
CrossRefDirect Link - Sheffield, F.M.L., 1933. The development of assimilatory tissue in Solanaceous hosts infected with aucuba mosaic of tomato. Ann. Applied Biol., 20: 57-69.
CrossRefDirect Link - Siddique, Z., K.P. Akhtar, A. Hameed, N. Sarwar, Imran-Ul-Haq and S.A. Khan, 2014. Biochemical alterations in leaves of resistant and susceptible cotton genotypes infected systemically by cotton leaf curl Burewala virus. J. Plant Interact., 9: 702-711.
CrossRefDirect Link - Singh, H.P., S. Kaur, D.R. Batish and R.K. Kohli, 2014. Ferulic acid impairs rhizogenesis and root growth and alters associated biochemical changes in Mung bean (Vigna radiata) hypocotyls. J. Plant Interact., 9: 267-274.
CrossRefDirect Link - Sinha, A. and M. Srivastava, 2010. Biochemical changes in mungbean plants infected by Mungbean yellow mosaic virus. Int. J. Virol., 6: 150-157.
CrossRefDirect Link - Song, X.S., Y.J. Wang, W.H. Mao, K. Shi, Y.H. Zhou, S. Nogues and J.Q. Yu, 2009. Effects of cucumber mosaic virus infection on electron transport and antioxidant system in chloroplasts and mitochondria of cucumber and tomato leaves. Physiologia Plantarum, 135: 246-257.
CrossRefDirect Link - Sulman, M., G. Fox, A. Osman, A. Inkerman, P. Williams and M. Michalowitz, 2001. Relationship between total peroxidase activity and susceptibility to black point in mature grain of some barley cultivars. Proceedings of the 10th Australian Barley Technical Symposium, September 16-20, 2001, Canberra, ACT, Australia.
- Sulmon, C., G. Gouesbet, I. Couee and A. El Amrani, 2004. Sugar-induced tolerance to atrazine in Arabidopsis seedlings: Interacting effects of atrazine and soluble sugars on psbA mRNA and D1 protein levels. Plant Sci., 167: 913-923.
CrossRefDirect Link - Tornero, P., R.A. Chao, W.N. Luthin, S.A. Goff and J.L. Dangl, 2002. Large-scale structure-function analysis of the Arabidopsis RPM1 disease resistance protein. Plant Cell, 14: 435-450.
CrossRefPubMedDirect Link - Upadhyaya, A., D. Sankhla, T.D. Davies, N. Sankhla and B.N. Smith, 1985. Effect of paclobutrazol on the activities of some enzymes of activated oxygen metabolism and lipid peroxidation in senescing soybean leaves. J. Plant Physiol., 121: 453-461.
CrossRefDirect Link - Shekhawat, U.K., T.R. Ganapathi and A.B. Hadapad, 2012. Transgenic banana plants expressing small interfering RNAs targeted against viral replication initiation gene display high-level resistance to banana bunchy top virus infection. J. Gen. Virol., 93: 1804-1813.
CrossRefDirect Link - Vanacker, H., T.L.W. Carver and C.H. Foyer, 1998. Pathogen-induced changes in the antioxidant status of the apoplast in barley leaves. Plant Physiol., 117: 1103-1114.
Direct Link - Vanitha, S.C., S.R. Niranjana and S. Umesha, 2009. Role of phenylalanine ammonia lyase and polyphenol oxidase in host resistance to bacterial wilt of tomato. J. Phytopathol., 157: 552-557.
CrossRefDirect Link - Watson, M.A., 1955. The effect of sucrose spraying on symptoms caused by beet yellows virus in sugar beet. Ann. Applied Biol., 43: 672-685.
CrossRefDirect Link - Yoshida, K., P. Kaothien, T. Matsui, A. Kawaoka and A. Shinmyo, 2003. Molecular biology and application of plant peroxidase genes. Applied Microbiol. Biotechnol., 60: 665-670.
CrossRefDirect Link - Zhao, C.J., A.R. Wang, Y.J. Shi, L.Q. Wang, W.D. Liu, Z.H. Wang and G.D. Lu, 2008. Identification of defense-related genes in rice responding to challenge by Rhizoctonia solani. Theoret. Applied Genet., 116: 501-516.
CrossRefDirect Link - Zhuang, B.C., B. Xu and L. Liao, 1993. Change of superoxide dismutase, peroxidase and storage protein in soyabean leaves after inoculation with soyabean mosaic virus. Acta Phytopathologica Sinica, 23: 261-265.
Direct Link - Kuroda, M., T. Oaiawa and H. Imagawa, 1990. Changes in chloroplast peroxidase activities in relation to chlorophyll loss in barley leaf segments. Physiologia Plantarum, 80: 555-560.
CrossRefDirect Link - Radwan, D.E.M., K.A. Fayez, S.Y. Mahmoud and G. Lu, 2010. Modifications of antioxidant activity and protein composition of bean leaf due to Bean yellow mosaic virus infection and salicylic acid treatments. Acta Physiologiae Plantarum, 32: 891-904.
CrossRefDirect Link - Jockusch, H., 1966. The role of host genes, temperature and polyphenoloxidase in the necrotization of TMV infected tobacco tissue. J. Phytopathol., 55: 185-192.
CrossRefDirect Link - Mohamed, H., A.A. El-Hady, M. Mansour and A.E.R. El-Samawaty, 2012. Association of oxidative stress components with resistance to flax powdery mildew. Trop. Plant Pathol., 37: 386-392.
CrossRefDirect Link - Rodriguez, M., E. Taleisnik, S. Lenardon and R. Lascano, 2010. Are Sunflower chlorotic mottle virus infection symptoms modulated by early increases in leaf sugar concentration? J. Plant Physiol., 167: 1137-1144.
CrossRefDirect Link